 2018, Vol. 42
2018, Vol. 42文章信息
- 王远超, 梁翠, 线薇微, 张效嘉. 2018.
- WANG Yuan-chao, LIANG Cui, XIAN Wei-wei, ZHANG Xiao-jia. 2018.
- 基于生态通道模型的长江口及邻近海域生态系统能流动态分析
- Ecopath based dynamic analyses of energy flows of Yangtze estuary and its adjacent waters
- 海洋科学, 42(5): 54-67
- Marine Sciences, 42(5): 54-67.
- http://dx.doi.org/10.11759/hykx20170816001
-
文章历史
- 收稿日期:2017-11-22
- 修回日期:2018-01-23




2. 中国科学院大学, 北京 100049;
3. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室, 山东 青岛 266071
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Marine Ecology and Environmental Science Laboratory, National Laboratory for Marine Science and Technology, Qingdao 266071, China
长江口是中国最大的河口, 三级分叉, 四级入海, 长江口及邻近海域水深一般不超过60 m, 属于中等强度潮汐的河口[1]。长江径流携带大量泥沙和有机物质, 为生物生存提供丰富的物质输送。由于人类海洋开发活动日益频繁, 长江口及邻近海域生态系统面临众多生态问题, 如水质污染、生境破碎化、生物多样性下降、群落结构简单化, 以及渔业资源严重衰退和渔场功能丧失等[2]。长江口生态系统目前处于亚健康状态, 并有向病态方向发展的趋势[3]。
生态系统能流网络模型被广泛应用于生态系统结构和功能研究, 这其中包括食物网、营养级、生态位重叠、生态效率、生态系统成熟度相关指标等, 可以评估渔业活动对生态系统结构与功能的影响, 对渔业资源管理具有重大意义。构建能流网络模型的方法有很多, 其中非常流行的就是构建生态通道模型(Ecopath模型), 该方法在海洋保护区研究、渔业资源评估、生态系统结构与功能分析等研究领域被广泛使用, 用于发现、建立渔业生产与生态系统营养级之间的相关性[4]。国内运用Ecopath进行生态系统能流网络建模的案例开始增多, 如胡菊香等[5]运用Ecopath模型对2010年汤浦水库水生态系统结构调控措施及实施效果进行评估, 并以Ecopath模型为辅助构建浮游生物完整性评价体系; 2016年杨超杰等[6]基于Ecopath模型估算莱州湾朱旺人工鱼礁区日本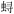
运用生态营养通道模型对长江口及邻近海域进行建模分析的研究, 主要以季节为时间尺度展开。林群等[7]研究了2000年秋季和2006年秋季长江口及毗邻水域生态系统能流结构, 结果显示2006年相对于2000年该区域碎屑食物链的重要性有所下降, 渔获物平均营养级下降, 系统均不成熟。韩瑞等[8]研究了2000年秋、2006年秋、2012年秋长江口水域在三峡工程蓄水前后的状态, 结果显示蓄水前中后期, 该水域均处于不成熟阶段, 蓄水后系统净效率和再循环率升高。而以一年为时间尺度进行模型构建的仅见张效嘉和线薇微[9]的研究, 他们构建了1985—1986年长江口及邻近海域生态系统生态通道模型, 结果显示1985—1986年该生态系统牧食食物链占据主导地位, 渔获物平均营养级为3.47, 在此研究基础上, 针对不同年份进行该区域能流结构与功能的对比分析还是缺乏的。
本文根据2004年长江口及邻近海域综合调查数据, 以一年为时间尺度, 基于Ecopath模型研究长江口及邻近海域生态系统能流网络, 了解长江口生态系统整体特征, 并以1985—1986年长江口及邻近海域综合调查[8]为背景, 探讨了该河口生态系统近20年来对自然干扰和人为干扰的响应, 现报道如下。
1 材料与方法 1.1 研究区域及数据来源本研究的调查区域为长江口及邻近海域, 经纬度为121°00′E~123°30′E、30°30′N~32°20′N, 在调查区域内共设置40个站位, 具体站位分布见图 1。
 |
| 图 1 长江口及邻近海域调查站位 Fig. 1 Sampling stations in Yangtze Estuary and its adjacent waters |
本研究于2004年2月、5月、8月和11月4个月份进行调查, 每个航次在122°E以东的35个站位中通常选取15个站位进行渔业资源拖网调查。渔业资源调查采用功率为255 kW的调查船, 调查网具为150.5 m×96.5 m轻拖网, 最大网衣网目为200 mm, 网目大小沿网身轴线向囊网递减, 囊网网目为30 mm, 调查站位拖速为2~3 kn, 拖网时间为1 h; 在调查期间对于所捕获的渔业资源进行分类鉴定, 并记录物种的质量和数量; 浮游植物样品通过小型浮游生物网垂直拖网获得, 浮游动物样品则是通过中型和大型浮游生物网垂直拖网获取; 底栖生物样品的获取是在各站位用采泥器(取样面积0.1 m2)取样, 经0.5 mm筛网冲洗获取。样品的获取与处理依据海洋调查规范GB/T12763-2007。
1.2 研究方法 1.2.1 Ecopath模型及基本原理Ecopath是一个静态模型, 以功能组为基本单位, 并遵循热力学定律, 研究生态系统能量流动网络。Ecopath包含两个核心方程, 其中方程(1)表明, 每个功能群必须保持质量守恒, 即捕捞量、消费量、生物量的积累和输出, 都不可以超过一个组的生产量, 因此需要对模型进行平衡处理, 确保生态营养效率(Ecological Efficiency, EE)不能超过1[10]。
各功能群生产量可以分为5个流向:
| $ {P_i} = {Y_i} + M{2_i} \times {B_i} + N{M_i} + B{A_i} + M{0_i} \times {B_i} $ | (1) |
式中,
Ecopath模型假定在给定一段时间内(通常为一年)系统能量守恒, 功能群间能量平衡通过方程(2)来确保:
| $ {Q_i} = {P_i} + {R_i} + {U_i} $ | (2) |
式中,
方程(1)也可表达为:
| $ \begin{array}{l} {B_i} \times {\left( {\frac{P}{B}} \right)_i} = \sum\limits_j {{B_j} \times } {\left( {\frac{Q}{B}} \right)_j} \times D{C_{ij}} + {Y_i} + N{M_i} + B{A_i} + \\ \quad \quad {B_i} \times {\left( {\frac{P}{B}} \right)_i} \times (1 - E{E_i}) \end{array} $ | (3) |
式中,
将生态学特征相同或相似的生物划为一组(如生长率、消费率、饵料、栖息水层及捕食物系等)[10-11]; 根据本文研究目的将相关的重要物种列为同一功能组[9, 12]。据此将2004年长江口及邻近海域生物划分为隐埋性底栖动物、头足类、蟹类、虾类、底层混合食性鱼类、底层游泳生物食性鱼类、底层无脊椎动物食性鱼类、中下层混合食性鱼类、中下层游泳生物食性鱼类、中下层浮游生物碎屑食性鱼类、中上层混合食性鱼类、中上层游泳生物食性鱼类、中上层浮游生物碎屑食性鱼类、水母、浮游动物、浮游植物、碎屑共17个功能组。
1.2.3 Ecopath模型数据输入与调试生物量(Biomass, B)主要通过2004年调查数据来估计, 其中鱼类、虾类、蟹类和头足类的生物量通过扫海面积法估计, 隐埋性底栖动物与浮游动物的生物量分别靠底泥样品和浮游动物垂直拖网样品估计, 有机碎屑生物量用有机碎屑与初级生产力的经验公式估算[9]。
在生态系统能量平衡情况下, 鱼类生产量/生物量(P/B)的值很难计算, 可以用总死亡系数Z估算[10], 总死亡系数的计算可以用(1)年龄结构捕捞曲线(age-structured catch curves)估计, 或者(2)运用体长转换捕捞曲线(length-converted catch curves)估计, 还可根据方程(3), 将总死亡系数Z分为自然死亡系数M、捕捞死亡系数F, 分别计算, 再进行加和。
渔获量Y通过中国渔业统计年鉴中的资料估计得出[9]。对于难以估计的参数(如Q/B和DC等), 则通过相关文献或历史资料[9], 并结合数据库FishBase来确定及经验公式来估算[10]。
Ecopath模型中, 生态系统的能量流动以生物湿重(t/km2)形式表示, 时间一般为1年。Ecopath模型参数输入后, 调整相关参数以使得所有的生态营养效率满足0 < EE < 1。Ecopath模型的准确性和实用性高低取决于参数来源的可靠性和准确性, 用Pedigree模块来评价数据和模型的整体质量[10]。
2 结果与分析 2.1 长江口及邻近海域功能组划分2004年长江口及邻近海域生态系统分为17个功能组, 各功能组所包含的生物种类见表 1。与1985—1986年调查结果相比[9], 此次水母作为一个新增功能组出现在本分析中。水母资源量在2004年左右显著提高。1998—2003年长江口及舟山渔场水母的相对资源密度(CPUE)在100 kg/h以上, 水母数量远高于90年代水平, 自20世纪90年代中后期起, 东海北部及黄海海域连年发生大型水母暴发现象, 其中2003年暴发程度最严重[15]。这种变化可能与近些年渔业捕捞强度的增加, 许多食水母鱼类资源衰退有关[16], 此变化直接影响河口能流结构。
| 序号 | 功能组 | 主要种类 |
| 1 | 隐埋性底栖动物 | 多毛类(Polychaeta)、软体类(Mollusca)、甲壳类(Crustacea)、棘皮类(Echinodermata)等 |
| 2 | 头足类 | 双喙耳乌贼(Sepiola birostrata), 四盘耳乌贼(Euprymna morsei), 日本枪乌贼(Loliolus japonica), 长蛸(Octopus variabilis) |
| 3 | 蟹类 | 双斑(Charybdis bimaculata), 口虾蛄(Oratosquilla oratoria), 三疣梭子蟹(Portunus trituberculatus), 红星梭子蟹(Portunus sanguinolentus), 日本(Charybdis japonica), 红线黎明蟹(Matuta planipes), 日本关公蟹(Dorippe japonica), 中华隆背蟹(Carcinoplax sinica) |
| 4 | 虾类 | 脊腹褐虾(Crangon affinis), 葛氏长臂虾(Palaemon gravieri), 脊尾白虾(Exopalaemon carinicauda), 戴氏赤虾(Metapenaeopsis dalei), 鲜明鼓虾(Alpheus distinguendus), 鹰爪虾(Trachypenaeus curvirostris), 细螯虾(Leptochela gracilis), 中国对虾(Fenneropenaeus chinensis) |
| 5 | 底层混合食性鱼类 | 虻鲉(Erisphex pottii), 棘头梅童鱼(Collichthys lucidus), 星点东方鲀(Takifugu niphobles), 细纹狮子鱼(Liparis tanakae), 暗纹东方鲀(Takifugu obscurus), 单指虎鲉(Minous monodactylus), 鲬(Platycephalus indicus), 蓝圆鲹(Decapterus maruadsi), 竹筴鱼(Trachurus japonicus), 短尾大眼鲷(Priacanthus macracanthus) |
| 6 | 底层游泳生物食性鱼类 | 黄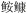 (Lophius litulon), 海鳗(Muraenesox cinereus), 长蛇鲻(Saurida elongata), 六指马鲅(Polynemus sextarius), 细鳞 (Lophius litulon), 海鳗(Muraenesox cinereus), 长蛇鲻(Saurida elongata), 六指马鲅(Polynemus sextarius), 细鳞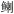 (Therapon jarbua) (Therapon jarbua) |
| 7 | 底层无脊椎动物食性鱼类 | 六丝矛尾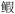 虎鱼(Chaeturichthys hexanema), 矛尾虎鱼(Chaeturichthys stigmatias), 细条天竺鲷(Apogonichthys lineatus), 短鳍红娘鱼(Lepidotrigla micropterus), 少鳞 虎鱼(Chaeturichthys hexanema), 矛尾虎鱼(Chaeturichthys stigmatias), 细条天竺鲷(Apogonichthys lineatus), 短鳍红娘鱼(Lepidotrigla micropterus), 少鳞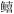 (Sillago japonica), 发光鲷(Acropoma japonicum), 绿鳍马面鲀(Thamnaconus modestus) (Sillago japonica), 发光鲷(Acropoma japonicum), 绿鳍马面鲀(Thamnaconus modestus) |
| 8 | 中下层混合食性鱼类 | 黄姑鱼(Nibea albiflora), 小黄鱼(Larimichthys polyactis), 皮氏叫姑鱼(Johnius belengerii), 白姑鱼(Argyrosomus argentatus) |
| 9 | 中下层游泳生物食性鱼类 | 带鱼(Trichiurus japonicus), 龙头鱼(Harpodon nehereus) |
| 10 | 中下层浮游生物碎屑食性鱼类 | 银鲳(Pampus argenteus), 燕尾鲳(Pampus nozawae), 刺鲳(Psenopsis anomala) |
| 11 | 中上层混合食性鱼类 | 刀鲚(Coilia ectenes), 鳓(Ilisha elongata), 鲐(Pneumatophorus japonicus) |
| 12 | 中上层游泳生物食性鱼类 | 蓝点马鲛(Scomberomorus niphonius), 油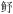 (Sphyraena pinguis) (Sphyraena pinguis) |
| 13 | 中上层浮游生物碎屑食性鱼类 | 赤鼻棱鳀(Thryssa kammalensis), 凤鲚(Coilia mystus), 黄鲫(Setipinna taty), 鳀(Engraulis japonicus), 七星底灯鱼(Benthosema pterotum), 康氏小公鱼(Anchoviella commersonii) |
| 14 | 水母 | 霞水母(Cyanea sp.) |
| 15 | 浮游动物 | 桡足类(Copepods), 枝角类(Cladocera), 糠虾类(Mysidacea), 端足类(Amphipoda), 磷虾类(Euphausiacea), 樱虾类(Sergestidae), 介形类(Ostracoda), 等足类(Isopoda), 毛颚类(Chaetognatha), 浮游贝类(Planktonic Mollusca), 被囊类(Tunicata)等 |
| 16 | 浮游植物 | — |
| 17 | 碎屑 | 溶解有机碳、颗粒有机碳 |
从生物量B(t/km2)的调查数据可以看出, 与1985—1986年调查结果[9]相比(图 2), 除了底层游泳生物食性鱼类、底层无脊椎动物食性鱼类、中下层混合食性鱼类、中上层游泳生物食性鱼类、水母、碎屑的生物量有所增加之外, 其余11个功能组的生物量都存在不同程度下降, 其中隐埋性底栖动物下降了4.57 t/km2, 浮游植物下降了4.09 t/km2, 相对变化较大; 碎屑的生物量增加量为124.85 t/km2, 增加幅度很大。水母的生物量为5.55 t/km2, 头足类2004年资源量与之前变化不大, 但是资源种类有所减少, 并有小型化的趋势。
 |
| 图 2 2004年与1985—1986年长江口及邻近海域生物量(不含碎屑)[9] Fig. 2 The Biomass of Yangtze Estuary and its adjacent waters ecosystem in 2004 and between 1985 and 1986(excluding detritus)[9] 功能组: 1.隐埋性底栖动物; 2.头足类; 3.蟹类; 4.虾类; 5.底层混合食性鱼类; 6.底层游泳生物食性鱼类; 7.底层无脊椎动物食性鱼类; 8.中下层混合食性鱼类; 9.中下层游泳生物食性鱼类; 10.中下层浮游生物碎屑食性鱼类; 11.中上层混合食性鱼类; 12.中上层游泳生物食性鱼类; 13.中上层浮游生物碎屑食性鱼类; 14.水母; 15.浮游动物; 16.浮游植物 Functional groups: 1. Benthos; 2. Cephalopods; 3. Crabs; 4. Shrimps; 5. Benthopelagic mixed-feeding fish; 6. Benthopelagic nekton- feeding fish; 7. Benthopelagic invertebrate-feeding fish; 8. Demersal mixed-feeding fish; 9. Demersal nekton-feeding fish; 10. Demersal plankton-feeding fish; 11. Pelagic mixed-feeding fish; 12. Pelagic nekton-feeding fish; 13. Pelagic plankton-feeding fish; 14. Jellyfish; 15. Zooplankton; 16. Phytoplankton |
EE是衡量生态系统能流的关键因子, 表示生产量中被捕食者消耗或者其他方式输出的生物量所占的比重, 刚刚超过0的数值表明这个功能组几乎不被其他的功能组消耗。
生态系统模型参数的输入和输出见表 2, 从中可以看出, 每个功能组的生态营养效率均为0 < EE < 1, 表明Ecopath模型通过调试, 生态系统的输入和输出已达到平衡, Ecopath模型的Pedigree指数为0.722, 表明质量可以接受[9]。表 2中水母和碎屑的EE分别为0.226和0.277, 相对较低; 隐埋性底栖动物的EE为0.047, 表明其几乎不被捕食者消耗, 输出也很少; 其他的功能组EE均超过0.900, 生态营养效率均值大于1985—1986年调查数据[9]。
| 功能群 | 营养级 | 生物量/ (t/km2) | 生产量/生物量 | 消费量/生物量 | 生态营养效率 | 渔获量/(t/(km–2·a)) |
| 头足类 | 4.34 | 0.172 | 3 | 9.25 | 0.981 | 0.506 |
| 中下层游泳生物食性鱼类 | 4.1 | 1.005 | 2.317 | 9.189 | 0.933 | 1.154 |
| 底层游泳生物食性鱼类 | 4.03 | 0.514 | 1.624 | 4.393 | 0.935 | 0.23 |
| 底层混合食性鱼类 | 3.95 | 0.338 | 2.197 | 18.011 | 0.989 | 0.726 |
| 中上层游泳生物食性鱼类 | 3.95 | 0.0679 | 1.713 | 10.863 | 0.923 | 0.107 |
| 中下层混合食性鱼类 | 3.44 | 1.518 | 2.236 | 7.779 | 0.99 | 0.288 |
| 虾类 | 3.02 | 0.367 | 8 | 28 | 0.972 | 0.285 |
| 中下层浮游生物碎屑食性鱼类 | 3.02 | 1.23 | 2.143 | 13.247 | 0.937 | 0.28 |
| 底层无脊椎动物食性鱼类 | 3.01 | 0.668 | 2.052 | 17.142 | 0.934 | — |
| 中上层混合食性鱼类 | 2.96 | 0.167 | 2.545 | 7.641 | 0.915 | 0.376 |
| 中上层浮游生物碎屑食性鱼类 | 2.95 | 5.211 | 2.74 | 12.885 | 0.933 | 0.234 |
| 蟹类 | 2.83 | 0.15 | 3.5 | 12 | 0.934 | 0.419 |
| 水母 | 2.1 | 5.552 | 5 | 20 | 0.226 | 0.036 |
| 隐埋性底栖动物 | 2.03 | 15.001 | 4.02 | 15.36 | 0.047 | 0.178 |
| 浮游动物 | 2.01 | 6.325 | 25 | 180 | 0.798 | 0.419 |
| 浮游植物 | 1 | 12.883 | 200 | — | 0.337 | — |
| 碎屑 | 1 | 217.692 | — | — | 0.277 | — |
| 注:斜粗体数字表示模型估计的参数, 表中数据按营养级从大到小排列。 | ||||||
1985—1986年调查结果[9]显示隐埋性底栖动物EE为0.751, 头足类EE为0.791, 碎屑EE为0.465, 浮游植物EE为0.503, 浮游动物EE没有变化, 为0.789。对比发现, 2004年隐埋性底栖生物的生物量为2.03 t/km2, 没有大的变化, 但是EE降低明显, 被捕食或输出量所占比例降低明显; 头足类的EE明显增加, EE接近1, 由于EE的值是很多因素综合作用的结果, 比如捕捞死亡、其他死亡因素等, 本次调查显示, 头足类捕捞死亡率为2.94, 排在所有功能组捕捞死亡率第一位, 这可能对解释这种现象有一定指导作用; 碎屑的EE下降明显, 流通受阻明显, 这也与碎屑生物量B的增加相一致, 2004年碎屑生物量为217.69 t/km2, 相对1985—1986年调查结果增加了134.47%。
2004年长江口及邻近海域生态系统EE总体平均值大于20年前, 但是浮游植物和碎屑的EE减小的幅度很大, 浮游植物趋向于沉积到碎屑中, 碎屑趋向于累积。对于河口生态系统碎屑食物链是很重要的环节, 其比重表征生态系统的成熟度[17], 2004年长江口及邻近海域碎屑食物链所占比重为44%, 与1985—1986年调查结果(43%)基本持平。说明碎屑量在增加, 但是碎屑中参与能量循环的碎屑所占比例在下降。
从图 3可以看出, 1985—1986年的调查结果[9]相比, 第三、四、五、六营养级的转换效率都有了很明显的提高, 在第四营养级和第六营养级处分别出现较高的转换效率, 而1985—1986年只在第四营养级处出现了转换效率的最高值。

2004年功能组的营养级为1~4.34, 与1985—1986年调查结果[9]相比, 最高营养级有所下降, 资源结构变动明显。其中, 第一营养级包括浮游植物与碎屑; 第二营养级包括水母、浮游动物、隐埋性底栖动物; 第三营养级包括蟹类、虾类、底层无脊椎动物食性鱼类、中下层浮游生物碎屑食性鱼类、中上层混合食性鱼类、中上层浮游生物碎屑食性鱼类、中下层混合食性鱼类; 第四营养级包括底层混合食性鱼类、头足类、底层游泳生物食性鱼类、中下层游泳生物食性鱼类、中上层游泳生物食性鱼类。
水母功能组营养级为2.10, 处在第二营养级附近, 与浮游动物、隐埋性底栖动物处在相近的营养级; 中下层混合食性鱼类营养级与1985—1986年的调查结果[9]一致; 底层无脊椎动物食性鱼类从1985—1986年[9]第四营养级(营养级为4.08)变化到第三营养级(营养级为3.01);在此次调查结果中, 头足类的营养级为4.34, 处在最高营养级, 而1985—1986年调查结果[9]显示当时头足类营养级为3.17, 头足类和底层无脊椎动物食性鱼类在食物链中的地位变动较大, 与其食物组成、渔业生产活动和环境变动有关系。表 3显示, 1985—1986年底层无脊椎动物食性鱼类中处于第四营养级的鱼类占比达到96.0%[9], 2004年本功能组99.9%的鱼类处于第三营养级, 这反映的是功能组食性上发生了变化, 趋向于低营养级食物。表 4显示, 1985—1986年[9]88.7%的头足类处在第三营养级, 到了2004年, 66.0%的头足类处在第四营养级。一方面可能本功能组第四营养级鱼类承受的捕食压力过大, 导致功能组总体生物量和物种比例发生变化, 另一方面, 环境的改变以及食物组成矩阵DC的准确性和时效性都可能引起这种结构变化, 营养级变动具体机制有待进一步研究。
图 4中纵坐标表示各个生态系统功能组所处的营养级, 能量用生物湿重(t/km2)表示, 灰色圆圈表示功能组, 圆圈的大小表示功能组生物量的大小, 图中线条表示能流比重, 红色越深表示能流比重越大。
 |
| 图 4 长江口及邻近海域能流图(单位: t/km2) Fig. 4 Energy flows of Yangtze estuary and its adjacent waters ecosystem (Unit: t/km2) |
图 4中展示了牧食食物链和碎屑食物链联合交织形成的网状结构, 可以看出牧食食物链中生产者浮游植物将能量主要传递给了第二营养级的浮游动物和水母:浮游植物在浮游动物和水母的能量来源中所占比例均超过60.22%, 其中浮游动物是最主要的初级消费者, 在长江口及邻近海域生态系统能量流动和物质循环中起到关键作用; 在碎屑食物链中, 碎屑在隐埋性底栖生物的能量来源中所占比达到了83.28%, 在水母和浮游动物的能量来源中所占比例均超过了30.11%。浮游动物与水母的生物量比较接近, 两者对于浮游植物和碎屑的消耗在整个能流网络中都占据很大的比例, 不同的是流入水母的能流相对于浮游动物而言, 并不能高效的传递到高营养级, 水母生物量的增加会加剧初级生产的消耗。头足类的主要能量来源是虾类、底层无脊椎动物食性鱼类和底层游泳生物食性鱼类; 蟹类的能量来源中, 隐埋性底栖生物、水母和碎屑占比较大, 主要是隐埋性底栖生物, 占比39.97%;虾类主要取食浮游动物; 中上层浮游生物碎屑食性鱼类在底层混合食性鱼类、中上层游泳生物食性鱼类能量来源中占比均超过93.08%, 为主要能量来源。
2.4 生态位重叠分析Ecopath模型采用Pianka的方法[18], 进行捕食者重叠(predator overlap)和被捕食者重叠(prey overlap)指数分析。如果指数为0, 说明两个功能群之间没有营养生态位的重叠; 如果为1, 则说明两者对相同的猎物有相同的需求, 存在竞争。
表 5显示, 一些功能组之间的重叠指数达到甚至接近0.90, 彼此之间存在较强竞争关系, 具体如下:虾类与底层无脊椎动物食性鱼类、虾类与中下层浮游生物碎屑食性鱼类、虾类与中上层混合食性鱼类、虾类与中上层浮游生物碎屑食性鱼类、底层混合食性鱼类与中上层游泳生物食性鱼类、底层无脊椎动物食性鱼类与中下层浮游生物碎屑食性鱼类、底层无脊椎动物食性鱼类与中上层混合食性鱼类、底层无脊椎动物食性鱼类与中上层浮游生物碎屑食性鱼类、中下层浮游生物碎屑食性鱼类与中上层混合食性鱼类、中下层浮游生物碎屑食性鱼类与中上层浮游生物碎屑食性鱼类、中上层混合食性鱼类与中上层浮游生物碎屑食性鱼类之间的捕食重叠指数均接近甚至超过0.90。
| 功能组 | 食物重叠指数 | ||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| 1 | 1 | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 2 | 0 | 1 | — | — | — | — | — | — | — | — | — | — | — | — | — |
| 3 | 0.391 | 0.036 | 1 | — | — | — | — | — | — | — | — | — | — | — | — |
| 4 | 0.028 | 0.002 | 0.347 | 1 | — | — | — | — | — | — | — | — | — | — | — |
| 5 | 0 | 0.065 | 0.002 | 0 | 1 | — | — | — | — | — | — | — | — | — | — |
| 6 | — | 0.1 | 0.003 | 0 | 0.003 | 1 | — | — | — | — | — | — | — | — | — |
| 7 | 0.03 | 0.002 | 0.019 | 0.854 | 0.001 | 0.001 | 1 | — | — | — | — | — | — | — | — |
| 8 | 0.023 | 0.033 | 0 | 0.688 | 0.621 | 0 | 0.722 | 1 | — | — | — | — | — | — | — |
| 9 | 0.01 | 0.461 | 0.023 | 0.323 | 0.231 | 0.054 | 0.29 | 0.433 | 1 | — | — | — | — | — | — |
| 10 | 0.03 | 0.002 | 0.121 | 0.945 | 0.001 | 0 | 0.962 | 0.73 | 0.311 | 1 | — | — | — | — | — |
| 11 | 0.081 | 0.002 | 0.059 | 0.878 | 0.001 | 0.001 | 0.995 | 0.735 | 0.304 | 0.968 | 1 | — | — | — | — |
| 12 | 0 | 0.057 | 0.001 | 0.001 | 0.999 | 0.002 | 0.001 | 0.626 | 0.23 | 0.001 | 0.001 | 1 | — | — | — |
| 13 | 0.09 | 0.002 | 0.024 | 0.856 | 0.001 | 0 | 0.997 | 0.728 | 0.295 | 0.964 | 0.997 | 0.001 | 1 | — | — |
| 14 | 0.575 | 0 | 0.187 | 0.138 | 0 | — | 0.134 | 0.108 | 0.05 | 0.136 | 0.158 | 0 | 0.16 | 1 | — |
| 15 | 0.57 | 0 | 0.177 | 0.016 | 0 | — | 0.007 | 0.006 | 0.003 | 0.007 | 0.029 | 0 | 0.033 | 0.986 | 1 |
| 注:功能组同图 2。 | |||||||||||||||
其中底层混合食性鱼类与中上层游泳生物食性鱼类、底层无脊椎动物食性鱼类与中上层混合食性鱼类、底层无脊椎动物食性鱼类与中上层浮游生物碎屑食性鱼类、中上层混合食性鱼类与中上层浮游生物碎屑食性鱼类之间的猎物重叠指数均大于0.99, 接近1, 说明这些组合之间在生态系统中存在非常明显的捕食竞争关系, 生物居住空间场所的空间生态位、捕食与被捕食关系的营养生态位、活动时间等要素非常接近, 鱼类的营养生境的大小与鱼类的活动能力及捕食水平有关。
2.5 混合营养影响(Mixed Trophic Impact, MTI)分析混合营养影响(Mixed Trophic Impact, MTI)[19]用于分析由于一个功能组的生物量改变对其他功能组生物量产生的效应, 即图 5中右侧图例所示功能组生物量稍微增加对于上侧功能组生物量产生的直接或者间接的影响。其中方向向上的空心方格表示正面影响, 方向向下的实心方格表示负面影响; 方格的高度代表影响程度的大小。
 |
| 图 5 2004年长江口及邻近海域混合营养影响 Fig. 5 MTI of Yangtze estuary and its adjacent waters ecosystem in 2004 |
图 5显示, 水母、底层混合食性鱼类、底层游泳生物食性鱼类、中下层游泳生物食性鱼类、中上层游泳生物食性鱼类等顶级捕食者对大部分的功能组会产生负面影响, 即处于较高营养级的捕食者会对它们所钟爱的猎物产生负面影响; 与此同时, 这一影响也会对它们猎物的猎物产生较弱的非直接的正面影响; 与之统一的是, 处于较低营养级的功能组会对其他大多数的功能组产生正面影响, 如浮游植物、浮游动物; 由于群落内部竞争, 几乎所有的功能组自身数量的稍微增加都会对自身产生负面影响; 同时可以看出, 捕捞(Fleet1)对大多数对象都有负面影响。
2004年捕捞对头足类和蟹类的影响相对于1985—1986年调查结果[9]有比较明显的变化, 1985—1986年捕捞对头足类的影响是正面的, 对于蟹类有相对较小的负面影响; 2004年的调查结果显示, 捕捞对头足类和蟹类有明显的负面影响, 这与头足类在我国的捕捞情况相一致。1983年之后, 头足类开始在长江口渔场出现短暂的渔汛, 一些渔业公司将头足类作为专门捕捞对象, 期间已有外来船只进行头足类的捕捞, 1983至1990年间渔获量表现稳定[20], 其代价是对于头足类的捕捞压力越来越大, 一方面加速了头足类种群的更新, 另一方面也加速了头足类的衰退。到了21世纪初, 作为渔业生产渔获的第二支撑, 头足类处于生态劣势地位[21], 以前的优势种严重衰退, 转变为小型种类[22]。蟹类和底层混和食性鱼类对渔业的影响由之前的负面影响变为正面影响, 中下层混合食性鱼类对中上层游泳生物食性鱼类的影响由正面影响变为负面影响; 虾类和蟹类对底层觅食性的鱼类在2004年显示为较弱或不明显的负面影响, 与1985—1986年调查结果[9]相比有所不同。
通过Ecopath模型评估显示, 浮游动物和中上层浮游生物碎屑食性鱼类是该生态系统的关键功能组, 关键功能组相对总影响指数(Relative total impact)分别为1和0.814。图 5显示, 浮游动物对水母、浮游动物、浮游植物和碎屑都有很强的负面影响, 而对于鱼类功能组皆为正面影响, 与1985—1986年相比, 2004年浮游动物生物量下降了12.4%, 作为初级生产者与较高营养级功能组之间联系的桥梁, 浮游动物生物量的降低对渔业资源生物量的上限会产生限制作用。中上层浮游生物碎屑食性鱼类在鱼类功能组中处于较低营养级, 属于较高营养级渔业资源的饵料生物。图 5显示, 该功能组对于浮游动物、碎屑、中上层混合食性鱼类和底层无脊椎动物食性鱼类等有较明显的负面影响, 而对于中上层游泳动物食性鱼类、底层混和食性鱼类等有着明显的正面影响, 与1985—1986年相比营养相互作用有着明显的变化, 1985—1986年研究结果显示中上层浮游生物碎屑食性鱼类对于各个鱼类功能组均呈现负面影响。
2.6 长江口及邻近海域生态系统能流分布图 6显示, 牧食食物链各个营养级占系统总流量Total system throughput (TST)/(t∙km–2∙a–1)的比值TST(%)从生产者的40.68%, 第二营养级的23.06%, 到第三营养级已经降到了2.029%, 第四营养级为0.327%, 第五营养级为0.0318%, 主要营养级可以确定为这五个。碎屑食物链中碎屑的TST(%)为33.87%。
 |
| 图 6 长江口及邻近海域线性食物链 Fig. 6 Linear food chain of Yangtze Estuary and its adjacent waters ecosystem |
表 6显示, 牧食食物链和碎屑食物链第二营养级的转换效率分别为9.9%和7.4%, 与1985—1986年的调查结果[9]相比, 第三、四、五、六营养级的转换效率都有了很明显的提高, 在第四营养级和第六营养级处分别出现较高的转换效率; 而1985—1986年只在第四营养级处出现了转换效率的最高值[9]。在总能流中直接来自碎屑的能量占比为44%, 直接来自初级生产者的能量占比为56%, 表明2004年长江口及邻近海域生态系统以牧食食物链为主要的能量传递途径, 辅之以碎屑食物链, 碎屑食物链所占能流比与1985—1986年接近。
| 来源 | 营养级 | |||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | |
| 2004年生产者 | — | 9.9 | 17.4 | 20.5 | 19.3 | 28.3 |
| 1985—1986年生产者[9] | 10.2 | 10.4 | 16.4 | 11.1 | 10.7 | |
| 2004年碎屑 | — | 7.4 | 17.8 | 20.4 | 19.4 | 28.3 |
| 1985—1986年碎屑[9] | 9.2 | 13.7 | 17.2 | 10.9 | 10.7 | |
| 2004总能流 | — | 8.8 | 17.6 | 20.5 | 19.3 | 28.3 |
| 1985—1986年总能流[9] | — | 9.7 | 11.7 | 16.8 | 11.0 | 10.7 |
| 2004年 | 1985—1986年[9] | |||||
| 碎屑食物链所占能流比: | 0.44 | 0.43 | ||||
| 转化效率(营养级Ⅱ-Ⅳ几何均数) | ||||||
| 初级生产者转换效率: | 15.2% | 12.0% | ||||
| 碎屑转换效率: | 13.9% | 12.9% | ||||
| 总转化效率: | 14.7% | 12.4% | ||||
长江口及邻近海域生态系统统计量详细刻画了生态系统的能流特征, 通过这些统计量我们可以对生态系统规模、生态系统成熟度[23-24]、渔业捕捞营养级[25]等方面做出分析, 见表 7。
| 参数 | 2004年Value | 1985—1986年Value[9] |
| 总消耗量Sum of all consumption (SC)/(t/(km2·a)) | 1619.914 | 1798.019 |
| 总输出量Sum of all exports (SEX)/(t/(km2·a)) | 1556.943 | 759.079 |
| 总呼吸量Sum of all respiratory flows (TR)/(t/(km2·a)) | 1019.639 | 1048.565 |
| 流向碎屑总量Sum of all flows into detritus (TDET)/(t/(km2·a)) | 2145.585 | 1415.223 |
| 系统总流量Total system throughput (TST)/(t/(km2·a)) | 6342.081 | 5020.886 |
| 系统总生产量Sum of all production (TP)/(t/(km2·a)) | 2852.892 | 2099.306 |
| 渔获物平均营养级Mean trophic level of the catch | 3.458 | 3.475 |
| 净效率Gross efficiency (catch/net p.p.) | 0.002 | 0.001 |
| 总净初级生产量Calculated total net primary production (NPP)/(t/(km2·a)) | 2576.600 | 1807.644 |
| 总初级生产量/总呼吸量Total primary production/total respiration (TPP/TR) | 2.527 | 1.724 |
| 系统净生产量Net system production (NSP)/(t/(km2·a)) | 1556.962 | 759.079 |
| 总初级生产量/总生物量Total primary production/total biomass (TPP/TB) | 50.355 | 31.483 |
| 总生物量/总流量Total biomass/total throughput (TB/T) | 0.008 | 0.011 |
| 总生物量Total biomass (excluding detritus, TB)/t/(km2·a)) | 51.169 | 57.416 |
| 总渔获量Total catch/(t/(km2·a)) | 5.238 | - |
| 系统连接指数Connectance Index (CI) | 0.539 | 0.471 |
| 系统杂食指数System Omnivory Index (SOI) | 0.069 | 0.103 |
| Finn循环指数Finn’s Cycling Index (FCI)% Of total throughput | 4.200 | 9.350 |
| Finn平均路径长度Finn’s mean path length (MPL) | 2.461 | 2.778 |
系统总流量(TST)是一个系统所有流量的总和, 从一个侧面反映系统的大小, 2004年TST为6342.081 t/(km2∙a), 相比于1985—1986年[9]的5020.886 t/(km2∙a)增加了26.31%; 2004年系统总生产量(TP)为2852.892 t/(km2∙a), 比1985—1986年[9]的2099.306 t/(km2∙a)增加了35.90%, 表明生态系统的规模增大明显。
2.7.2 生态系统成熟度(1) 总初级生产量/总呼吸量(TPP/TR):在生态系统发展期大于1或者小于1, 表示系统在增大或者缩小, 生物量在积累或者减少, 接近成熟期数值接近1, 在系统受到污染或者开发的时候会低于1[10], 2004年为2.527, 比1985—1986年[9]大, 说明长江口及邻近海域生态系统趋于不成熟, 资源趋于累积, 两个时期的比值都大于北部湾1997—1999年[26]的3.182, 大亚湾2012年[27]为2.18, 一定程度上说明长江口生态系统比北部湾生态系统成熟, 而大亚湾的成熟度以本指标来看要优于长江口。
(2) 总初级生产量/总生物量(TPP/TB):生态系统发展期值比较大, 说明资源量在积累, 成熟期较低。2004年为50.355, 比1985—1986年[9]大, 说明生态系统向资源积累的方向发展, 生态系统趋于不成熟。
(3) 总生物量/总流量(TB/T):直接表示生态系统成熟度的指标, 在生态系统成熟过程中会比较小, 并逐渐变大, 2004年为0.008, 比1985—1986年[9]小, 说明系统的成熟度降低了。
(4) 系统净生产量(NSP):系统总初级生产量与总呼吸量的差值, 成熟的生态系统数值接近0, 2004年为1556.962, 大于1985—1986年[9]的759.080, 系统成熟度降低。
(5) 食物链:生态系统发展期食物链以牧食食物链为主, 呈线状; 成熟期以碎屑食物链为主, 呈网状[17]。2004年生态系统流经碎屑食物链的能流占比44%, 1985—1986年[9]流经碎屑食物链的能流占比43%, 较为接近。Finn平均路径长度(MPL)表示流经食物链的每个循环的平均长度, 数值越高表示系统发展越成熟, 2004年为2.461, 1985—1986年[9]为2.778, 数值减小。每个循环的平均长度减小, 系统成熟度下降。FCI循环指数(FCI)表示生态系统中参与循环的能流在总能流中所占的比重, 数值越大说明再循环能流所占比例越大, 系统能流效率高, 系统成熟度高。2004年FCI为4.20, 比1985—1986年[9]减小明显, 说明成熟度下降, 系统再循环的效率下降。系统连接指数(CI)表示一个给定食物网实际链数与理论最大链数之比。系统杂食指数(SOI)表示不同营养级之间食性相互作用, 以食物摄入量的对数为权重。2004年与1985—1986年相比, CI增大, 食物网结构更加复杂; 而SOI变小, 如何解释两者的变化还不明了。
(6) 总生物量(TB):可以表征系统的成熟度, 一般认为系统在达到成熟的时候数值会达到最大, 2004年为51.169, 比1985—1986年[9]小, 说明系统成熟度降低了。
以上指标说明, 2004年长江口及邻近海域生态系统相较于1985—1986年[9]变得不成熟, 生态系统向着相反的方向发展, 属于退行演替(retrogressive succession)。
2.7.3 渔获物平均营养级渔获物平均营养级(Mean trophic level of the catch)和净效率(Gross efficiency)分别从静态和动态两个方面表示渔业现状, 渔获物平均营养级是表示渔业资源总体利用水平的指标, 2004年为3.458, 接近最高营养级, 长江口及邻近海域渔业捕捞主要集中在较高营养级类群。随着渔业的发展, 对渔业资源的捕捞压力增加, 渔业捕捞将向低营养级变化, 20世纪90年代北部湾渔获物平均营养级为2.920[28], 低于长江口, 说明渔业生产对于北部湾的影响较大, 渔业资源相对脆弱。净效率在不同的生态系统之间变化较大, 值越大表示渔业捕捞主要集中在低营养级, 随着渔业的发展, 这个值会增大。2004年长江口及邻近海域为0.002, 1985—1986年为0.0009[9], 说明渔获物平均营养级有向低营养级演化的趋势, 渔业资源有趋于退化的迹象, 渔业生产发展, 对生态系统的压力增大。
3 结论根据2004年调查结果, 将长江口及邻近海域生态系统划分为隐埋性底栖动物、头足类、蟹类、虾类、底层混合食性鱼类、底层游泳生物食性鱼类、底层无脊椎动物食性鱼类、中下层混合食性鱼类、中下层游泳生物食性鱼类、中下层浮游生物碎屑食性鱼类、中上层混合食性鱼类、中上层游泳生物食性鱼类、中上层浮游生物碎屑食性鱼类、水母、浮游动物、浮游植物, 1个碎屑功能组, 共17个功能组。各个功能组营养级为1~4.34, 头足类和底层无脊椎动物食性鱼类在食物链中的营养级发生了较大幅度的变动; 从生物量可以看出, 20年间, 在所调查的17个功能组中, 有11个功能组的生物量都出现了不同程度的下降; 生态位重叠分析得出的结果表明一些功能组之间在营养生态位、空间生态位上有着明显的相似性, 对于维持生态系统结构的稳定有着至关重要的作用; 混合营养分析的结果表明, 2004年捕捞对头足类和蟹类的混合营养效应, 与20年前[9]相比出现了明显的反转现象; 2004年长江口及邻近海域生态系统各个营养级之间能量转换效率明显提高, 这和功能组之间生物量的变动有关, 同时受自然因素和人为因素的影响; 流经食物网每个循环的长度减小, 参与循环的能流在总能流中所占的比重减小, 生态系统能流网络趋向简单化。
与1985—1986年调查数据[9]对比, 2004年长江口及邻近海域生态系统总体趋向于不成熟, 能流再循环的效率降低, 生态系统向着相反的方向发展, 属于退行演替(Regressive succession), 外界干扰对生态系统的压力较大, 引起生态系统结构和功能变化的环境要素及机理, 包括渔业捕捞、环境污染、三峡大坝蓄水带来的河口水文、化学组成及含沙量的变化[29]等, 都需要进一步深入地研究; 渔业捕捞平均营养级呈下降趋势, 渔业发展, 对生态系统的压力增大, 渔业资源有衰退的趋势。长江口及邻近海域生态系统不健康程度相对增加, 对于长江口及邻近海域生态系统的管理与保护急需引起人们的关注。本文用Ecopath模型分析了近20年间长江口及邻近海域生态系统结构的变动情况, 可以为生物多样性保护、海域生态环境治理与改善、渔业资源评估与管理、渔业资源生态学等研究提供参考。
| [1] |
罗秉征. 河口及近海的生态特点与渔业资源[J]. 长江流域资源与环境, 1992, 1(1): 24-30. Luo Bingzheng. Ecological characteristics and fishery resources of the Yangtze river estuary and adjacent sea[J]. Resources and environment in the Yangtze Basin, 1992, 1(1): 24-30. |
| [2] |
王金辉, 徐韧, 秦玉涛, 等. 长江口基础生物资源现状及年际变化趋势分析[J]. 中国海洋大学学报(自然科学版), 2006, 36(5): 821-828. Wang Jinhui, Xu Ren, Qin Yutao, et al. The basic biological resources and variation during the last decades in the Changjiang (Yangtze river) Estuary[J]. Periodical of Ocean University of China, 2006, 36(5): 821-828. |
| [3] |
周晓蔚, 王丽萍, 郑丙辉. 长江口及毗邻海域生态系统健康评价研究[J]. 水利学报, 2011, 42(10): 1201-1208, 1217. Zhou Xiaowei, Wang Liping, Zheng Binghui. Ecosystem health assessment for the Changjiang River Estuary and its adjacent sea area[J]. Journal of Hydraulic Engineering, 2011, 42(10): 1201-1208, 1217. |
| [4] |
仝龄, 唐启升, DanielPauly. 渤海生态通道模型初探[J]. 应用生态学报, 2000, 11(3): 435-440. Tong Ling, Tang Qisheng, Daniel Pauly. A preliminary approach on mass-balance Ecopath model of the Bohai Sea[J]. Chinese Journal of Applied Ecology, 2000, 11(3): 435-440. DOI:10.3321/j.issn:1001-9332.2000.03.027 |
| [5] |
胡菊香, 米玮洁, 沈强, 等.汤浦水库水生态系统结构调控措施及实施效果评价[C].成都: 2016年中国水产学会学术年会论文摘要集, 2016, 129. Hu Juxiang, Mi Weijie, Shen Qiang, et al. Application and evaluation of the regulation and control measures of the ecosystem in Tangpu Reservoir[C]. Chengdu: China Society of Fisheries Academic conference Abstract Set In 2016, 2016, 129. |
| [6] |
杨超杰, 吴忠鑫, 刘鸿雁, 等. 基于Ecopath模型估算莱州湾朱旺人工鱼礁区日本 、脉红螺捕捞策略和刺参增殖生态容量[J]. 中国海洋大学学报, 2016, 46(11): 168-177.Yang Chaojie, Wu Zhongxin, Liu Hongyan, et al. The fishing strategy of Charybdis japonica and Rapana venosa and the carrying capacity of Apostichopus japonucus in Zhuwang, Laizhou artificial reef ecosystem based on Ecopath model[J]. Periodical of Ocean University of China, 2016, 46(11): 168-177. |
| [7] |
林群, 金显仕, 郭学武, 等. 基于Ecopath模型的长江口及毗邻水域生态系统结果和能量流动研究[J]. 水生态学杂志, 2009, 2(2): 28-36. Lin Qun, Jin Xianshi, Guo Xuewu, et al. Study on the structure and energy flow of the Yangtze river estuary and adjacent waters ecosystem based on Ecopath model[J]. Journal of Hydroecology, 2009, 2(2): 28-36. |
| [8] |
韩瑞, 陈求稳, 王丽, 等. 基于生态通道模型的长江口水域生态系统结构与能量流动分析[J]. 生态学报, 2016, 36(15): 4907-4918. Han Rui, Chen Qiuwen, Wang Li, et al. Analysis of the ecosystem structure and energy flow of the Yangtze River estuary and adjacent seas based on the Ecopath model[J]. Acta Ecologica Sinica, 2016, 36(15): 4907-4918. |
| [9] |
张效嘉, 线薇微. 1985-1986长江口生态系统能流网络分析[J]. 海洋科学, 2016, 40(7): 60-72. Zhang Xiaojia, Xian Weiwei. Energy network analysis and fish community in adjacent waters of Yangtze estuary[J]. Marine Sciences, 2016, 40(7): 60-72. |
| [10] |
Villy Christensen, Carl J. Walters, Daniel Pauly and Robyn Forrest. Ecopath with Ecosim version 6 User Guide[EB/OL]. Lenfest Ocean Futures Project, http://www.ecopath.org/2008-11-21.
|
| [11] |
许思思, 宋金明, 李学刚, 等. 渔业捕捞对渤海渔业资源及生态系统影响的模型研究[J]. 资源科学, 2011, 33(6): 1153-1162. Xu Sisi, Song Jinming, Li Xuegang, et al. Modeling of Fishing Effects on Fishery Resources and Ecosystems of the Bohai Sea[J]. Resources Science, 2011, 33(6): 1153-1162. |
| [12] |
罗秉征, 韦晟, 窦硕增. 长江口鱼类食物网与营养结构的研究[J]. 海洋科学集刊, 1997, 38(1): 143-153. Luo Bingzheng, Wei Sheng, Dou Shuozeng. Study on food web and trophic structure of fish in the changjiang river estuary[J]. Studia Marina Sinica, 1997, 38(1): 143-153. |
| [13] |
Polis G A. Food webs, trophic cascades, and community structure[J]. Aust J Ecol, 1994, 19: 121-136. |
| [14] |
Shannon L J, Moloney C L, Jarre A, et al. Trophic flows in the southern Benguela during the 1980s and 1990s[J]. Journal of Marine Systems, 2003, 39: 83-116. DOI:10.1016/S0924-7963(02)00250-6 |
| [15] |
高倩, 陈佳杰, 徐兆礼, 等. 长江口及邻近海域浮游水螅水母、管水母和栉水母的丰度分布与季节变化[J]. 生态学报, 2015, 35(22): 7328-7337. Gao Qian, Chen Jiajie, Xu Zhaoli, et al. Abundance distribution and seasonal variation of medusa, siphonophores, and ctenophores in the Changjiang (Yangtze River) estuary and the adjacent East China Sea[J]. Acta Ecologica Sinica, 2015, 35(22): 7328-7337. |
| [16] |
严利平, 李圣法, 丁峰元. 东海、黄海大型水母类资源动态及其与渔业关系的初探[J]. 海洋渔业, 2004, 26(01): 9-12. Yan Liping, Li Shengfa, Ding fengyuan. The preliminary studies on the dynamics of Macro-jellyfish resources and their relationship with fisheries in the East China Sea and Yellow Sea[J]. Marine Fisheries, 2004, 26(01): 9-12. DOI:10.3969/j.issn.1004-2490.2004.01.002 |
| [17] |
沈国英, 黄凌风, 郭丰, 等. 海洋生态学[M]. 北京: 科学出版社, 2010. Shen Guoying, Huang Lingfeng, Guo Feng, et al. Marine Ecology[M]. Beijing: CSPM, 2010. |
| [18] |
Pianka E P. Niche overlap and diffuse competition[J]. Proc Natl Acad Sci USA, 1974, 71(5): 2141-2145. DOI:10.1073/pnas.71.5.2141 |
| [19] |
Ulanowicz R, Puccia C. Mixed trophic impacts in ecosystems[J]. Coenoses, 1990, 5(1): 7-16. |
| [20] |
张明云. 1991-1993年东黄海鱿等头足类拖网生产概况[J]. 海洋渔业, 1994, 16(5): 208-210. Zhang Mingyun. Cephalopods (squid, et al.) trawl production situation in the east Yellow Sea during 1991- 1993[J]. Marine Fisheries, 1994, 16(5): 208-210. |
| [21] |
李建生, 李圣法, 任一平. 长江口渔场渔业生物群落结构的季节变化[J]. 中国水产科学, 2004, 11(5): 432-439. Li Jiansheng, Li Shengfa, Ren Yiping. Seasonal variety of fishery biology community structure in fishing ground of the Yangtze estuary[J]. Journal of Fishery Sciences of China, 2004, 11(5): 432-439. DOI:10.3321/j.issn:1005-8737.2004.05.009 |
| [22] |
覃涛, 俞存根, 陈全震, 等. 舟山渔场及邻近海域头足类(Cephalopod)种类组成和数量分布[J]. 海洋与湖沼, 2011, 42(1): 124-130. Qin Tao, Yu Cungen, Chen Quanzhen, et al. Species composition and quantitative distribution study on Cephalopod in the Zhoushan fishing ground and adjacent waters[J]. Oceanologia et Limnologia Sinica, 2011, 42(1): 124-130. |
| [23] |
Odum E P. Fundamentals of Ecology[M]. Philadelphia: W B Saunders Co, 1971: 574.
|
| [24] |
Odum E P. The strategy of ecosystem development[J]. Science, 1969, 104: 262-270. |
| [25] |
Marta C, Alberto S, Isabel P, et al. An ecological model of the Northern and Central Adriatic Sea: Analysis of ecosystem structure and fishing impacts[J]. Journal of Marine Systems, 2007, 67: 119-154. DOI:10.1016/j.jmarsys.2006.10.002 |
| [26] |
陈作志, 邱永松, 贾晓平, 等. 捕捞对北部湾海洋生态系统的影响[J]. 应用生态学报, 2008, 19(7): 1604-1610. Chen Zuozhi, Qiu Yongsong, Jia Xiaoping, et al. Effects of fishing on the marine ecosystem of Beibu Gulf[J]. Chinese Journal of Applied Ecology, 2008, 19(7): 1604-1610. |
| [27] |
莫宝霖, 秦传新, 陈丕茂, 等. 基于Ecopath模型的大亚湾海域生态系统结构与功能初步分析[J]. 南方水产科学, 2017, 13(2): 9-19. Mo Baolin, Qin Chuanxin, Chen Pimao, et al. Preliminary analysis of structure and function of Daya Bay ecosystem based on Ecopath model[J]. South China Fisheries Science, 2017, 13(2): 9-19. DOI:10.3969/j.issn.2095-0780.2017.02.002 |
| [28] |
陈作志, 邱永松, 贾晓平, 等. 基于Ecopath模型的北部湾生态系统结构和功能[J]. 中国水产科学, 2008, 15(3): 460-468. Chen Zuozhi, Qiu Yongsong, Jia Xiaoping, et al. Structure and function of Beibu Gulf ecosystem based on Ecopath model[J]. Journal of Fishery Sciences of China, 2008, 15(3): 460-468. DOI:10.3321/j.issn:1005-8737.2008.03.012 |
| [29] |
线薇微, 刘瑞玉, 罗秉征. 三峡水库蓄水前长江口生态与环境[J]. 长江流域资源与环境, 2004, 13(2): 119-122. Xian Weiwei, Liu Ruiyu, Luo Bingzheng. Environment of the Changjiang Estuary before the Sluice Construction in the three gorges reservoir[J]. Resources and Environment in the Yangtze Basin, 2004, 13(2): 119-122. DOI:10.3969/j.issn.1004-8227.2004.02.004 |