 2022, Vol. 46
2022, Vol. 46文章信息
- 徐东会, 齐衍萍, 张炫钊, 杜小媛, 韩龙江, 刘潇, 周辉, 潘玉龙. 2022.
- XU Dong-hui, QI Yan-ping, ZHANG Xuan-zhao, DU Xiao-yuan, HAN Long-jang, LIU Xiao, ZHOU Hui, PAN Yu-long. 2022.
- 秦皇岛近岸海域夏季小型水母分布特征
- Distribution characteristics of small medusae in the Qinhuangdao coastal waters during summer
- 海洋科学, 46(11): 29-37
- Marine Sciences, 46(11): 29-37.
- http://dx.doi.org/10.11759/hykx20211013001
-
文章历史
- 收稿日期:2021-10-13
- 修回日期:2022-04-24



2. 自然资源部 渤海生态预警与保护修复重点实验室, 山东 青岛 266033
2. Key Laboratory of Ecological Prewarning and Protection of Bohai Sea, Ministry of Natural Resources, Qingdao 266033, China
全球近海生态系统受到气温升高、人类活动加剧、渔业资源过度捕捞等压力的影响, 海洋生态灾害频发。我国渤、黄、东海自20世纪90年代起开始频发水母灾害, 给渔业资源、沿海工业和旅游业等带来严重影响: 2003年黄东海水母暴发导致渔业减产60%; 2013年起, 海月水母旺发连年威胁红沿河核电站, 甚至造成非计划停机; 营口、秦皇岛、青岛等多地海水浴场均发生过水母蛰人致死事件, 水母灾害日趋严重, 这种现象不仅在海洋中容易观察到的大型水母(伞径 > 2.5 cm)种类中发生, 也发生在不容易观察到的小型水母(伞径 < 2.5 cm)种类中[1-7]。水母暴发受到前所未有的重视, 成为全球性问题。目前, 针对水母生态学价值的研究大多集中在包括钵水母在内的大型水母上, 而对包括水螅水母和栉水母在内的小型水母的研究较少, 多集中在生物形态特征、分类描述和水平分布等方面[5-9]。
秦皇岛地区海洋经济快速发展, 城市人口日渐增多, 对秦皇岛地区的海洋生态环境带来巨大压力。该海域海水浴场、海水养殖业众多且发达, 但近年来频频发生的水母暴发对其海洋经济发展和生态环境带来巨大压力。目前, 已有不少关于秦皇岛水域水母特征的分析研究, 但主要集中于大型水母的群落特征方面, 迄今少见关于该海域小型水母的专题报道[10-12]。本研究利用2014—2018年每年8月在秦皇岛海域开展的海洋调查所获得的浮游动物资料, 分析了小型水母群落的种类组成和数量变化, 对该海域小型水母的分布特征进行研究, 探讨了小型水母群落与环境因子之间的关系, 以期为小型水母暴发研究积累参考资料, 并为秦皇岛海域的生态环境保护提供科学依据。
1 材料与方法 1.1 调查海域及方法分别于2014年8月6日—18日、2015年8月7日—23日、2016年8月10日—29日、2017年8月9日—26日和2018年8月2日—27日, 使用渔船在秦皇岛海域(滦河口-北戴河生态监控区)开展共计24个站位的浮游动物调查(见图 1)。采用标准的浮游生物大网(网口直径80 cm, 网衣长270 cm, 筛绢孔径505 μm)由底层到表层垂直拖网的方式采集小型水母样品, 用5%福尔马林海水溶液对样品进行固定后带回实验室完成分类鉴定。
 |
| 图 1 调查区域及站位分布 Fig. 1 Area of the investigation and sampling stations |
同步监测了所有调查站位的表层温度、盐度、pH、溶解氧、悬浮物、无机氮、磷酸盐、叶绿素a。参照《海洋监测规范—海水分析》(GB17378—2007)完成水质样品的采集和分析测定。采用pH计法测定pH、碘量法测定溶解氧、重量法测定悬浮物、次溴酸盐氧化法测定铵盐、萘乙二胺分光光度法测定亚硝酸盐、锌-镉还原法测定硝酸盐、抗坏血酸还原的磷钼蓝法测定磷酸盐、荧光分光光度法《海洋监测规范》(GB17378.4—2007)测定叶绿素。
1.2 数据处理与分析 1.2.1 丰度根据每立方米水体中的个体数计算小型水母丰度。
1.2.2 优势种小型水母的优势种是根据每个水母种类的优势度值(Y)来确定, 以Y≥0.02的种类作为优势种[13]。
| $ Y=(n_{i}/N)×f_{i}, $ | (1) |
式中, ni为第i种水母的个体数量, N为所有种类水母的总个体数, fi为第i种水母在各站位出现的频率。
1.2.3 小型水母与环境因子关系小型水母丰度与环境因子之间的关系[14], 采用多元统计软件PRIMER V6.1的主成分分析(PCA)、BIOENV和RELATE程序进行分析。环境要素主要包括: 温度、盐度、pH、溶解氧、悬浮物、无机氮、磷酸盐、叶绿素a及水深等。通过SPSS v18.0软件的Kruskal-Wallis H程序对2014—2018年夏季秦皇岛水域环境要素的差异性进行检验, 采用多元回归程序对小型水母丰度与浮游动物丰度的相关性进行分析。
2 结果 2.1 环境因子2014—2018年夏季秦皇岛水域表层温度、盐度、pH、溶解氧、悬浮物、无机氮、磷酸盐、叶绿素a的年际变化如图 2显示。秦皇岛水域表层温度变化范围为25.9~27.8 ℃, 平均温度为26.9 ℃ (图 2a); 秦皇岛水域表层盐度变化范围为29.9~ 30.8, 平均盐度为30.5(图 2b); 秦皇岛水域表层pH变化范围为8.0~8.2, 平均pH为8.0(图 2c); 秦皇岛水域表层溶解氧浓度变化范围为7.3~9.2 mg/L, 平均溶解氧浓度为8.2 mg/L(图 2d); 秦皇岛水域表层悬浮物浓度变化范围为9.8~24.6 mg/L, 平均悬浮物浓度为18.5 mg/L(图 2e); 秦皇岛水域表层无机氮浓度变化范围为0.061~0.171 mg/L, 平均无机氮浓度为0.122 mg/L(图 2f); 秦皇岛水域表层磷酸盐浓度变化范围为0.002~0.008 mg/L, 平均磷酸盐浓度为0.006 mg/L(图 2g); 秦皇岛水域表层叶绿素浓度变化范围为3.5~13.5 μg/L, 平均叶绿素浓度为6.8 μg/L (图 2h)。差异性检验分析结果显示: 2014—2018年夏季秦皇岛水域, 不同年份间温度、盐度、pH、溶解氧、悬浮物、无机氮、磷酸盐和叶绿素a等环境因子差异极显著(P < 0.01)。
 |
| 图 2 2014—2018年夏季秦皇岛水域表层环境因子变化 Fig. 2 Variations in the environmental factors at the surface layer in the Qinhuangdao coastal waters during the summer 2014–2018. |
2014—2018年夏季秦皇岛水域环境因子的PCA分析结果表明, 5个年份采样站位的海水环境存在明显差异(图 3)。在PCA1方向上, 影响不同年份水母群落的环境因子依次为pH、溶解氧、悬浮物、叶绿素、盐度、无机氮、磷酸盐、温度和水深等。在PCA2方向上, 影响不同年份水母群落的环境因子依次为无机氮、盐度、磷酸盐、温度、水深和叶绿素等。
 |
| 图 3 环境因子PCA排序 Fig. 3 PCA sequence diagram of the environmental factors |
2014—2018年夏季秦皇岛水域浮游动物调查共鉴定水母2门, 包括水螅水母12种、栉水母1种, 合计种类数为13种(表 1)。2018年夏季浮游动物总种类数为30种, 水母种类数仅为2种, 在2014—2018年间种类数最少, 占浮游动物总种类数的6.7%; 2014年夏季浮游动物总种类数为33种, 水母种类数为7种, 占浮游动物总种类数的21.2%; 2015年夏季浮游动物总种类数为36种, 水母种类数为7种, 占浮游动物总种类数的19.4%; 2016年夏季浮游动物总种类数为42种, 水母种类数为6种, 占浮游动物总种类数的14.3%; 2017年夏季浮游动物总种类数为36种, 水母种类数为7种, 占浮游动物总种类数的19.4%。
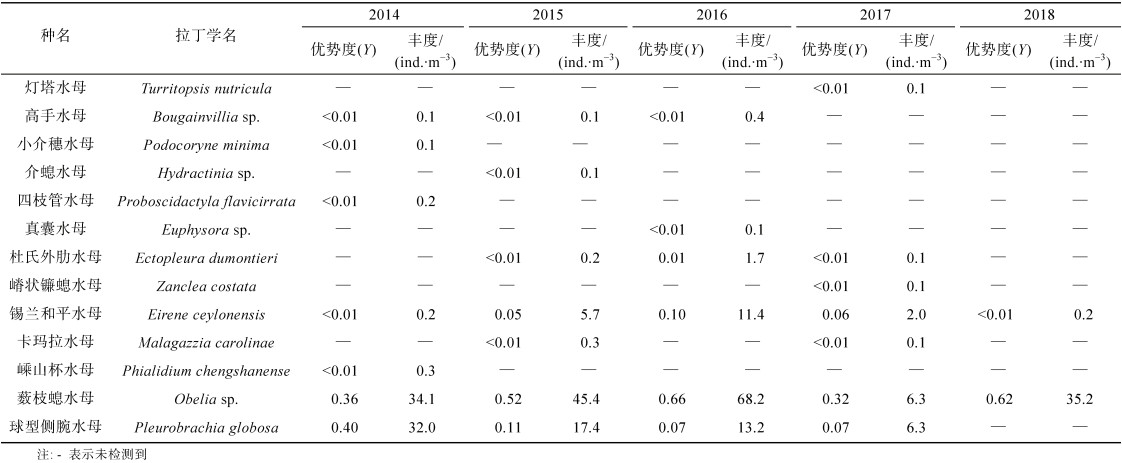
|
夏季秦皇岛水域水母优势种类包括水螅水母和栉水母(表 1), 其中2014年水母的优势种包括薮枝螅水母(Obelia sp.)和球型侧腕水母(Pleurobrachia globosa); 2015和2016年水母的优势种均包括锡兰和平水母(Eirene ceylonensis)、薮枝螅水母和球型侧腕水母; 2017年水母的优势种包括锡兰和平水母、薮枝螅水母和球型侧腕水母; 2018年水母的优势种为薮枝螅水母。
2.3 水母丰度及水平分布特征2014—2018年夏季秦皇岛水域浮游动物的总平均丰度为330.7 ind./m3, 水母的平均丰度为56.2 ind./m3, 占浮游动物总平均丰度的17.4%, 其中水螅水母类平均丰度为42.4 ind./m3, 栉水母类平均丰度为13.8 ind./m3 (表 1)。图 4结果显示, 2014—2018年5年间夏季秦皇岛水域水母丰度在2014—2016年呈现逐年增长的趋势, 2017年水母丰度下降明显, 2018年水母丰度略有增加。2016年夏季秦皇岛水域水母的平均丰度最高, 为95.0 ind./m3占浮游动物总丰度(675.1 ind./m3)的14.1%; 2017年夏季秦皇岛水域水母的平均丰度最低, 为14.9 ind./m3占浮游动物总丰度(162.4 ind./m3)的9.2%; 2014年夏季秦皇岛水域水母丰度为66.9 ind./m3, 占浮游动物总丰度(241.7 ind./m3)的27.68%; 2015年夏季秦皇岛水域水母丰度为69.1 ind./m3, 占浮游动物总丰度(293.0 ind./m3)的23.6%; 2018年夏季秦皇岛水域水母丰度为35.4 ind./m3, 占浮游动物总丰度(281.4 ind./m3)的12.6%。2014—2018年水螅水母类的数量占绝对优势, 而栉水母类的数量表现出明显的逐年下降趋势(图 4)。多元回归分析结果显示: 2014—2016年, 小型水母丰度与浮游动物丰度极显著正相关(P < 0.01); 2017年, 小型水母丰度与浮游动物丰度正相关; 2018年, 小型水母丰度与浮游动物丰度显著正相关(P < 0.05)。
 |
| 图 4 2014—2018年夏季秦皇岛水域水母丰度变化 Fig. 4 Variations in medusae abundance in the Qinhuangdao coastal waters during summer 2014–2018 |
2014—2018年夏季秦皇岛水域水母总丰度的平面分布见图 5。2014年夏季秦皇岛水域水母总丰度较高, 其平均值为66.9 ind./m3, 总丰度最高值出现在调查水域西南部4号站(359.4 ind./m3)。2015年夏季秦皇岛水域水母总丰度较高, 其平均值为69.1 ind./m3, 总丰度最高值出现在调查水域西北部23号站(620.0 ind./m3)。2016年夏季秦皇岛水域水母总丰度很高, 其平均值为95.0 ind./m3, 总丰度最高值出现在调查水域西南部1号站(770.0 ind./m3)。2017年夏季秦皇岛水域水母总丰度很低, 其平均值为14.9 ind./m3, 总丰度最高值出现在调查水域西南部1号站(118.7 ind./m3)。2018年夏季秦皇岛水域水母总丰度较低, 其平均值为35.4 ind./m3, 总丰度最高值出现在调查水域西部12号站(237.5 ind./m3)。2016年夏季秦皇岛水域水母总丰度 > 2015年夏季秦皇岛水域水母总丰度 > 2014年夏季秦皇岛水域水母总丰度 > 2018年夏季秦皇岛水域水母总丰度 > 2017年夏季秦皇岛水域水母总丰度。
 |
| 图 5 2014—2018年夏季秦皇岛水域水母丰度(ind./m3)平面分布 Fig. 5 Horizontal distribution of medusae abundance (ind./m3) in the Qinhuangdao coastal waters during summer 2014–2018 |
通过BIOENV分析的水母与环境因子间的相关性系数结果见表 2。双因子分析结果显示, 2014年水母丰度与pH/磷酸盐的相关性最高(P < 0.01); 2015年水母丰度与溶解氧/无机氮的相关性最高(P < 0.01); 2016年水母丰度与磷酸盐/温度的相关性最高(P < 0.01); 2017年水母丰度与无机氮/温度的相关性最高(P < 0.01); 2018年水母丰度与悬浮物/温度的相关性最高(P < 0.01)。三因子分析结果显示, 2014年水母丰度与pH/溶解氧/磷酸盐的相关性最高(P < 0.01); 2015年水母丰度与溶解氧/无机氮/温度的相关性最高(P < 0.01); 2016年水母丰度与盐度/磷酸盐/温度的相关性最高(P < 0.01); 2017年水母丰度与pH/无机氮/温度的相关性最高(P < 0.01); 2018年水母丰度与悬浮物/无机氮/温度的相关性最高(P < 0.01)。四因子分析结果显示, 2014年水母丰度与pH/盐度/溶解氧/磷酸盐的相关性最高(P < 0.01); 2015年水母丰度与溶解氧/悬浮物/无机氮/温度的相关性最高(P < 0.01); 2016年水母丰度与盐度/悬浮物/磷酸盐/温度的相关性最高(P < 0.01); 2017年水母丰度与pH/盐度/无机氮/温度的相关性最高(P < 0.01); 2018年水母丰度与pH/悬浮物/无机氮/温度的相关性最高(P < 0.01)。
| 因子 | 参数 | Spearman相似性系数ρs | ||||
| 2014 | 2015 | 2016 | 2017 | 2018 | ||
| 双因子 | pH/磷酸盐 | 0.257** | ||||
| 溶解氧/无机氮 | 0.471** | |||||
| 磷酸盐/温度 | 0.321** | |||||
| 无机氮/温度 | 0.267** | |||||
| 悬浮物/温度 | 0.440** | |||||
| 三因子 | pH/溶解氧/磷酸盐 | 0.284** | ||||
| 溶解氧/无机氮/温度 | 0.452** | |||||
| 盐度/磷酸盐/温度 | 0.365** | |||||
| pH/无机氮/温度 | 0.263** | |||||
| 悬浮物/无机氮/温度 | 0.383** | |||||
| 四因子 | pH/盐度/溶解氧/磷酸盐 | 0.290** | ||||
| 溶解氧/悬浮物/无机氮/温度 | 0.436** | |||||
| 盐度/悬浮物/磷酸盐/温度 | 0.387** | |||||
| pH/盐度/无机氮/温度 | 0.235** | |||||
| pH/悬浮物/无机氮/温度 | 0.318** | |||||
| 注: *表示P<0.05; **表示P<0.01 | ||||||
秦皇岛位于渤海中部西侧, 2014—2018年夏季秦皇岛水域表层温度和盐度的年际变化呈现先升高后下降的趋势; 表层pH、溶解氧和叶绿素呈现总体上升的趋势; 表层悬浮物、无机氮呈现总体下降的趋势; 表层磷酸盐呈现先下降后上升的趋势。通过不同年份间环境因子的差异性检验, 2014—2018年夏季秦皇岛水域, 温度、盐度、pH、溶解氧、悬浮物、无机氮、磷酸盐和叶绿素a等环境因子差异极其显著。
秦皇岛水域水母的群落组成特征存在明显的年际差异, 2014—2018年夏季秦皇岛水域共记录小型水母13种分属2门, 其中包括12种水螅水母和1种栉水母, 历年夏季水母种类数介于6~7种, 但是在2018年夏季水母种类数仅为2种; 2014—2017年夏季秦皇岛水域水母的优势种类均包含薮枝螅水母和球型侧腕水母, 而2018年水母的优势种仅有薮枝螅水母; 历年夏季水母种类数占浮游动物总种类数的6.7%~21.2%。张万磊等[11]在2009—2011年进行的秦皇岛海域调查中, 对浮游动物的分析和鉴定结果总共记录了26种小型水母。本研究与其相比, 小型水母的种类数呈现显著的下降趋势。
2014—2016年夏季, 秦皇岛海域水母丰度呈逐年上升趋势, 2017年水母丰度显著下降, 2018年略有上升。秦皇岛海域水母丰度在2016年夏季最高, 2017年夏季最低。2014年至2018年, 水螅水母数量占绝对优势, 而栉水母数量呈现明显的逐年下降趋势。多元回归分析结果显示, 2014—2018年, 小型水母丰度与浮游动物丰度正相关。2014—2018年夏季秦皇岛水域水母总丰度的平面分布结果显示, 出现水母丰度最高值的调查站位均位于调查区域的近岸海域。在野外, 水母水螅体多附着在硬质或半硬质的基质上, 例如岩礁、贝壳、牡蛎壳、大型藻类表面、塑料玻璃等人工材料, 实验表明, 水母水螅体对附着的基质材料有一定的偏好性, 海湾扇贝壳最适合水螅体增殖, 其次海洋中人工材料的增多为水螅体繁殖提供了良好的附着基质[15]。多年来秦皇岛海港区和山海关区邻近海域养殖用海、码头、港口、人工岛等的建设为水母生长提供了良好的附着基, 这很可能是导致调查区域近岸海域水母丰度较高的重要因素之一。
PCA和RELATE分析结果表明, 2014—2018年夏季秦皇岛海域水母丰度与不同环境因子存在显著相关性。由于不同种类的水母对环境因子的适应性不同, 不同年份水母群落对环境因子的依赖表现也不同。高倩等[16]分析了2002年至2003年在长江口及其附近海域获得的海洋调查数据。结果表明, 水温是影响调查海区小型水母总丰度季节变化的主要环境因素, 小型水母丰度与表层温度呈显著正相关。同时, 盐度是影响小型水母平面分布格局的主要环境因子, 与夏季中层盐度呈正相关。赵亮等[17]研究发现, 夏、秋季水层中18~25 ℃的持续时间越长, 越有利于水螅体足囊萌发和第二年水螅横裂的补充, 导致第二年沙海蜇数量有增加的趋势, 而秦皇岛海域8月多年来温度为25~28 ℃, 略高于海月水母和沙海蜇等大型水母的最适生长温度。研究表明, 营养盐浓度升高及氮磷比例的改变会导致浮游生物数量的增加, 从而为水母种群的快速扩增提供物质基础, 而多个种类的水母及其水螅体均可对低氧环境产生较强的耐受性, 这一特性可使其能够适应多种生境[5]。郗艳娟等[18]对河北省沿海水域的海水理化因子与水母丰度进行了灰色关联分析。结果表明, 活性磷酸盐、无机氮、溶解氧、温度和盐度是影响河北沿海水母丰度的主要因素。本研究结果与其相似, 秦皇岛夏季水域水母群落结构的年际变化主要受温度、溶解氧、pH值、磷酸盐、无机氮、悬浮物和盐度等环境因素的影响。
4 结论1) 2014—2018年夏季, 秦皇岛近岸水域水母的群落组成特征存在明显的年际差异。在2014—2017年间秦皇岛水域水母种类数差异不大且种类组成丰富, 而2018年夏季水母种类数最少, 仅有2种。2014—2018年, 秦皇岛近岸水域水母的优势种包括薮枝螅水母、球型侧腕水母和锡兰和平水母。2014—2016年夏季, 秦皇岛水域水母丰度呈现逐年增长的趋势, 而2017年水母丰度显著下降, 2018年水母丰度略有增加。2016年夏季秦皇岛水域水母丰度最高, 2017年夏季秦皇岛水域水母丰度最低。2)秦皇岛夏季水域水母群落结构的年际变化, 主要受到温度、溶解氧、pH、磷酸盐、无机氮、悬浮物、盐度等环境因子和浮游动物等生物因子的显著影响。2014年水母丰度与pH、盐度、溶解氧和磷酸盐等环境因子的相关性最高; 2015年水母丰度与溶解氧和无机氮等环境因子的相关性最高; 2016年水母丰度与盐度、悬浮物、磷酸盐和温度等环境因子的相关性最高; 2017年水母丰度与无机氮和温度等环境因子的相关性最高; 2018年水母丰度与悬浮物和温度等环境因子的相关性最高; 2014—2018年, 水母丰度与生物因子浮游动物丰度均表现为正相关。
| [1] |
PITT K A, PURCELL J E. Jellyfish blooms: cause, conse-quences, and recent advances[J]. Hydrobiologia, 2009, 616: 1-289. DOI:10.1007/s10750-008-9599-2 |
| [2] |
PURCELL J E, GRAHAM W M, DUMONT H J. Jellyfish blooms: ecological and societal importance[J]. Hydrobiologia, 2001, 451: 1-333. DOI:10.1023/A:1011883019506 |
| [3] |
SUN X, WANG S, SUN S. Introduction to the China jellyfish project[J]. Chinese Journal of Oceanology and Limnology, 2011, 29(2): 491-492. DOI:10.1007/s00343-011-0491-y |
| [4] |
郭东晖, 李刚, 何静. 2007-2011年春季珠江口中东部水域水母研究[J]. 海洋与湖沼, 2012, 43(3): 584-588. GUO Donghui, LI Gang, HE Jing. Ecological studies on the medusae in the mid-eastern Pearl River estuary during the spring of 2007-2011[J]. Oceanologia et Lim nologia Sinica, 2012, 43(3): 584-588. |
| [5] |
张芳, 孙松, 李超伦. 海洋水母类生态学研究进展[J]. 自然科学进展, 2009, 19(2): 121-130. ZHANG Fang, SUN Song, LI Chaolun. Advances in Marine jellyfish ecology[J]. Progress In Natural Scien ce, 2009, 19(2): 121-130. DOI:10.3321/j.issn:1002-008X.2009.02.001 |
| [6] |
孙松. 对黄、东海水母暴发机理的新认知[J]. 海洋与湖沼, 2012, 43(3): 406-410. SUN Song. New perception of jellyfish bloom in the East China Sea and Yellow Sea[J]. Oceanologia et Limnologia Sinica, 2012, 43(3): 406-410. |
| [7] |
孙松, 张芳, 李超伦, 等. 黄海小型水母的分布特征[J]. 海洋与湖沼, 2012, 43(3): 429-437. SUN Song, ZHANG Fang, LI Chaolun, et al. The distribution pattern of small medusae in the Yellow Sea[J]. Oceanologia et Limnologia Sinica, 2012, 43(3): 429-437. |
| [8] |
黄旭光, 郭东晖, 肖武鹏, 等. 九龙江口春季微型浮游生物数量变动及其与小型水母消长的关系[J]. 海洋与湖沼, 2012, 43(3): 579-583. HUANG Xuguang, GUO Donghui, XIAO Wupeng, et al. The relationship between quantitative changes of microplankton and population dynamics of small medusa in the Jiulong River Estuary in spring of 2011[J]. Oceanologia et Limnologia Sinica, 2012, 43(3): 579-583. |
| [9] |
冯颂, 孙松, 李超伦, 等. 胶州湾半球美螅水母(Clytia hemisphaerica)数量周年变动及对浮游动物摄食压力估算[J]. 海洋与湖沼, 2012, 43(3): 464-470. FENG Song, SUN Song, LI Chaolun, et al. Annual changes in Clytia hemisphaerica abundance and its grazing pressure on zooplankton populatin in the Jiaozhou Bay[J]. Oceanologia et Limnologia Sinica, 2012, 43(3): 464-470. |
| [10] |
郑向荣, 李燕, 饶庆贺, 等. 秦皇岛近海大型水母暴发性增长原因探析[J]. 河北渔业, 2014, 2: 16-20. HENG Xiangrong, LI Yan, RAO Qinghe, et al. An analysis of the cause of explosive of macro-jellyfish in inshore Qinghuangdao area[J]. Hebei Fisheries, 2014, 2: 16-20. |
| [11] |
张万磊, 李莉, 张建乐. 秦皇岛海域水母类特征分析[J]. 河北渔业, 2015, 2: 16-18. ZHANG Wanlei, LI Li, ZHANG Jianle. Features of jellyfishes in Qinhuangdao Sea area[J]. Hebei Fisheries, 2015, 2: 16-18. |
| [12] |
刘婧美, 饶庆贺. 秦皇岛近岸海域水母暴发原因分析及防治对策研究[J]. 河北渔业, 2016, 1: 55-57. LIU Jingmei, RAO Qinghe. Study of the cause & countermeasures of jellyfishes outbreaks at offshore area in Qinhuangdao[J]. Hebei Fisheries, 2016, 1: 55-57. |
| [13] |
MCHAUGHTON S J. Relationships among functional properties of Californian grassland[J]. Nature, 1967, 216(5111): 168-168. |
| [14] |
SOUISSI S, IBANEZ F, HAMADOU R B, et al. A new multivariate mapping method for studying species assemblages and their habitats: example using bottom trawl surveys in the Bay of Biscay (France)[J]. Sarsia, 2001, 86: 527-542. |
| [15] |
王彦涛, 孙松, 李超伦. 海月水母水螅体附着选择性研究[J]. 海洋与湖沼, 2012, 43(6): 1091-1095. WANG Yantao, SUN Song, LI Chaolun. Study on substrate choice and settlement preferences of planula larvae of Aurelia sp. 1[J]. Oceanologia et Limnologia Sinica, 2012, 43(6): 1091-1095. |
| [16] |
高倩, 陈佳杰, 徐兆礼, 等. 长江口及邻近海域浮游水螅水母、管水母和栉水母的丰度分布与季节变化[J]. 生态学报, 2015, 35(22): 7328-7337. GAO Qian, CHEN Jiajie, XU Zhaoli, et al. Abundance distribution and seasonal variation of medusae, siphonophores, and ctenophores in the Changjiang (Yangtze River) Estuary and the adjacent East China Sea[J]. Acta Ecologica Sinica, 2015, 35(22): 7328-7337. |
| [17] |
赵亮, 李夏, 张芳. 黄海水温与沙海蜇丰度年际变化的相关分析[J]. 海洋与湖沼, 2016, 47(3): 564-571. ZHAO Liang, LI Xia, ZHANG Fang. On relationship between inter-annual variation in water temperature regime and Nemopilema nomurai abundance in the Yellow Sea[J]. Oceanologia et Limnologia Sinica, 2016, 47(3): 564-571. |
| [18] |
郗艳娟, 吴新民, 郑向荣, 等. 灰关联分析法研究水母密度与海水理化因子的关系[J]. 河北渔业, 2012, 8: 5-6. XI Yanjuan, WU Xinmin, ZHENG Xiangrong, et al. The grey relational analysis of the relationship between jellyfish density and physical and chemical properties of seawater[J]. Hebei Fisheries, 2012, 8: 5-6. |