 2022, Vol. 46
2022, Vol. 46文章信息
- 张亮, 王俊健, 王岚, 张海舟, 张乃星, 蒲思潮. 2022.
- ZHANG Liang, WANG Jun-jian, WANG Lan, ZHANG Hai-zhou, ZHANG Nai-xing, PU Si-chao. 2022.
- 青岛近岸海域浮游动物群落结构特征
- Zooplankton community in the sea near Qingdao
- 海洋科学, 46(8): 88-100
- Marine Sciences, 46(8): 88-100.
- http://dx.doi.org/10.11759/hykx20210920001
-
文章历史
- 收稿日期:2021-09-20
- 修回日期:2022-05-21




2. 国家海洋局北海预报中心, 山东 青岛 266061;
3. 北部湾大学海洋学院, 广西 钦州 535011
2. North China Sea Marine Forecasting Center of SOA, Qingdao 266061, China;
3. College of Oceanography, Beibu Gulf University, Qinzhou 535011, China
青岛市位于山东半岛南岸, 其近海及邻近海域位于南黄海西北部。青岛近岸分布有多个海湾, 如鳌山湾、胶州湾、灵山湾、琅琊台湾等, 是我国重要的渔业养殖和港口海域。随着海洋经济的迅速发展, 青岛近海海域海洋环境面临日益加大的压力[1], 近海海洋水体环境容易受到人为扰动, 引起海洋生物群落变化, 进而影响海洋生态系统的结构与功能[2]。浮游动物是海洋生态系统中的次级生产力, 是海洋生物多样性的重要组成[3], 浮游动物种类组成、群落动态变化对于整个海洋生态系统的结构和功能都有着十分重要的影响[4]。
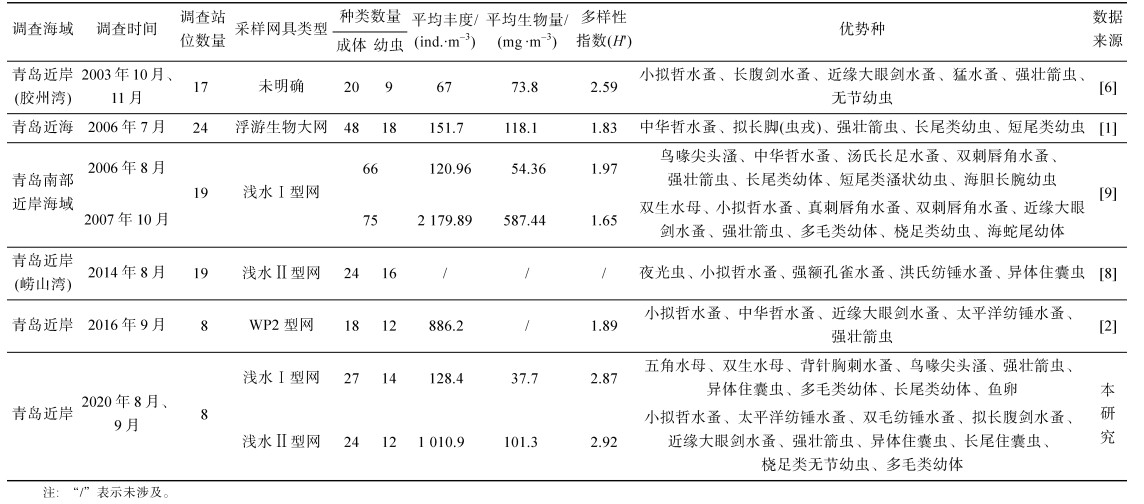
目前, 对青岛邻近海域浮游动物群落特征的研究多集中于胶州湾[5-7]、崂山湾[8], 针对整个青岛近岸海域浮游动物的研究也有报道, 王晓等[1]利用2006年7月在青岛近海海域的调查资料, 研究了青岛近海浮游动物的种类组成、群落结构及多样性; 王宁等[2]利用2015—2016年在青岛近海海域采集的浮游动物样品, 分析了浮游动物的种类组成、丰度、优势种、生物多样性以及季节变化; 寇建山[9]根据2006年8月、12月, 2007年4月、10月在青岛南部近岸海域的浮游动物调查资料, 研究了该海域浮游动物的种类组成、丰度、生物量和优势种, 分析了该海域浮游动物的群落结构, 并探讨了浮游动物群落结构特征与环境因子的相关性。
浮游动物种类组成及数量变化能够反应环境变化状况, 可作为环境监测的生物性指标, 在海洋环境保护中有重要意义。为合理地开发利用海洋资源, 科学地保护海洋环境, 及时掌握近岸海域浮游动物最新的群落结构特征显得尤为重要。为了解青岛近岸海域浮游动物的群落结构特征, 本文以2020年8—9月调查数据为基础, 分析青岛海域浮游动物群落结构, 调查中同时使用浅水Ⅰ、Ⅱ型浮游生物网采集浮游动物, 浅水Ⅱ型浮游生物网网目较小, 避免了仅使用浅水Ⅰ浮游生物网采集导致大量小型浮游动物的遗漏, 同时使用浅水Ⅰ、Ⅱ型浮游生物网采集浮游动物能够更全面和准确地研究浮游动物群落结构, 同时对比分析了浅水Ⅰ、Ⅱ型浮游生物网所采集浮游动物的差异。并将本次调查研究结果与以往调查资料进行对比, 为深入研究该海域浮游动物群落结构提供重要的基础数据。
1 材料与方法 1.1 采样与分析于2020年8月20—22日、29日, 9月22日、28日在青岛近岸海域调查采集浮游动物, 共布设8个采样站位(图 1), 站位水深为6.0~35.0 m。浮游动物采用浅水Ⅰ型浮游生物网(网口面积为0.20 m2, 材质为CQ14筛绢)和浅水Ⅱ型浮游生物网(网口面积为0.08 m2, 材质为CB36筛绢)采集, 样品自底(距离海底2 m)至表垂直拖网获得。采集样品用5%福尔马林海水溶液固定, 带回实验室后按照《海洋监测规范》[10]浮游生物生态调查中的相关要求, 采用体视显微镜以个体计数法进行分析, 分析包括种类鉴定、计数, 并利用电子天平进行浮游动物湿重生物量的测定。本研究还同时对与浮游动物群落有重要影响的环境因子进行监测, 主要包括水深、水温、盐度和叶绿素a等。
 |
| 图 1 采样站位 Fig. 1 Location of sampling stations |
丰度、生物量和优势度按照以下公式进行计算:
| $ C=\frac{n}{V}, $ | (1) |
| $ M=\frac{m}{V}, $ | (2) |
| $ Y=\frac{n_i}{N} \times f_i, $ | (3) |
式中, C为某种类的丰度(ind./m3), n为该站位浮游动物某种类的个数(ind.), V为该站位滤水量(m3), M为某种类的生物量(mg/m3), m为该站位浮游动物某种类的质量(mg), Y为某种类的优势度, ni为第i种浮游动物的个体数(ind.), N为浮游动物的总个体数(ind.), fi为第i种浮游动物在调查站位中出现的频率。当优势度Y≥0.02时, 认定该物种为优势种[11]。
1.2.2 生物多样性采用香农-韦弗(Shannon-Weaver)多样性指数(H′)[12]、Margalef种类丰富度指数(D)[13]和Pielou均匀度指数(J)[14]进行浮游动物群落生物多样性分析, 公式如下:
| $ H^{\prime}=-\sum\limits_{i=1}^s P_i \log _2 P_i, $ | (4) |
| $ D=\frac{S-1}{\log _2 N}, $ | (5) |
| $ J=\frac{H^{\prime}}{\log _2 S}, $ | (6) |
式中,
采用ArcMap 10.4.1软件对获得的浮游动物丰度、生物量及多样性的平面分布情况进行图件绘制。
1.2.4 生物多样性评价陈清潮等[15]根据多样性指数的大小将生物多样性阈值(Dv=H′×J)分为5个等级, Dv < 0.6, 为Ⅰ级(多样性差), 0.6≤Dv≤1.5为Ⅱ级(多样性一般), 1.6≤Dv≤2.5为Ⅲ级(多样性较好), 2.6≤Dv≤3.5为Ⅳ级(多样性丰富), Dv > 3.5为Ⅴ级(多样性非常丰富)。根据调查的结果, 依据陈清潮等[15]对群落丰富程度的分级对青岛近岸近海域浮游动物的多样性进行评价分析。
1.2.5 群落划分采用PRIMER 6.0中的CLUSTER模块对浮游动物群落进行聚类分析, 并进行MDS标序分析。在进行群落结构分析时, 为减少机会种对群落结构的干扰, 先去掉总体中相对丰度小于1%的种, 但保留其中在任一站位相对丰度大于3%的种。原始的丰度数据经四次方根转化和标准化后, 以Bray-Curtis相似性系数为基础构建相似性矩阵, 然后使用等级聚类分析将样品逐级连接成组, 通过树枝图来表示群落结构[16]。非度量MDS标序按照样品间的非相似性等级顺序将样品排放在标序图中, 通过压力系数(stress)的范围判断分析结果的可用性: stress < 0.01, 结果完全可信; 0.01 < stress < 0.05, 可信; 0.05 < stress < 0.1, 基本可信; 0.1 < stress < 0.2, 结果有参考价值, 但某些细节不可信[17]。
2 结果与分析 2.1 水文环境2020年8—9月调查海域环境因子见表 1。调查海域表层温度为22.40~26.96 ℃, 平均为24.29 ℃; 底层温度为21.64~26.83 ℃, 平均为23.25 ℃; 透明度为1.5~12.0 m, 平均为4.50 m; 表层溶解氧质量浓度为6.48~8.05 mg/L, 平均为7.09 mg/L; 底层溶解氧质量浓度为5.21~7.14 mg/L, 平均为6.42 mg/L; 表层盐度为29.807~31.411, 平均为30.822; 底层盐度为30.800~31.565, 平均为31.240; 表层叶绿素a质量浓度为0.332~9.71 μg/L, 平均为3.45 μg/L; 底层叶绿素a质量浓度为0.561~3.84 μg/L, 平均为1.98 μg/L。透明度和盐度分布趋势一致, 总体上呈近岸低, 远岸高; 叶绿素a质量浓度总体上呈近岸高, 远岸低的趋势; 调查海域南部水温低于北部; 溶解氧无明显分布规律。
| 站位 | 水深/m | 层次 | 水温/℃ | 透明度/m | 溶解氧质量浓度/(mg·L–1) | 盐度 | 叶绿素a质量浓度/(μg·L–1) |
| A1 | 25.0 | 表 | 24.97 | 3.0 | 8.05 | 30.893 | 7.23 |
| 底 | 22.51 | 6.03 | 31.252 | 3.84 | |||
| A2 | 25.0 | 表 | 26.96 | 12.0 | 7.20 | 30.671 | 1.05 |
| 底 | 26.83 | 5.21 | 31.462 | 1.05 | |||
| A3 | 6.0 | 表 | 25.86 | 1.5 | 6.69 | 30.306 | 2.01 |
| A4 | 8.5 | 表 | 23.60 | 2.5 | 6.91 | 30.924 | 3.28 |
| A5 | 6.0 | 表 | 24.26 | 2.0 | 6.48 | 29.807 | 9.71 |
| A6 | 32.0 | 表 | 22.40 | 10.0 | 7.16 | 31.411 | 0.332 |
| 底 | 21.64 | 6.91 | 31.565 | 0.561 | |||
| A7 | 15.0 | 表 | 22.95 | 3.0 | 7.26 | 30.957 | 2.85 |
| 底 | 22.27 | 7.14 | 31.105 | 3.33 | |||
| A8 | 35.0 | 表 | 23.29 | 2.0 | 6.94 | 30.809 | 1.16 |
| 底 | 23.02 | 6.79 | 30.800 | 1.11 |
本次调查共发现浮游动物成体29种(见表 2), 其中水螅水母类7种, 占总种数的24%; 管水母类和被囊类各2种, 分别占总种数的7%; 桡足类14种, 占总种数的48%; 栉水母类、枝角类、端足类和毛颚类各1种, 分别占总种数的3%; 另外, 发现浮游幼虫14类。
| 序号 | 中文名 | 拉丁文名或英文名 | 类群 | 浅水Ⅰ型网 | 浅水Ⅱ型网 |
| 1 | 半球美螅水母 | Clytia hemisphaerica | 水螅水母类 | + | + |
| 2 | 薮枝螅水母 | Obelia spp. | 水螅水母类 | + | + |
| 3 | 嵊山秀氏水母 | Sugiura chengshanense | 水螅水母类 | + | + |
| 4 | 鳞茎高手水母 | Bougainvillia muscus | 水螅水母类 | + | |
| 5 | 嵴状镰螅水母 | Zanclea costata | 水螅水母类 | + | |
| 6 | 细颈和平水母 | Eirene menoni | 水螅水母类 | + | |
| 7 | 黑球真唇水母 | Eucheilota menoni | 水螅水母类 | + | + |
| 8 | 五角水母 | Muggiaea atlantica | 管水母类 | + | + |
| 9 | 双生水母 | Diphyes chamissonis | 管水母类 | + | + |
| 10 | 球型侧腕水母 | Pleurobrachia globosa | 栉水母类 | + | + |
| 11 | 中华哲水蚤 | Calanus sinicus | 桡足类 | + | + |
| 12 | 小拟哲水蚤 | Paracalanus parvus | 桡足类 | + | + |
| 13 | 腹针胸刺水蚤 | Centropages abdominalis | 桡足类 | + | |
| 14 | 瘦尾胸刺水蚤 | Centropages tunuiremis | 桡足类 | + | + |
| 15 | 背针胸刺水蚤 | Centropages dorsispinatus | 桡足类 | + | + |
| 16 | 双刺唇角水蚤 | Labidocera bipinnata | 桡足类 | + | + |
| 17 | 真刺唇角水蚤 | Labidocera euchaeta | 桡足类 | + | + |
| 18 | 太平洋真宽水蚤 | Eurytemora pacifica | 桡足类 | + | |
| 19 | 太平洋纺锤水蚤 | Acartia pacifica | 桡足类 | + | + |
| 20 | 双毛纺锤水蚤 | Acartia bifilosa | 桡足类 | + | + |
| 21 | 拟长腹剑水蚤 | Oithona similis | 桡足类 | + | + |
| 22 | 近缘大眼剑水蚤 | Corycaeus affinis | 桡足类 | + | + |
| 23 | 猛水蚤 | Harpacticoida | 桡足类 | + | + |
| 24 | 捷氏歪水蚤 | Tortanus derjugini | 桡足类 | + | |
| 25 | 鸟喙尖头溞 | Penilia avirostris | 枝角类 | + | + |
| 26 | 钩虾 | Gammaridea | 端足类 | + | |
| 27 | 强壮箭虫 | Sagitta crassa | 毛颚类 | + | + |
| 28 | 异体住囊虫 | Oikopleura dioica | 被囊类 | + | + |
| 29 | 长尾住囊虫 | Oikopleura lonicauda | 被囊类 | + | + |
| 30 | 桡足类无节幼虫 | Copepod Nauplius larva | 浮游幼虫 | + | + |
| 31 | 多毛类幼体 | Polychaeta larva | 浮游幼虫 | + | + |
| 32 | 长尾类幼体 | Macrura larva | 浮游幼虫 | + | + |
| 33 | 双壳类幼虫 | Bivalves larva | 浮游幼虫 | + | + |
| 34 | 短尾类大眼幼虫 | Megalopa larva | 浮游幼虫 | + | |
| 35 | 短尾类溞状幼虫 | Brachyura zoea larva | 浮游幼虫 | + | + |
| 36 | 腹足类幼体 | Gastropoda larva | 浮游幼虫 | + | + |
| 37 | 阿利玛幼虫 | Alima larva | 浮游幼虫 | + | + |
| 38 | 歪尾类幼体 | Anomura larva | 浮游幼虫 | + | + |
| 39 | 海星羽腕幼虫 | Bipinnaria larva | 浮游幼虫 | + | + |
| 40 | 海蛇尾长腕幼虫 | Ophiopluteus larva | 浮游幼虫 | + | + |
| 41 | 帚虫类幼虫 | Phoronis larva | 浮游幼虫 | + | + |
| 42 | 鱼卵 | Fish egg | 浮游幼虫 | + | + |
| 43 | 仔鱼 | Fish larva | 浮游幼虫 | + | |
| 注: “+”表示出现, 空白表示未出现。 | |||||
浅水Ⅰ型网采集到的浮游动物种类数略多于浅水Ⅱ型网, 浅水Ⅰ型网采集到成体27种、浮游幼虫14类, 浅水Ⅱ型网采集到成体24种、浮游幼虫12类, 浅水Ⅰ型网、浅水Ⅱ型网均采集到的浮游动物成体有22种, 浮游幼虫有12类; 浅水Ⅰ型网、浅水Ⅱ型网采集到的浮游动物种类组成方面, 桡足类和浮游幼虫在种类组成上占优势, 其次为水螅水母类。
2.3 优势种本次调查共出现浮游动物优势种成体12种, 幼体4类(表 3), 其中桡足类6种、枝角类1种、管水母类2种、毛颚类1种、被囊类2种, 桡足类在优势种种类组成上占优势。浅水Ⅰ型网采集到优势种9种/类, 浅水Ⅱ型网采集到优势种10种/类, 两种不同网型采集到的优势种有所不同, 均采集到的优势种有强壮箭虫、异体住囊虫、多毛类幼体, 其中浅水Ⅰ型网采集到优势种平均丰度明显高于浅水Ⅱ型网。浅水Ⅰ型网采集到优势种多为体型较大的浮游动物种类, 如五角水母、双生水母等, 浅水Ⅱ型网采集到优势种多为体型较小的浮游动物种类, 如小拟哲水蚤、太平洋纺锤水蚤、双毛纺锤水蚤、拟长腹剑水蚤、近缘大眼剑水蚤等。
| 中文名 | 拉丁文名或英文名 | 浅水Ⅰ型网 | 浅水Ⅱ型网 | |||||
| 出现频率/% | 优势度 | 平均丰度/(ind.·m–3) | 出现频率/% | 优势度 | 平均丰度/(ind. ·m–3) | |||
| 五角水母 | Muggiaea atlantica | 38 | 0.05 | 12.9 | ||||
| 双生水母 | Diphyes chamissonis | 63 | 0.05 | 6.9 | ||||
| 小拟哲水蚤 | Paracalanus parvus | 100 | 0.18 | 139.9 | ||||
| 背针胸刺水蚤 | Centropages dorsispinatus | 38 | 0.05 | 19.5 | ||||
| 太平洋纺锤水蚤 | Acartia pacifica | 38 | 0.02 | 108.8 | ||||
| 双毛纺锤水蚤 | Acartia bifilosa | 88 | 0.03 | 65.7 | ||||
| 拟长腹剑水蚤 | Oithona similis | 50 | 0.04 | 37.3 | ||||
| 近缘大眼剑水蚤 | Corycaeus affinis | 100 | 0.14 | 169.8 | ||||
| 鸟喙尖头溞 | Penilia avirostris | 50 | 0.07 | 10.9 | ||||
| 强壮箭虫 | Sagitta crassa | 88 | 0.04 | 5.6 | 100 | 0.09 | 61.1 | |
| 异体住囊虫 | Oikopleura dioica | 88 | 0.06 | 12.1 | 100 | 0.12 | 151.7 | |
| 长尾住囊虫 | Oikopleura lonicauda | 50 | 0.04 | 120.9 | ||||
| 桡足类无节幼虫 | Copepodite Nauplius larva | 100 | 0.02 | 22.1 | ||||
| 多毛类幼体 | Polychaeta larva | 88 | 0.03 | 3.7 | 88 | 0.03 | 22.1 | |
| 长尾类幼虫 | Macrura larva | 75 | 0.04 | 7.5 | ||||
| 鱼卵 | Fish egg | 63 | 0.03 | 3.9 | ||||
| 注: 空白表示未出现。 | ||||||||
浮游动物平均生物量(湿质量)为69.5 mg/m3, 其中浅水Ⅰ型网平均生物量(湿质量)为37.7 mg/m3, 范围是4.1~115.8 mg/m3, 最低生物量出现在A8号站, 最高出现在A1号站(图 2a); 浅水Ⅱ型网平均生物量(湿质量)为101.3 mg/m3, 范围是19.2~ 365.6 mg/m3, 最低生物量出现在A8号站, 最高出现在A3号站(图 2b)。从平面分布看, 调查海域浮游动物生物量总体呈近岸高于外海, 北部高于南部的趋势。
 |
| 图 2 浮游动物生物量平面分布图(单位: mg/m3) Fig. 2 Horizontal distribution of zooplankton biomass |
浮游动物丰度平均为569.7 ind./m3, 其中浅水Ⅰ型网平均丰度为128.4 ind./m3, 范围是2.0~320.9 ind./m3, 最低丰度出现在A8号站, 最高出现在A1号站(图 3a); 浅水Ⅱ型网平均丰度为1 010.9 ind./m3, 范围是15.0~ 3 556.3 ind./m3, 最低丰度出现在A6号站, 最高出现在A3号站(图 3b)。从平面分布看, 调查海域浮游动物丰度总体呈近岸高于外海, 北部高于南部的趋势, 与生物量分布趋势基本一致。
 |
| 图 3 浮游动物丰度平面分布图(单位: ind./m3) Fig. 3 Horizontal distribution of zooplankton abundance |
对利用浅水Ⅰ型网和浅水Ⅱ型网调查采集的浮游动物数据进行多样性指数计算, 浅水Ⅰ型网调查采集的浮游动物多样性指数(H′)是2.00~3.75, 平均为2.87; 浅水Ⅱ型网是2.40~3.35, 平均为2.92, 平面分布情况见图 4。浅水Ⅰ型网调查浮游动物均匀度指数(J)是0.47~0.93, 平均为0.54; 浅水Ⅱ型网是0.60~0.92, 平均为0.70, 平面分布情况见图 5。浅水Ⅰ型网调查浮游动物丰富度指数(D)是1.34~3.04, 平均为2.18; 浅水Ⅱ型网是1.61~2.56, 平均为2.11, 平面分布情况见图 6。水平分布上浅水Ⅰ型网调查多样性指数(H′)近岸高于远海, 浅水Ⅱ型网调查近岸低于远海; 浅水Ⅰ型网和浅水Ⅱ型网调查均匀度指数(J)总体呈北部调查海域低于南部的分布趋势; 浅水Ⅰ型网和浅水Ⅱ型网调查丰富度指数(D)总体呈北部调查海域高于南部的分布趋势。浅水Ⅰ型网调查平均均匀度指数(J)、丰富度指数(D)略高于浅水Ⅱ型网调查, 多样性指数(H′)略低于浅水Ⅱ型网调查, 两种网具调查结果在群落多样性方面看有一定的差异。
 |
| 图 4 浮游动物多样性指数(H′)平面分布 Fig. 4 Horizontal distribution of Shannon-Weaver (H′) |
 |
| 图 5 浮游动物均匀度指数(J)平面分布 Fig. 5 Horizontal distribution of Pielou's evenness index (J) |
 |
| 图 6 浮游动物丰富度指数(D)平面分布 Fig. 6 Horizontal distribution of Margalef's species richness diversity (D) |
依据陈清潮等[15]对生物群落丰富程度的分级, 浅水Ⅰ型网采集调查海域浮游动物多样性阈值平均为2.2, 多样性评价等级为Ⅲ级, 等级描述多样性较好; 浅水Ⅱ型网采集调查海域浮游动物多样性阈值平均为2.1, 多样性评价等级为Ⅲ级, 等级描述多样性较好; 但两种网具采集浮游动物在各调查站位的多样性等级评价结果上有一定的差异。各站位评价结果见表 4。
| 站位 | 浅水Ⅰ型网 | 浅水Ⅱ型网 | |||||||
| 多样性指数(H′) | 均匀度(J) | 阈值(DV) | 等级描述 | 多样性指数(H′) | 均匀度(J) | 阈值(DV) | 等级描述 | ||
| A1 | 3.75 | 0.74 | 2.8 | 多样性丰富 | 3.35 | 0.71 | 2.4 | 多样性较好 | |
| A2 | 2.00 | 0.47 | 0.9 | 多样性一般 | 3.12 | 0.66 | 2.1 | 多样性较好 | |
| A3 | 3.42 | 0.76 | 2.6 | 多样性丰富 | 2.85 | 0.60 | 1.7 | 多样性较好 | |
| A4 | 2.74 | 0.67 | 1.8 | 多样性较好 | 2.49 | 0.59 | 1.5 | 多样性一般 | |
| A5 | 2.71 | 0.69 | 1.9 | 多样性较好 | 2.40 | 0.60 | 1.4 | 多样性一般 | |
| A6 | 2.49 | 0.83 | 2.1 | 多样性较好 | 3.31 | 0.92 | 3.0 | 多样性丰富 | |
| A7 | 3.46 | 0.89 | 3.1 | 多样性丰富 | 2.87 | 0.75 | 2.2 | 多样性较好 | |
| A8 | 2.41 | 0.93 | 2.2 | 多样性较好 | 2.99 | 0.79 | 2.4 | 多样性较好 | |
| 平均 | 2.87 | 0.75 | 2.2 | 多样性较好 | 2.92 | 0.70 | 2.1 | 多样性较好 | |
根据各调查站位浮游动物种类组成进行CLUSTER聚类和MDS标序分析, 结果见图 7和图 8。各调查站位之间浮游动物群落相似性较低, 在14%的相似度上浅水Ⅰ型网采集的浮游动物可划分为2个群落, 群落Ⅰ包含A1、A2、A3站位, 群落Ⅱ包含其余站位, MDS图中压力系数为0.03, 结果可信。在20%的相似度上浅水Ⅱ型网采集的浮游动物划分为2个群落, 群落Ⅰ包含A6、A7、A8站位, 群落Ⅱ包含其余站位, MDS图中压力系数为0.05, 结果可信。根据聚类分析结果(见图 7), 浅水Ⅰ型、浅水Ⅱ型网采集浮游动物的调查站位均划分为2个聚类群落, 但不同网具采集的浮游动物划分的2个群落所包含的站位不完全一致, 结果有一定差异, 聚类结果区别在A4、A5站位的归属。
 |
| 图 7 浮游动物群落种类组成CLUSTER聚类结果 Fig. 7 CLUSTER results of zooplankton community |
 |
| 图 8 浮游动物群落种类组成MDS标序分析结果 Fig. 8 MDS analysis results of zooplankton community |
浮游动物种类数量和种类组成是生物群落的基本组成成分, 通过对比不同年度物种数的变化, 可以在一定程度上了解该海域浮游动物的群落变化趋势。将本研究调查结果与青岛近岸海域的历史调查资料进行对比, 见表 5。与2006年7月青岛近海调查数据[1]相比, 青岛近岸海域浮游动物种类数有所减少; 平均丰度和平均生物量有所降低; 多样性指数所有升高。与2006年8月青岛南部近岸调查数据[9]相比, 青岛近岸海域浮游动物种类数有所减少; 平均丰度和多样性指数有所升高; 平均生物量有所降低。与2016年9月青岛近岸调查数据[2]相比, 青岛近岸海域浮游动物种类数有所增加; 多样性指数有所升高。造成这种差异的原因主要有以下几个方面: (1) 采样站位的布设位置和数量不同, 不同的采样站位所在位置海域的环境水文和海水水质条件不同, 一般情况下, 采样站位数量越多采集到浮游动物的种类数量越多。(2) 采样网具不同, 浅水Ⅱ型浮游生物网衣孔径为0.160 mm, WP2型浮游生物网衣孔径为0.200 mm, 浮游生物大网和浅水Ⅰ型浮游生物网衣孔径为0.505 mm, 浅水Ⅱ型浮游生物网和WP2型浮游生物网衣孔径与浮游生物大网和浅水Ⅰ型浮游生物网相比, 网衣孔径更小, 该型网能采集更多的小型浮游生物, 因此造成该网型采集的浮游动物丰度和生物量较高。(3) 调查季节的差异, 几次调查的季节有所不同, 不同季节的调查结果有所不同。
多样性指数除了可以用来描述生物群落的多样性特征, 还可以用于评价海洋环境质量、判定海洋环境受污染程度。一般分为以下几个层次, H′值小于1, 属于重污染; H′值为1~2, 属于中等污染; H′值为2~3, 属于轻度污染; H′值大于3, 属于清洁[18]。浅水Ⅰ型网和浅水Ⅱ型网调查浮游动物平均H′分别为2.87和2.92, 根据评判标准[18], 调查海域环境为轻度污染, 浅水Ⅰ型网调查结果除A2、A4、A5、A6、A8站多样性指数为2~3, 属于轻度污染, 其余调查站位均高于3, 属于清洁。浅水Ⅱ型网调查结果除A3、A4、A5、A7、A8多样性指数为2~3, 属于轻度污染, 其余调查站位均高于3, 属于清洁。浅水Ⅰ型网和浅水Ⅱ型网调查A4、A5、A8站多样性指数(H′)、丰富度指数(D)较低, 原因可能是A4、A5站位邻近青岛港, A8站邻近董家口港区, 受港区航行船舶扰动的影响, 导致这几个站位多样性指数(H′)、丰富度指数(D)偏低。
3.3 两种网具调查结果的比较本次调查利用浅水Ⅰ型网和浅水Ⅱ型网进行浮游动物的调查, 在采集到浮游动物种类数量方面, 从整个调查海域分析浅水Ⅰ型网采集的种类数量略多于浅水Ⅱ型网, 采样效率基本一致, 从单个站位上看, 除A1、A7号站浅水Ⅰ型网采集的种类数量多于浅水Ⅱ型网外, 其余站位浅水Ⅰ型网采集的种类数量均少于浅水Ⅱ型网。在丰度和生物量方面, 无论是整个调查海域还是单个站位, 浅水Ⅱ型网的采集的浮游动物丰度和生物量均远大于浅Ⅰ型网。通过对比分析数据后发现, 浅水Ⅱ型网不光在对小拟哲水蚤、双毛纺锤水蚤、太平洋纺锤水蚤、拟长腹剑水蚤、近缘大眼剑水蚤、双壳类幼虫等小型浮游动物, 同样对于强壮箭虫等大型浮游动物, 在数量采集上浅水Ⅱ型网远高于浅水Ⅰ型网。目前, 国内许多关于浮游动物的研究报道大都以浅水Ⅰ型浮游生物网的调查结果为主, 其结果分析偏重于大中型浮游动物的研究, 相对地, 小型浮游动物却没有受到足够的重视[19]。随着海洋生态系统的进一步深入研究, 小型浮游动物作为次级生产力的重要组成部分, 在海洋生态系统中也发挥同样重要的作用[20]。因此, 要深入了解小型浮游动物在海洋环境中的分布规律, 应当重视浅水Ⅱ型浮游生物网采集结果。有研究者认为, 中小型浮游动物在海洋生态系统中具有不可替代的生态功能, 仅利用浅水Ⅰ型浮游生物网采集浮游动物, 其数据结果将导致小型浮游动物实际数量严重偏低[21], 本研究结果得到了同样的结论。有研究结果表明, 由于人类开发活动等因素的影响, 许多海域水体中的浮游动物发生了一定的变化, 主要表现为, 大中型种类数量逐渐减少, 小型个体逐渐增多, 整个浮游动物群落朝向小型化发展趋势[22]。在这种发展趋势下, 如果仅采用浅水Ⅰ型浮游生物网进行浮游动物的采集, 研究结果必将低估该调查海域浮游动物中小型种类的数量, 造成该海域浮游动物丰度和生物量结果偏低。有研究表明, 在对大中型浮游动物采样效率大致相等的前提下, 在描述小型浮游动物的分布状况方面, 浅水Ⅱ型浮游生物网的数据更为准确[23]。
一般情况下, 浮游动物群落结构分析并不会因为所使用的网具不同而发生大的改变[24], 但本次研究结果表明不同网具采集的浮游动物聚类分析结果存在一定的差异, 浅水Ⅰ型网采集的浮游动物可划分为群落Ⅰ(A1、A2、A3站位), 群落Ⅱ(A4、A5、A6、A7、A8站位), 浅水Ⅱ型网采集的浮游动物划分为群落Ⅰ(A1、A2、A3、A4、A5站位), 群落Ⅱ(A6、A7、A8站位), 聚类结果区别在A4、A5站位的归属, 分析原因可能是浅水Ⅱ型网采集A4、A5站位与A1、A2、A3站位在浮游动物优势种数量上相似性较高造成的。
因此, 在今后浮游动物调查中, 根据调查需要可同时利用2种浮游生物网具进行浮游动物的采集, 综合考虑不同网具的浮游动物调查结果。同时利用2种网具进行调查, 有利于采集到更多种类, 从而获得更加全面的调查结果。
| [1] |
王晓, 王宗灵, 刘萍, 等. 夏季青岛近海浮游动物种类组成、群落结构及多样性[J]. 海洋科学进展, 2009, 27(3): 376-383. WANG Xiao, WANG Zongling, LIU Ping, et al. Species composition, community structure and diversity of zooplankton in Qingdao coastal area in summer[J]. Adva n ces in Marine Science, 2009, 27(3): 376-383. DOI:10.3969/j.issn.1671-6647.2009.03.011 |
| [2] |
王宁, 庄昀筠, 刘光兴, 等. 青岛近海浮游动物的群落特征研究[J]. 海洋环境科学, 2021, 40(2): 190-199. WANG Ning, ZHUANG Yunyun, LIU Guangxing, et al. Zooplankton community structures of Qingdao coastal water[J]. Marine Environmental Science, 2021, 40(2): 190-199. |
| [3] |
沈国英, 施并章. 海洋生态学[M]. 第二版. 北京: 科学出版社, 2002. HEN Guoying, SHI Bingzhang. Marine ecology[M]. Second edition. Beijing: Science Press, 2002. |
| [4] |
陈学超, 朱丽岩, 黄瑛, 等. 南黄海浮游动物群落结构研究[J]. 海洋科学, 2017, 41(10): 41-49. CHEN Xuechao, ZHU Liyan, HUANG Ying, et al. Community structure of the zooplankton in the Southern Yellow Sea[J]. Marine Sciences, 2017, 41(10): 41-49. |
| [5] |
孙松, 李超伦, 张光涛, 等. 胶州湾浮游动物群落长期变化[J]. 海洋与湖沼, 2011, 42(5): 625-631. SUN Song, LI Chaolun, ZHANG Guangtao, et al. Long- term changes in the zooplankton community in the Jiaozhou bay[J]. Oceanologia et Limnologia Sinica, 2011, 42(5): 625-631. |
| [6] |
周青云, 宋微, 刘永清, 等. 2003年胶州湾红岛海域生物现状的初步研究Ⅱ——浮游动物现状的初步调查与研究[J]. 海洋科学, 2005, 26(6): 80-83. ZHOU Qingyun, SONG Wei, LIU Yongqing, et al. Preliminary research in 2003 on biological resources near hongdao island of Jiaozhou Bay—Ⅱ—zooplankton[J]. Marine Sciences, 2005, 26(6): 80-83. |
| [7] |
黄凤鹏, 孙爱荣, 王宗灵, 等. 胶州湾浮游动物的时空分布[J]. 海洋科学进展, 2010, 28(3): 332-341. HUANG Fengpeng, SUN Airong, WANG Zongling, et al. Spatial and temporal distribution of zooplankton in the Jiaozhou Bay[J]. Advances in Marine Science, 2010, 28(3): 332-341. DOI:10.3969/j.issn.1671-6647.2010.03.009 |
| [8] |
时永强, 左涛, 袁伟, 等. 夏季崂山湾浮游动物群落结构及空间分布特征[J]. 海洋与湖沼, 2017, 48(5): 990-997. SHI Yongqiang, ZUO Tao, YUAN Wei, et al. Community structure and spatial distribution of zooplank in Laoshan Bay in summer[J]. Oceanologia et Limnologia Sinica, 2017, 48(5): 990-997. |
| [9] |
寇建山. 青岛南部近海浮游动物群落结构研究[D]. 青岛: 中国海洋大学, 2011. KOU Jianshan. Study on the zooplankton community in offshore waters of south Qingdao[D]. Qingdao: Ocean University of China, 2011. |
| [10] |
国家质量监督检验检疫总局, 国家标准化管理委员会. 海洋监测规范: 第7部分近海污染生态调查和生物监测: GB17378.7—2007[S]. 北京: 中国标准出版社, 2008. General Administration of Quality Supervision, Inspection and Quarantine of the PRC, Standardization Administration of the PRC. The specification for marine monitoring-Part 7: Ecological survey for off shore pollution and biological monitoring: GB17378.7-2007[S]. Beijing: China Standard Press, 2008. |
| [11] |
徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 1989, 8(4): 13-15. XU Zhaoli, CHEN Yaqu. Aggregated intensity of dominant species of zooplankton in autumn in the East China Sea and Yellow Sea[J]. Journal of Ecology, 1989, 8(4): 13-15. |
| [12] |
SHANNON C E, WEAVER W. The mathematical theory of communication[M]. Urbana, IL: The University of Illinois Press, 1949, 125.
|
| [13] |
MARGALEF R. Information theory in ecology[J]. Ge neral systematics, 1958(3): 36-71. |
| [14] |
PIELOU E C. An introduction to mathematical ecology[M]. New York: Wiley-Interscience, 1969: 1-286.
|
| [15] |
陈清潮, 黄良民, 尹健强, 等. 南沙群岛海区浮游动物多样性研究[M]//中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物多样性研究I. 北京: 海洋出版社, 1994: 42-50. CHEN Qingchao, HUANG Liangmin, YIN Jianqiang, et al. Studies on the zooplanktonic biodiversities in the waters around Nansha islands[M]// Nansha Comprehensive Scientific Investigation Team, Chinese Academy of Sciences. Studies on marine biodiversity of the Nansha Islands and neighbouring waters I. Beijing: China Ocean Press, 1994: 42-50. |
| [16] |
周红, 张志南. 大型多元统计软件PRIMER的方法原理及在底栖群落生态学中的应用[J]. 青岛海洋大学学报, 2003, 33(1): 58-64. ZHOU Hong, ZHANG Zhinan. Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J]. Journal of Ocean University of Qingdao, 2003, 33(1): 58-64. |
| [17] |
CLARKE K R, GORLEY R N. PRIMER v5: User Manual/Tutorial[M]. Plymouth: PRIMER-E Ltd. 2001.
|
| [18] |
蔡立哲, 马丽, 高阳, 等. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报(自然科学版), 2002, 41(5): 641-646. CAI Lizhe, MA Li, GAO Yang, et al. Analysis on assessing criterion for polluted situation using soecies diversity index of marine macrofauna[J]. Journal of Xia men University (natural science), 2002, 41(5): 641-646. |
| [19] |
徐潇峰. 乳山湾及邻近海域浮游动物生态学研究[D]. 青岛: 中国海洋大学, 2013. XU Xiaofeng. Study on the zooplankton community in Rushan Bay and adjacent sea area[D]. Qingdao: Ocean University of China, 2013. |
| [20] |
张武昌, 张翠霞, 肖天. 海洋浮游生态系统中小型浮游动物的生态功能[J]. 地球科学进展, 2009, 24(11): 1195-1201. ZHANG Wuchang, ZHANG Cuixia, XIAO Tian. Role of microzooplankton in marine planktonic ecosystem[J]. Advances in Earth Science, 2009, 24(11): 1195-1201. |
| [21] |
王荣, 张鸿雁, 王克, 等. 小型桡足类在海洋生态系统中的功能作用[J]. 海洋与湖沼, 2002, 33: 453-460. WANG Rong, ZHANG Hongyan, WANG Ke, et al. Function performed by small copepods in marine ecosystem[J]. Oceanologia et Limnologia Sinica, 2002, 33: 453-460. |
| [22] |
王友绍, 王肇鼎, 黄良民. 近20年来大亚湾生态环境的变化及其发展趋势[J]. 热带海洋学报, 2004, 23(5): 85-95. WANG Youshao, WANG Zhaoding, HUANG Liangmin. Environment changes and trends in daya bay in recent 20 years[J]. Journal of Tropical Oceanography, 2004, 23(5): 85-95. |
| [23] |
朱艺峰, 王银, 林霞, 等. 象山港两种网目网采浮游动物群落比较[J]. 应用生态学报, 2012, 23(8): 2277-2286. ZHU Yifeng, WANG Yin, LIN Xia, et al. A comparison of zooplankton communities collected by two types of nets with different mesh sizes in Xiangshan Bay of Zhejiang, East China[J]. Chinese Journal of Applied Ecology, 2012, 23(8): 2277-2286. |
| [24] |
连喜平, 谭烨辉, 刘永宏, 等. 两种浮游生物网对南海北部浮游动物捕获效率的比较[J]. 热带海洋学报, 2013, 32(3): 33-39. LIAN Xiping, TAN Yehui, LIU Yonghong, et al. Comparison of capture efficiency for zooplankton in the northern South China Sea, using two plankton mesh sizes[J]. Journal of Tropical Oceanography, 2013, 32(3): 33-39. |