 2015, Vol. 46
2015, Vol. 46文章信息
- 汪洋, 吴常文. 2015.
- WANG Yang, WU Chang-Wen. 2015.
- 中街山列岛岩礁海域鱼类群落多样性研究
- FISH COMMUNITY DIVERSITIES IN REEF WATERS OF ZHONGJIESHAN ISLANDS
- 海洋与湖沼, 46(4): 776-785
- Oceanologia et Limnologia Sinica, 46(4): 776-785.
- http://dx.doi.org/10.11693/hyhz20150100003
-
文章历史
- 收稿日期:2015-01-05
- 改回日期:2015-02-11




群落多样性是构成群落的物种多样性在时间和空间上的反应,它是把物种数、丰富度和均匀度等结合起来考虑的统计方法,在群落物种稳定性研究,比较群落历史演替情况和人类活动对群落生态的影响等方面研究具有重要意义(李涛,2010)。从种类组成特征研究群落多样性,迄今已有较长的历史,方法也相对比较成熟。鱼类群落多样性研究方面已有大量的报道(朱鑫华等,1996; 李圣法等,2007; 姜亚洲等,2008; 王迎宾等,2012),但这些研究基本上是对开阔海域的鱼类群落进行研究,对各海域岩礁区鱼类群落多样性的系统研究报道较少,在近些年加大增殖放流规模和实施渔业资源修复工程等专项行动前提下,亟需加快岛礁鱼类资源群落多样性的历史和现状调查,为其可持续发展提供理论支持。
岩礁海域生境具有很高的生物多样性和群落稳定性,是一种不可缺失的自然生态系统(Taylor,1998)。不但是饵料生物的良好附着基,也成为了鱼类群落繁殖、发育、摄食和躲避敌害等的“天然良港”,形成了多样化的鱼类群落。Guidetti(2000)认为有丰富海藻生长繁殖的岩礁海域比没有海藻场的泥质和沙质生境有更高的鱼类群落多样性。Barros等(2001)认为一些生物的栖息密度会随着距离岩礁生境的远近呈梯度分布。意大利、西班牙、英国和澳大利亚等沿岸国家均对岩礁生境的鱼类多样性进行了多方面的研究(Underwood et al,1996; Taylor,1998; Azzurro et al,2010; De Raedemaecker et al,2010)。在国内,对南海重要珊瑚礁过渡性水域软骨鱼(陈国宝等,2006),西沙群岛主要岛礁鱼类物种多样性(孙典荣等,2005; 王雪辉等,2011),枸杞岛海藻场优势种鱼类群体(章守宇等,2011)等有过探索性报道,但研究的时间尺度鲜有包括全年所有月份的,难以研究岩礁鱼类对生境的交错利用的特点,在这方面只有马鞍列岛岩礁生境鱼类群落研究中(汪振华等,2012)有所涉及,但整体上看,岩礁海域生境鱼类资源调查在时间和空间尺度上都还比较缺乏,这跟岩礁海域鱼类资源亟待保护的现实需求存在很大的差距。
中街山列岛位于浙江舟山群岛东部,介于东海岱衢洋和黄大洋之间,处于舟山渔场的中心区域,历史上有“东海鱼仓”的美誉,2006年5月经批准设立了国家级海洋生态自然保护区。保护区内岛礁星罗棋布,水道错综复杂,常年受到台湾暖流和黄海水团的影响以及钱塘江和长江径流东流控制,营养丰富的饵料物质得以在中街山列岛岩礁海域聚集,成为我国主要经济鱼类重要的栖息、生长繁殖场所和必经洄游通道。但过去对该岩礁海域的相关研究多见于水质环境(阳丹等,2012)、海藻(王志铮等,2002)、产卵场(唐逸民等,1984)、甲壳类、虾蟹类(徐开达等,2012a,b,c)、底栖动物和游泳动物整体等(张龙等,2012; 刘小雄等,2014),专门的鱼类资源调查较少,仅有梁君等(2013,2014)对该海域的鱼类群落有过专门的调查研究,但调查研究的时间尺度和渔获网具都还有很大的局限性。本文基于中街山列岛岩礁海域2013年11月—2014年10月每月的流刺网和延绳钓渔获数据,对鱼类种类组成、群落多样性、迁移指数和受干扰程度等进行分析研究,填补该岩礁海域两种网具同步调查作业的鱼类资源数据空白,丰富中街山列岛岩礁海域鱼类群落生态学内容,探究岩礁海域生境生态学功能,为该海域鱼类资源的养护修复、可持续利用和有效管理提供科学依据,为维系生物群落多样性和实现整个海域的生态平衡提供理论支持。
1 材料与方法 1.1 研究区域和站点分布2013年11月至2014年10月采用流刺网和延绳钓调查作业在中街山列岛岩礁海域开展鱼类资源调查,本次调查参照当地捕捞经验,并依据捕捞干扰最小化的采样原则,设置了6个延绳钓调查站点和7个流刺网调查站点(图 1)。
 |
| 图 1 调查站位分布图 Fig. 1 The deployment of stations for investigation |
本研究用流刺网和延绳钓进行随机采样,采样时间为每个月大潮汛前的2—4天,延绳钓调查船为浙普渔5699号,流刺网调查船为浙普渔5327号。延绳钓采用多股胶丝绳,主纲为10mm左右,放钓的长度随着放钓的数量变动,基本维持在80—100m,总共有70—80枚钓钩,选用质量较好的不锈钢材质,支绳长1.0m,钓钩与钓钩间的距离为1.2m; 每个钓钩的宽度为20—30mm,长度为30—60mm左右。三重刺网为本研究选用的流刺网,单片网衣的宽度为1.2m,长度为75m,网目 尺寸5cm网衣在中间,尺寸20cm的网衣在两侧,一组共5片。渔获鱼类鉴定到种,样品分析按国家质检总局2007年的《海洋调查规范》(GB/T12763.6-2007)进行,记录渔获物种类、尾数和质量,质量精确到1g。
1.3 数据处理和统计分析为了深入分析研究海域鱼类群落的生态功能,渔获鱼类按适温性、分布水层、利用岩礁生境的空间尺度大小、利用岩礁生境的时间跨度和出现率分析其生态类型(汪振华等,2012)。
本研究采用相对重要性指数(index of relative importance,IRI)(Pinkas et al,1971)分析每个月出现的优势种。研究海域鱼类群落多样性分别采用Margalef(1968)种类丰富度指数(D),Pielou(1966)均匀度指数(J′)和基于鱼类尾数的Shannon-Wiener多样性指数(H′)(Whilm,1968)分析。使用更替指数(AI)和迁移指数(MI)分析群落的稳定性,利用ABC曲线对岩礁海域鱼类群落受到的外界干扰状况进行评价。
各指数表达式如下:
| IRI=100×(Pi+Wi)×Fi | (1) |
| D=(S-1)/lnN | (2) |
| H′=$ - \sum\limits_{i = 1}^s {} $Pi lnPi | (3) |
| J′= H′/ln S | (4) |
| AI=100×(C+B)/(A-B) | (5) |
| MI=100×(C-B)/(A-B) | (6) |
上述各式中,Pi代表第i种渔获鱼类尾数占总尾数的比例,Wi代表第i种渔获鱼类质量占总质量的比例。Fi代表第i种渔获鱼类出现的站点数占总调查站点数的比例,S代表渔获鱼类种类数,N代表渔获鱼类总尾数,W代表渔获鱼类总质量,A、B和C分别代表所调查月份中渔获鱼类总种类数、迁出鱼类数和新迁入鱼类数。本文将IRI≥10,且单次采样样本数≥10的鱼类定为优势种。
为了了解研究区域的鱼类渔获效率,本研究把流刺网和延绳钓两种捕捞方式分别标准化,用渔获鱼类尾数和质量两种方式计算单位捕捞努力量渔获量(CPUE)。由于不同渔具努力量的计算方式差异,本研究将1km作为流刺网作业的单位捕捞努力量,100钓作为延绳钓的单位捕捞努力量。另外为了区分按尾数和按质量计算的单位捕捞努力量渔获量,通过前者计算的单位捕捞努力量渔获量,按照延绳钓和流刺网分别称为上钓率(尾/100钓)和渔获率(尾/km),通过后者计算的单位捕捞努力量渔获量称作CPUE,分别记为kg/100钓和kg/km(孙典荣等,2006)。
2 结果 2.1 岩礁海域鱼类种类组成和月变化本次调查共采集鱼类5052尾,总生物量1704059g,隶属9目28科42属,共计49种(表 1)。采集到的硬骨鱼纲鲈形目种类达26种,占总渔获种类的53.06%,在分类学中占有绝对的优势; 然后鲱形目以6种占第二位,占总渔获鱼类种数的12.24%; 鲽形目和鲱形目各以5种,并列第三位,相应各占10.20%; 其余渔获种类相对较少,鲀形目和鳗鲡目各2种分别占4.08%,灯笼鱼目、鲇形目和鲻形目各1种,分别占总渔获种类的2.04%。
| 目 Order | 科 Family | 属 Genus | 种 Species |
| 灯笼鱼目 Myctophiformes | 1 | 1 | 1 |
| 鲽形目 Pleuronectiformes | 3 | 4 | 5 |
| 鲱形目 Clupeiformes | 2 | 5 | 6 |
| 鲈形目 Perciformes | 12 | 21 | 26 |
| 鳗鲡目 Anguilliformes | 2 | 2 | 2 |
| 鲇形目 Siluriformes | 1 | 1 | 1 |
| 鲀形目 Tetraodontiformes | 2 | 2 | 2 |
| 鲉形目 Scorpaeniformes | 4 | 5 | 5 |
| 鲻形目 Mugiliformes | 1 | 1 | 1 |
从不同生态类型来看,中街山列岛岩礁海域全年采集暖水性鱼类25种,占总渔获种类的51.02%; 暖温性鱼类24种,占总渔获物的48.98%,没有采集到冷水性鱼类。在栖息水层上,全年渔获近底层鱼类26种,占总数的53.06%,底层和中上层鱼类分别为12和11种,各占24.49%和22.45%; 近底层鱼类季节变化规律明显,5—11月维持在15种左右,12—4月保持在9种左右; 中上层鱼类6—9月维持在7种左右,10—5月仅有2—3种左右,种类最少的在2—3月,每月分别采集到1种; 底层鱼类除11月、1月和2月维持在3—4种外,其余月份基本稳定在8种左右。从渔获鱼类对该生境利用的空间上分析,研究海域出现最少的为河口洄游性鱼类3种(占6.12%),其次为近海洄游性鱼类22种,约占44.90%,出现最多的是常年生活在该海域的定居性鱼类,总共有24种,占总渔获鱼类种类的48.98%。从鱼类对岩礁海域利用的时间上看,本研究获得常年种28种(占57.14%),季节性种类19种(占38.78%),偶见种2种(占4.08%)分别为褐蓝子鱼(Siganus fuscescens)和鳓(Ilisha elongata)。
2.2 岩礁海域优势种分析和群落稳定性
全年在研究区域渔获优势鱼类10种,如表 2所示,底层中,平鲉科的褐菖鲉(Sebastiscus marmoratus),康吉鳗科的星康吉鳗(Conger myriaster)和海鳗科的海鳗(Muraenesox talabonoides)共3种为优势种。其中褐菖鲉是每月渔获的优势种,也是全年的绝对优势种,其年相对重要性指数为26.76; 星康吉鳗和海鳗只在春秋季少数月份中成为优势种。近底层中,石首鱼科的黄姑鱼(Nibea albiflora)、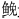 鱼(Miichthys miiuy)和小黄鱼(Pseudosciaena polyactis),鲷科的黑 棘鲷(Sparus macrocephlus)和
鱼(Miichthys miiuy)和小黄鱼(Pseudosciaena polyactis),鲷科的黑 棘鲷(Sparus macrocephlus)和 科的鲈鱼(Lateolabrax japonicus)5种为优势种。黄姑鱼是研究海域除7月以外的优势种,其年IRI=37.23,还远远高于褐菖鲉,成为全年中优势度最高的鱼类。鱼仅在1月、2月和7月未成为优势种,年相对重要性指数为22.39,仅次于褐菖鲉。小黄鱼、鲈鱼和黑棘鲷均只在个别月份有较大的捕渔获量。中上层优势鱼类主要是鳀科的赤鼻棱鳀(Thryssa kammalensis)和鲻科的鲻鱼(Mugil cephalus)2种。赤鼻棱鳀是近海洄游鱼类,仅在9月和10月成为优势种。鲻鱼在12月和1月的捕捞量明显高于其余月份。
科的鲈鱼(Lateolabrax japonicus)5种为优势种。黄姑鱼是研究海域除7月以外的优势种,其年IRI=37.23,还远远高于褐菖鲉,成为全年中优势度最高的鱼类。鱼仅在1月、2月和7月未成为优势种,年相对重要性指数为22.39,仅次于褐菖鲉。小黄鱼、鲈鱼和黑棘鲷均只在个别月份有较大的捕渔获量。中上层优势鱼类主要是鳀科的赤鼻棱鳀(Thryssa kammalensis)和鲻科的鲻鱼(Mugil cephalus)2种。赤鼻棱鳀是近海洄游鱼类,仅在9月和10月成为优势种。鲻鱼在12月和1月的捕捞量明显高于其余月份。
| 种名 | 栖息水层 | 月份 | |||||||||||
| 11 | 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 鲈鱼(Lateolabrax japonicus) | ▲ | 10.93 | 18.56 | 11.82 | 10.76 | ||||||||
| 小黄鱼(Pseudosciaena polyactis) | ▲ | 17.54 | 14.58 | 11.22 | |||||||||
| 黑鲷(Sparus macrocephlus) | ▲ | 20.47 | 17.78 | 11.40 | 11.99 | ||||||||
| 星康吉鳗(Conger myriaster) | ■ | 20.70 | 25.58 | ||||||||||
| 黄姑鱼(Nibea albiflora) | ▲ | 31.14 | 56.40 | 68.48 | 31.82 | 36.26 | 44.20 | 52.68 | 21.60 | 11.20 | 42.30 | 50.69 | |
| 褐菖鲉(Sebastiscus marmoratus) | ■ | 29.06 | 19.82 | 28.68 | 53.99 | 25.54 | 21.63 | 31.52 | 21.28 | 19.87 | 20.82 | 19.14 | 29.79 |
| 海鳗(Muraenesox cinereus) | ■ | 13.78 | |||||||||||
| 赤鼻棱鳀(Thryssa kammalensis) | ○ | 11.06 | 11.88 | ||||||||||
| | ▲ | 27.55 | 29.72 | 66.69 | 22.10 | 43.20 | 27.85 | 15.33 | 16.03 | 20.19 | |||
| 鲻鱼(Mugil cephalus) | ○ | 15.41 | 17.95 | ||||||||||
| 栖息水层三种符号的含义: ■. 底层鱼类,▲. 近底层鱼类,○. 中上层鱼类 | |||||||||||||
各月份更替指数(AI)和迁移指数(MI)如图 2所示,该年度群落稳定性最差的为1月,AI值高达700,7月份群落稳定性最高,AI值仅为13.33。11月、1月、2月、9月和10月MI值为负,说明这几个月迁出种大于迁入种,其中1月MI值的绝对值最大,说明温度改变对研究海域鱼类的迁移具有明显影响,其余月份MI值为正,迁入种大于迁出种。
 |
| 图 2 各月份渔获鱼类的更替指数和迁移指数Fig. 2 The alternation index and migration index of fish within each month reef waters of Zhongjieshan Islands |
由ABC曲线可知,就各月份鱼类群落受干扰程度而言,6月份生物量优势度曲线在数量优势度曲线之上,且W值为0.185,是所有月份中最大的,所以6月份鱼类群落受到的干扰最小,群落处于最稳定的状态。其余月份的W值除2月以外均为正值,说明其余月份鱼类群落未受到严重干扰,但我们也应清醒的看到,各个月份的W值均不大,2月、11月、5月、8月和10月等月份,生物量和数 量优势度曲线相交,说明鱼类群落受到了中等干扰,个别月份(2月)W值为负数,且生物量优势曲线几乎全部在数量优势度曲线之下,研究区域的鱼类群落有可能受到严重干扰。
 |
| 图 3 研究区域各月份鱼类群落的ABC曲线以及W统计值 Fig. 3 ABC curves and W value of fish community in study area each month |
流刺网和延绳钓作业CPUE分析如图 4所示,流刺网作业中渔获率7月最高,2月最低,分别为65.45尾/km,22.22尾/km,CPUE在9月份最高,2月份最低,分别为19.73 kg/km和4.82 kg/km,月变化趋势均呈现出随温度升高波动的上行变化,随温度降低波动 的下行变化。延绳钓作业上钓率和CPUE均在春季的3月最低,分别为4.03尾/100钓,1.33 kg/100钓,上钓率夏季的8月最高,为5.72尾/100钓,CPUE秋季的11月最高,为2.57 kg/100钓,月变化趋势方面上钓率是4个单位努力量捕获量中波动相对平稳的。
 |
| 图 4 中街山列岛岩礁海域流刺网和延绳钓作业CPUE月变化 Fig. 4 Monthly variations of CPUE of drift-net and long-lining in reef waters of Zhongjieshan Islands |
岩礁海域鱼类种类数和多样性月变化箱形图见图 5所示:(1)气温较低的冬季2月,渔获鱼类种类数最低,仅为12种,渔获鱼类种类数的变化范围也最小。最大值出现在夏季的8月,该月渔获种类达到32种。种类数月均值变化范围为8—21.5,气温较高的6—9月,渔获物种类明显高于其它月份,6月,随着气温升高,渔获物种类数有很明显的增加,到夏末秋初的9月,渔获物种类开始下降,下降幅度很大。(2)研究海域鱼 类丰富度2月份最低,为2.35。3—7月,丰富度不断增加,7月(4.93)达到最大,8月开始有所下降,9月起,丰富度下降最为明显,但气温较高的6—9月,鱼类的丰富度还是明显高于其余月份。(3)多样性(H′)最低值出现在11月的一次采样中,仅为1.40,最大值3.17出现在8月,月均值最大的出现在7—8月,为2.80左右。整体变化趋势上,随着气温下降多样性降低,气温升高多样性增高,但在12月和5月,多样性指数有波动现象发生。(4)本次调查研究海域鱼类群落的均匀度指数在2013年12月—2014年5月表现出微小波动的下行变化,然后略有增加,7月份达到最大,并在7月后开始下降,但从整体来看,均匀度指数变化幅度没有前3个指数明显。均匀度指数的最大值和最小值分别是0.97、0.74,均出现在2013年12月的调查中,这一个月均匀度指数变化范围最大。
 |
| 图 5 研究海域鱼类群落多样性指数月变化 Fig. 5 Box plots of monthly variation of fish community diversity index in study area |
作为一种不可替代的重要生态系统,岩礁海域的岛礁岩石,作为鱼类良好附着基和重要栖息场所,满足了岩礁鱼类群落生长繁殖、发育和躲避天敌等生
存需要,大量的岩礁鱼类群落因此在此生活(Barros,2005)。根据本研究结果,恋礁鱼类如褐菖鲉和星康吉鳗等为全年特征种,大泷六线鱼(Hexagrammos otakii)、斑头鱼(Agrammus agrammus)、海鳗和褐牙鲆(Paralichthys olivaceus)等在很多月份都有渔获。趋礁性鱼类黄姑鱼、黑鲷、鲈鱼、小黄鱼和![]() 鱼等也常年出没在该海域。蓝圆鲹(Decapterus maruadsi)、日本鳀(Engraulis japonicus)和赤鼻棱鳀(Thryssa kammalensis)等鱼类也一般在夏季进入该岩礁海域,组成了丰富多彩的鱼类群落。可以归纳为: 定居种对岩礁海域周年依赖、洄游性鱼类对岩礁海域季节性利用和其它偶见种类的暂时性停留共同展现了岩礁海域的生态功能(汪振华等,2012)。从每月渔获的洄游性鱼类占总渔获种类比例来看,6—10月渔获的洄游性鱼类相对较高,均达到40%以上,7月达到最高为48.39%,2月份最低,仅为8.33%,由此可见,夏季由于洄游性鱼类的到来,岩礁海域生境利用率达到最大。
鱼等也常年出没在该海域。蓝圆鲹(Decapterus maruadsi)、日本鳀(Engraulis japonicus)和赤鼻棱鳀(Thryssa kammalensis)等鱼类也一般在夏季进入该岩礁海域,组成了丰富多彩的鱼类群落。可以归纳为: 定居种对岩礁海域周年依赖、洄游性鱼类对岩礁海域季节性利用和其它偶见种类的暂时性停留共同展现了岩礁海域的生态功能(汪振华等,2012)。从每月渔获的洄游性鱼类占总渔获种类比例来看,6—10月渔获的洄游性鱼类相对较高,均达到40%以上,7月达到最高为48.39%,2月份最低,仅为8.33%,由此可见,夏季由于洄游性鱼类的到来,岩礁海域生境利用率达到最大。
从优势鱼类角度来看,全年渔获的优势鱼类较少,而且几乎全部为常年种或定居性鱼类。刘小雄(2014)认为是由该海域洄游鱼类种类较多造成的,梁君等(2013)的研究进一步说明了季节性洄游种和偶见种即使在种类数量中占有很大的比例,但不到25%的种类在群落生物量中起主导作用。所以定居性鱼类的生态功能可以从侧面反应该海域生态功能,可以在以后的研究中进一步分析。
3.2 岩礁海域群落稳定性岩礁鱼类群落的稳定性受到多种复杂因素的影响,目前的研究聚焦在内因和外因两个方面: 内因又分抵抗力和群落种类的迁移。抵抗力是群落抵制或忍受变化的能力,群落结构越简单抵抗力越弱,主要采用多样性指数来反映; 群落种类的迁移,主要通过更替指数,也就是AI值来体现,AI 值越低群落种类迁移率越低,稳定性也就越高(金显仕等,2009); 外因主要是指群落受到的干扰程度,通过ABC曲线进行分析(李圣法,2008; 姜亚洲等,2013)。
就内因角度来说,研究海域鱼类群落D和H′值2月份最低,也就是说由于气温下降,鱼类多样性降低,鱼类群落抵抗力减弱,处于最不稳定的状态。相反,气温升高的6—9月,D和H′值均为全年中最大,鱼类群落抵抗力最强。AI值方面表现出同样的趋势,气温较低的1月AI值高达700,群落稳定性最差,气温较高的7月AI值仅为13.33,群落稳定性最高。从气温较低的冬季到气温最高的夏季,随着气温升高鱼类群落的稳定性升高。梁君等(2013)的研究也证实了暖水性鱼类与暖温性鱼类种类数与水温同步的事实。
鱼类群落受到的外在干扰,主要来自海洋环境污染,Warwick于1986年提出丰度/生物量比较法(ABC曲线法)监测受到的干扰。该方法原理比较简单、还具有简单便捷的统计计算过程、不需要任何对比参照点和结果分析简单快捷等优势(曲方圆等,2009),在全球范围内得到了普遍应用,具有良好的分析效果。本研究ABC曲线W值除2月以外均为正值,说明研究海域鱼类群落未受到严重干扰,但各个月份的W值均不大,部分月份鱼类群落受到了中等干扰,2月W值为负数,且生物量优势曲线几乎全部在数量优势度曲线之下,这很有可能是中街山列岛岩礁海域鱼类群落受到严重干扰的一个预警信号。特别是2014年以来,研究海域海岛旅游业的大力发展,海岛旅游人数的大量增加给该海域生态承受能力带来了巨大挑战,旅游经济利益驱使下的过度捕捞和海域污染加剧,势必会给鱼类群落带来更严重的干扰。
3.3 研究海域渔获水平为了比较本研究的渔获水平,现将已报道的孙典荣等(2005,2006)和麦有攀等(1999)等的流刺网和延绳钓调查的结果列于表 3、表 4。从流刺网作业来看,此次调查与1998年中沙、南沙调查相比,本调查的渔获率明显较高,但CPUE较低,说明此次调查中小型鱼类居多; 与2003年西沙和中沙调查相比,无论是渔获率还是CPUE,本次调查均偏低,除研究海域的丰度不及外,最有可能是跟中街山列岛岛礁鱼类资源量下降有关。从延绳钓作业来看,本次调查的上钓率为4.91尾/100钓,比1998—1999年南沙,2003年西沙和中沙岩礁海域调查都要高,但本调查的CPUE仅为1.78 kg/100钓,这比南沙、中沙和西沙的调查结果都低,而且差异性非常大,说明中街山岩礁海域延绳钓渔获鱼类个体较少,属于中小型鱼类,而1998—1999年南沙,2003年西沙和中沙调查渔获个体稍大,属于大中型鱼类。所以,研究海域亟需通过增大渔具网目、延长伏季休渔时间和严厉打击非法捕捞等行之有效的管理手段,使中小型鱼类有更充足的生长繁殖空间。
| 海区 | 年份 | 努力量(km) | 渔获尾数(尾) | 鱼获重量(kg) | 渔获率(尾/km) | CPUE(kg/km) |
| 中沙、南沙 | 1998 | 312.46 | 1357 | 19938.00 | 4.34 | 63.81 |
| 西沙 | 2003 | 14.48 | 3623 | 1055.45 | 250.21 | 72.89 |
| 中沙 | 2003 | 16.12 | 933 | 351.44 | 57.90 | 21.80 |
| 中街山 | This survey | 74.70 | 3351 | 1053.61 | 44.03 | 13.43 |
| 海区 | 年份 | 努力量(钓) | 渔获尾数(尾) | 鱼获重量(kg) | 上钓率(尾/100钓) | CPUE(kg/100钓) |
| 西沙 | 2003 | 3740 | 77 | 1711.55 | 2.06 | 45.76 |
| 中沙 | 2003 | 6060 | 134 | 2294.32 | 2.21 | 37.86 |
| 南沙 | 1998—1999 | 4420 | 82 | 1235.00 | 1.94 | 29.26 |
| 中街山 | This survey | 35560 | 1759 | 667.39 | 4.91 | 1.67 |
从多样性指数上来看,研究海域气温较高的7—8月D、H′和J′值普遍高于冬季,可能的原因是夏季长江流域雨水较多导致长江入海径流量增大,给包含中街山列岛岩礁海域在内的舟山群岛海域带来了丰富的营养物质,并随着低盐和高盐性的两种水系锋带的形成,岩礁海域鱼类群落赖以生存繁衍的饵料生物迅猛增长造成的。另外,多样性指数H′与温度呈现显著的正相关关系,使得该海域夏季不仅能为定居性鱼类,也为洄游性和偶见性鱼类提供了良
好的繁衍生长场所。研究结果显示,12月和5月的D、H′和J′值均有不同程度的波动,前者可能的原因是随着冬季黑潮暖流的到来,与研究海域的沿岸流形成的陆架混合水对鱼类群落起决定性作用,暖水种鱼类的大量补充以及一部分定居种、正在进行生殖洄游的鲻鱼等的生殖造成的。后者则可能由于暖水性鱼类离开该生境而蓝圆鲹、日本鳀和赤鼻棱鳀等夏季常见种还未集群到达造成的。比较近些年的类似报道,张龙等(2012)和梁君等(2013)用底拖网在中街山列岛岩礁海域分别渔获鱼类24种、55种,梁君等(2014)和刘小雄等(2014)用定置刺网在中街山列岛岩礁海域渔获鱼类58种、40种,岩礁海域生境承载的鱼类多样性仍然较高。但是,现有鱼类数与20世纪90年代中期中街山列岛海域记录的124种(吴常文等,1995)相比,已经出现了很大的下滑,这应该引起足够的重视。从本研究梁君等(2014)、刘小雄等(2014)对该海域的研究来看,虽然渔获了大量近年来增殖放流的黑鲷、黄姑鱼和大黄鱼等经济种类,但鱼类群落结构还是表现为鱼类小型化和低质化的特点,无论是底拖网、流刺网还是延绳钓作业,中小型鱼类已逐渐替代传统的大型经济鱼类成为优势种,因此需要进一步加快该海域的岛礁鱼类资源的保护,加大海洋牧场建设步伐,进行岛礁性经济鱼类的修复和养护,进一步有针对性的实施增殖放流、预防和治理海域污染和延长伏季休渔等措施,实现中街山列岛岩礁海域鱼类的资源的可持续发展。
4 结论本文对中街山群岛岩礁海域鱼类群落多样性的研究表明,定居种对岩礁海域周年依赖、洄游性鱼类对岩礁海域季节性利用和其它偶见种类的暂时性停留共同展现了该岩礁海域的生态功能,但中街山列岛岩礁海域中小型鱼类已逐渐替代传统的大型经济鱼类成为优势种,鱼类群落也有演变为严重干扰的趋势。因此,笔者认为有必要加强该岩礁海域的基础研究和岛礁鱼类资源的针对性保护,为实现海岛生态系统良性循环和可持续发展提供科技支撑。
| 王志铮, 张义浩, 吴常文等, 2002. 中街山列岛底栖海藻的资源调查. 水产学报, 26(2): 189—192 |
| 王迎宾, 俞存根, 陈全震等, 2012. 春、夏季舟山渔场及其邻近海域鱼类群落格局. 应用生态学报, 23(2): 545—551 |
| 王雪辉, 杜飞雁, 林昭进等, 2011. 西沙群岛主要岛礁鱼类物种多样性及其群落格局. 生物多样性, 19(4): 463—469 |
| 曲方圆, 于子山, 隋吉星等, 2009. 丰度生物量比较法应用局限性. 海洋科学, 33(6): 118—121 |
| 朱鑫华, 杨纪明, 唐启升, 1996. 渤海鱼类群落结构特征的研究. 海洋与湖沼, 27(1): 6—13 |
| 刘小雄, 王伟定, 梁 君等, 2014. 中街山列岛岛礁生境游泳动物群落多样性及稳定性分析. 浙江海洋学院学报(自然科学版), 34(1): 14—18, 30 |
| 孙典荣, 邱永松, 林昭进等, 2006. 中沙群岛春季珊瑚礁鱼类资源组成的初步研究. 海洋湖沼通报, (3): 85—92 |
| 孙典荣, 林昭进, 邱永松, 2005. 西沙群岛重要岛礁鱼类资源调查. 中国海洋大学学报, 35(2): 225—231 |
| 阳 丹, 陈应华, 吴常文, 2012. 中街山列岛及其邻近海域海水质量评价. 浙江海洋学院学报(自然科学版), 31(1): 49—53 |
| 麦有攀, 王弗良, 张光烂等, 1999. 中沙、南沙群岛礁盘深水域中下层鱼类资源开发利用技术研究. 中国水产, (3): 48—49 |
| 李 涛, 2010. 北黄海及山东半岛南部近岸海域渔业资源群落结构的初步研究. 青岛: 中国海洋大学硕士学位论文, 3—4 |
| 李圣法, 2008. 以数量生物量比较曲线评价东海鱼类群落的状况. 中国水产科学, 15(1): 136—144 |
| 李圣法, 程家骅, 严利平, 2007. 东海大陆架鱼类群落的空间结构. 生态学报, 27(11): 4377—4386 |
| 吴常文, 吕永林, 虞顺成, 1995. 中街山岛礁区渔业综合开发建议. 海洋开发与管理, 12(1): 68—71 |
| 汪振华, 章守宇, 陈清满等, 2012. 马鞍列岛岩礁生境鱼类群落生态学. I. 种类组成和多样性. 生物多样性, 20(1): 41—50 |
| 张 龙, 徐开达, 张洪亮等, 2012. 中街山海域渔业动物群落结构的季节变化. 浙江海洋学院学报(自然科学版), 31(4): 290—294 |
| 陈国宝, 李永振, 陈新军等, 2006. 南海重要珊瑚礁过渡性水域软骨鱼类的组成与分布. 上海水产大学学报, 15(4): 461—467 |
| 金显仕, 单秀娟, 郭学武等, 2009. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 29(9): 4761—4772 |
| 姜亚洲, 林 楠, 袁兴伟等, 2013. 象山港游泳动物群落结构及多样性. 生态学杂志, 32(4): 920—926 |
| 姜亚洲, 程家骅, 李圣法, 2008. 东海北部鱼类群落多样性及其结构特征的变化. 中国水产科学, 15(3): 453—459 |
| 徐开达, 卢占晖, 薛利建等, 2012a. 中街山列岛保护区蟹类群落结构特征的研究. 上海海洋大学学报, 21(6): 1032—1037 |
| 徐开达, 宋海棠, 张 龙等, 2012b. 中街山列岛保护区虾类资源状况及其与环境因子的关系. 浙江海洋学院学报(自然科学版), 31(5): 421—425 |
| 徐开达, 张洪亮, 谢汉阳等, 2012c. 中街山列岛水域甲壳类资源及其群落多样性. 海洋渔业, 34(3): 308—315 |
| 唐逸民, 郑佩玉, 李永明等, 1984. 中街山曼氏无针乌贼产卵场生态环境及其资源保护. 浙江水产学院学报, 3(2): 131—140 |
| 章守宇, 王 蕾, 汪振华等, 2011. 枸杞岛海藻场优势种鱼类群体特征及其在不同生境中的差异. 水产学报, 35(9): 1399—1409 |
| 梁 君, 王伟定, 虞宝存等, 2014. 中街山列岛海洋保护区岛礁生境鱼类资源及群落多样性季节变化. 海洋与湖沼, 45(5): 979—989 |
| 梁 君, 徐汉祥, 王伟定, 2013. 中街山列岛海洋保护区鱼类物种多样性. 生态学报, 33(18): 5905—5916 |
| Azzurro E, Matiddi M, Fanelli E et al, 2010. Sewage pollution impact on Mediterranean rocky-reef fish assemblages. Marine Environmental Research, 69(5): 390—397 |
| Barros F, 2005. Evaluating the importance of predation on subtidal benthic assemblages in sandy habitats around rocky reefs. Acta Oecologica, 27(3): 211—223 |
| Barros F, Underwood A J, Lindegarth M, 2001. The influence of rocky reefs on structure of benthic macrofauna in nearby soft-sediments. Estuarine, Coastal and Shelf Science, 52(2): 191—199 |
| De Raedemaecker F, Miliou A, Perkins Rupert, 2010. Fish community structure on littoral rocky shores in the Eastern Aegean Sea: Effects of exposure and substratum. Estuarine, Coastal and Shelf Science, 90(1): 35—44 |
| Guidetti P, 2000. Differences among fish assemblages associated with nearshore Posidonia oceanica seagrass beds, rocky-algal reefs and unvegetated sand habitats in the Adriatic Sea. Estuarine, Coastal and Shelf Science, 50(4): 515—529 |
| Margalef R, 1968. Perspectives in Ecological Theory. Chicago: University of Chicago Press |
| Pielou E C, 1966. The use of information theory in the study of ecological succession. Journal of Theoretical Biology, 10: 370—383 |
| Pinkas L, Oliphant M S, Iverson I L K, 1971. Food habits of albacore, bluefin tuna, and bonito in California waters. California Department of Fish and Game Fish Bulletin, 152: 1—105 |
| Taylor R B, 1998. Density, biomass and productivity of animals in four subtidal rocky reef habitats: the importance of small mobile invertebrates. Marine Ecology Progress Series, 172: 37—51 |
| Underwood A J, Chapman M G, 1996. Subtidal assemblages on rocky reefs at a cliff-face sewage outfall (North Head, Sydney, Australia): What happened when the outfall was turned off? Marine Pollution Bulletin, 33(7—12): 293—302 |
| Whilm J L, 1968. Use of biomass units in Shannon’s formula. Ecology, 49: 153—156 |