 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 卢少坤, 李荣华, 母昌考, 宋微微, 郑维兵, 王春琳. 2015.
- LU Shao-Kun, LI Rong-Hua, MU Chang-Kao, SONG Wei-Wei, ZHENG Wei-Bing, WANG Chun-Lin. 2015.
- 三疣梭子蟹(Portunus trituberculatus)低盐耐受性和体重的遗传参数估计及其相关分析
- ESTIMATION ON GENETIC PARAMETERS FOR LOW SALINITY TOLERANCE AND BODY WEIGHT OF PORTUNNUS TRITUBERCULATUS
- 海洋与湖沼, 46(5): 1040-1045
- Oceanologia et Limnologia Sinica, 46(5): 1040-1045.
- http://dx.doi.org/10.11693/hyhz20150500134
-
文章历史
- 收稿日期: 2015-05-05
- 收修改稿日期: 2015-06-03




2. 国家海洋局第三海洋研究所 厦门 361005
2. Third Institute of Oceanography, State Oceanic Administration, Xiamen 361005, China
三疣梭子蟹(Portunus trituberculatus)属于甲壳纲、梭子蟹科、梭子蟹属,广泛分布于我国沿海海区(戴爱云等,1977)。根据2013中国渔业统计年鉴(农业部渔业局,2013),其捕捞量达400348.0t,养殖面积较广,是我国重要的海产经济蟹类之一。三疣梭子蟹属于广盐性物种,适应盐度在10—35,最适生长盐度在20—35,但低盐刺激会对其生理生化会造成一定的影响,王冲等(2010)研究发现盐度骤降会使幼蟹的摄食率、变态率及存活率下降。由于其养殖区域大部分为户外池塘或滩涂围网养殖,水体环境盐度较低,且易受大量降水等因素影响,可能会造成其养殖水体在短时间内盐度过低,研究其生长及低盐耐受性,对于提高养殖存活率及扩大养殖面积有重要意义。
遗传参数主要包括遗传力、遗传相关与重复力,主要是涉及遗传力和遗传相关,其在个体遗传评定、选择反应预测、育种方案设计等中都具有广泛的用途。近些年来,随着水产养殖业的发展,国内外对物种遗传参数评估及家系选育上研究也发展迅猛,主要涉及于生长、抗性、肉质等(Fjalestad et al,1993),如虹鳟(Oncorhynchus mykiss)的体重性状遗传力分析(Aulstad et al,1972)、凡纳滨对虾(Litopenaeus vannamei)、虾夷扇贝(Patinopecten yessoensis)、刺参(Stichopus japonicas)、大菱鲆(Scophthalmus maximus)等的生长遗传力分析(Pérez-Rostro et al,1999; 栾生等,2006; 马爱军等,2008; 梁峻等,2011)、太平洋牡蛎(Crassostrea gigas)的肉质遗传力分析(Langdon et al,2000)。水产生物与抗逆性遗传研究也较多,如黄永春等(2013)构建了凡纳滨对虾抗WSSV选育家系通过家系选育后其不仅死亡率低还延迟了其死亡高峰时间,王庆志等(2014)构建了虾夷扇贝耐高温育种家系并提供了家系稚贝的早期筛查方法,严福升(2009)研究了牙鲆(Paralichthys olivaceus)的生长相关性状遗传力估计及生长性状与耐热性状的相关性分析。在三疣梭子蟹中对抗逆性遗传力的研究还未见报道。本研究构建了20对三疣梭子蟹家系,以急性低盐胁迫致死率为衡量指标,应用全同胞方差组分析法,评估其60日龄盐度耐受性遗传力及60日龄的体重遗传力,并分析生长性状与低盐耐受性状的相关性。为选育具有低盐耐受性的三疣梭子蟹提供理论基础。
1 材料与方法 1.1 亲本选择2013年9月在浙江宁波象山、宁海、奉化三地选择身体健康、规格均一、未交配的三疣梭子蟹作为亲蟹。
1.2 实验方法 1.2.1 亲蟹交尾2013年9月,挑选体格健壮、生长良好的亲蟹在单体框中养殖,到可交配时,采用1雄对3雌进行定向交尾。交尾成功后,记录下各雌雄亲本的编号,并在雌蟹游泳足上打上标签,放入池中越冬。2014年3月下旬,将亲蟹转入种蟹池,期间以鲜活贝类进行营养强化。
1.2.2 幼体培育及养殖将要产卵的亲蟹转入育苗池,每个育苗池1只亲蟹,产卵前,往育苗池中加入适量的扁藻及轮虫,Z1—Z2投喂轮虫及人工配合饲料,Z3—Z4投喂卤虫无节幼体,大眼幼体期以冰冻卤虫为饵料,每一期往池中加一定比例的海水,连续充气,待基本变态成为一期稚蟹后出苗,总共获得全同胞家系20个,每个家系取稚蟹3000尾,放于水泥池中养殖,利用纱网围格分隔不同家系,并保持养殖条件一致。
1.2.3 生长性状测量在60日龄、90日龄时,从每个家系中选30个个体,利用电子天平测量其体重。
1.2.4 急性低盐致死率在每个家系60日龄时,随机取出90个个体,分三组,转入盐度为4的水体中,胁迫72h,并在12、24、48、72h处记录死亡螃蟹数目,统计其死亡情况并计算其死亡率。
1.2.5 分析方法利用60日龄、90日龄的体重数据,通过SPSS线性拟合功能,进行线性拟合,以斜率为生长速率。
盐度耐受性分类采用SPSS软件K-均值聚类方法对盐度耐受能力进行分类。
对于全同胞家系,利用全同胞家组分方差分析法计算其遗传力h2,利用SPSS一般线性模型计算各组分方差:
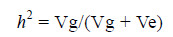
其中,Vg =(MSb-MSw)/n; Ve = MSb; Vg代表基因型方差组分; Ve代表环境方差组分; MSb(mean sums between)为组内均方; MSw(mean sums within)为组间均方。
2 结果 2.1 三疣梭子蟹60日龄和90日龄体重及增长率三疣梭子蟹60日龄和90日龄体重及其增长率见表 1,由表 1可见,各家系在60日龄时平均体重为(24.13±9.1)g,在90日龄时其平均体重为(77.50± 13.78)g。以体重为衡量标准,60日龄时体重较大的家系为1、2、14、18、20; 90日龄时体重较大的家系为3、4、6、7、18。通过线性拟合结果,发现三疣梭子蟹平均体重增长率为(0.79±0.14)g/d,体重增长率较快的家系有2、3、6、7、18。可见60日龄体重较大家系与90日龄存在明显不同,体重的增长率可以较好地说明其生长状况,故以体重增长率来衡量其生长状况,见图 1。
| 家系 | 60日龄体重均值(g) | 标准差 | 90日龄体重均值(g) | 标准差 | 增长率(g/d) | R2 |
| 1 | 39.91 | 8.92 | 77.57 | 11.81 | 0.83 | 0.97 |
| 2 | 46.04 | 12.81 | 83.63 | 25.81 | 0.91 | 0.98 |
| 3 | 24.22 | 10.58 | 91.96 | 28.69 | 0.93 | 0.81 |
| 4 | 22.50 | 5.28 | 87.07 | 27.26 | 0.88 | 0.80 |
| 5 | 14.96 | 5.51 | 69.38 | 25.53 | 0.70 | 0.76 |
| 6 | 27.86 | 7.07 | 92.16 | 26.23 | 0.94 | 0.84 |
| 7 | 27.57 | 13.61 | 100.49 | 22.98 | 1.02 | 0.81 |
| 8 | 24.12 | 7.10 | 82.27 | 26.16 | 0.84 | 0.83 |
| 9 | 13.27 | 5.91 | 87.04 | 33.76 | 0.86 | 0.71 |
| 10 | 21.27 | 8.86 | 63.27 | 25.16 | 0.65 | 0.86 |
| 11 | 12.48 | 5.58 | 58.74 | 22.76 | 0.59 | 0.76 |
| 12 | 16.27 | 23.88 | 69.41 | 18.86 | 0.70 | 0.78 |
| 13 | 10.29 | 5.62 | 43.67 | 15.03 | 0.44 | 0.78 |
| 14 | 30.90 | 11.68 | 85.43 | 20.27 | 0.89 | 0.88 |
| 15 | 15.24 | 5.81 | 57.61 | 20.08 | 0.58 | 0.81 |
| 16 | 22.90 | 10.17 | 75.14 | 22.94 | 0.77 | 0.84 |
| 17 | 25.50 | 6.37 | 79.98 | 6.87 | 0.82 | 0.85 |
| 18 | 32.96 | 6.51 | 87.66 | 17.54 | 0.91 | 0.89 |
| 19 | 25.37 | 5.35 | 77.23 | 11.20 | 0.80 | 0.86 |
| 20 | 29.12 | 6.56 | 80.20 | 9.27 | 0.83 | 0.88 |
 |
| 图 1 不同家系的体重增长率 Fig. 1 The weight-gain rate of different families |
各家系在12、24、48、72h下的死亡情况见图 2。由图可见: 部分家系在盐度4急性胁迫下48h死亡全部,大部分家系在72h处理后全部死亡,有少量家系在72h处理后,依旧有部分个体存活。计算各组死亡率方差,方差分析时发现其24h、48h胁迫下方差较大,为0.20及0.18,说明在24h、48h胁迫条件下,不同家系间的死亡率差异较大,在24h其平均致死率为50%。
 |
| 图 2 盐度4胁迫下各家系死亡率 Fig. 2 The mortality of the families under salinity 4 |
依据不同家系在盐度4不同时间下的死亡率对20个家系进行K-均值聚类分析,将其分为3类,聚类情况见表 2,各家系聚类中心见表 3并对各家系不同时间点下的聚类结果进行相关性分析,其中48h聚类结果与72h呈显著性相关(P<0.05),其它各小时组间并未发现显著相关。
| 性状 | 聚类结果 | ||
| 1 | 2 | 3 | |
| 12h | 1, 6, 12, 14, 16, 17, 19, 20 | 2, 3, 4, 9, 11, 13, 15, 18 | 5, 7, 8, 10 |
| 24h | 1, 6, 11, 12, 13, 14, 15, 16, 20 | 2, 3, 4, 9, 10, 17, 19 | 5, 7, 8 |
| 48h | 1, 6, 13, 14, 15 | 3, 4, 11, 12, 17, 18, 20 | 2, 5, 7, 8, 9, 10, 16, 19 |
| 72h | 6, 13, 14 | 1, 4, 11, 15 | 2, 3, 5, 7, 8, 9, 10, 12, 16, 17, 18, 19, 20 |
| 性状 | 聚类中心 | ||
| 1 | 2 | 3 | |
| 12h | 0.03 | 0.19 | 0.38 |
| 24h | 0.32 | 0.58 | 0.84 |
| 48h | 0.56 | 0.77 | 0.97 |
| 72h | 0.85 | 0.93 | 1.00 |
根据相关性分析,48h盐度耐受性与72h盐度耐受性分类显著相关(P<0.05),以表 2中48h低盐耐受性分类作为其耐受性分级指标,对增长率与盐度耐受性能力进行相关性分析,其相关系数为0.143,并检验无显著性相关(P=0.547)。
2.3 遗传力计算利用SPSS一般线性模型估计得全同胞组分的方差因素见表 4,并利用全同胞组分方差分析法计算,其60日龄的低盐三疣梭子蟹在低盐处理12、24、48、72h下对低盐耐受性的遗传力分别为0.13、0.18、0.21、0.29,其体重遗传力为0.45(表 5)。
| 性状 | 变异来源 | 均方 | 显著性 |
| 12h死亡率 | 全同胞组间 | 0.656 | 0.000 |
| 全同胞组内 | 0.123 | ||
| 24h死亡率 | 全同胞组间 | 1.238 | 0.000 |
| 全同胞组内 | 0.218 | ||
| 48h死亡率 | 全同胞组间 | 0.936 | 0.000 |
| 全同胞组内 | 0.133 | ||
| 72h死亡率 | 全同胞组间 | 0.601 | 0.000 |
| 全同胞组内 | 0.045 | ||
| 体重 | 全同胞组间 | 2832.415 | 0.000 |
| 全同胞组内 | 112.287 |
| 遗传力估计项目 | 12h | 24h | 48h | 72h | 生长 |
| Vg | 0.02 | 0.05 | 0.04 | 0.02 | 90.67 |
| Ve | 0.12 | 0.22 | 0.13 | 0.04 | 112.29 |
| 遗传力 | 0.13 | 0.18 | 0.21 | 0.29 | 0.45 |
遗传力反映了某一性状的差异有遗传成分所占的比例,主要有广义遗传力和狭义遗传力计算某一性状的遗传力对于选育优良性状具有重要意义。对水产生物许多物种的主要经济性状遗传力计算较多,如凡纳滨对虾其体重遗传力为0.37—0.45(Castillo- Juárez et al,2007),王庆志等(2012)计算了长牡蛎壳高的遗传力为0.161—0.387,壳长的遗传力为 0.139— 0.398。高保全等(2010)通过三疣梭子蟹37个家系评估了三疣梭子蟹的80日龄和120日龄的体重遗传为0.53和0.35,而本实验在60日龄下得到的三疣梭子蟹体重遗传力为0.45,与其较为接近,属于中高度遗传力。由此可见,三疣梭子蟹体重遗传力属于中高度遗传力,具有较好的选育前景。
3.2 三疣梭子蟹低盐耐受性遗传力分析关于水产动物对于抗逆性状的遗传力分析中,主要有抗病及一些对环境因子的抗逆性性状,中国对虾抗WSSV性状遗传力估计值0.10±0.03(杨翠华,2007)“黄海1号”耐高pH的遗传力在0.29—0.31之间,耐高氨氮的遗传力在0.15—0.22之间(黄付友,2007),Thodesen等(2013)研究奥利亚罗非鱼(Oreochromis aureus)对于冷水的耐受遗传力为0.05—0.08之间,蒋湘等(2014)利用盐度对九孔鲍(Haliotis diversicolor supertexta)的死亡率评估其低盐性状遗传力为0.056± 0.022,可见抗逆性状相对于生长性状来说遗传力较低。本文首次计算了三疣梭子蟹低盐胁迫条件不同时间下的遗传力为0.13—0.29,不同时间点估算的遗传力大小并不相同,随着低盐胁迫处理时间加长其遗传力变大,这在较为复杂的性状中较为常见,如Boomsma等(1998)通过研究多元遗传分析血压性状的遗传力,发现血压性状的遗传在不同性别间存在差异,认为血压受不同影响因子控制,各影响因子发挥的作用并不相同,进而导致其血压遗传力在不同性别中显现差异。Evans等(2006)发现斑胸草雀(zebra finch)在不同选择压力下,其后代的遗传力出现显著差异,根据表观遗传学的观点,环境压力对基因甲基化等造成影响,可以在不改变基因的情况下对后代产生影响,有些植物在不同的盐度胁迫后显现出不同的遗传力(Boyko et al,2011)。在本实验中,随着环境施压增大,可能使遗传因素造成的差异效果更加明显,导致表型上遗传方差变大,其在48h及72h胁迫下其遗传力均大于0.2。
3.3 三疣梭子蟹体重与低盐耐受性性状相关性生长及抗性性状一直是水产生物选育的重点(Gjedrem,1983),在对虾(Cock et al,2009)中研究较多,Argue等(2002)在凡纳滨对虾育种中,分别构建了以生长性状为主的家系及以生长性状加抗桃拉病毒(TSV)性状复合选育的家系进行,选育具有复合性状的优质品种。但其结果显示抗桃拉病毒性状与生长性状存在负相关。根据本实验结果,其生长性状与盐度耐受性性状相关系数为0.143,检验无显著性相关(P>0.05),虽然生长较快个体可能在体重上较大一定程度上会造成其对盐度的耐受能力上升,但根据本实验室构建的三疣梭子蟹低盐耐受性模型,6期以后三疣梭子蟹体重对低盐胁迫的耐受性已基本稳定,体重差异对其低盐耐受能力影响较小。虽然无法完全排除体重影响,且本实验结果并未出现两性状相关,可以认为在一定程度上生长与耐低盐性状无相关性,可以进行复合选育。
通过分析发现12h与24h的遗传力低于0.2,48h与72h的遗传力都高于0.2,但若以72h筛选其成活率过低,选择压力过大,由于48h耐受性与72h耐受性相关显著,且在48h的遗传力也高于0.2,故在选育过程中可采用48h处理以达到较佳选育效果。
本实验表明三疣梭子蟹生长性状属于中度遗传力,低盐耐受性性状属于中低遗传力,并且低盐耐受性状与生长性状并无相关性,本实验发现家系4、6、14具有较强的低盐耐受性及体重增长速率,可利用这些家系进行下一步的选育工作。
| 马爱军,王新安,杨志等, 2008.大菱鲆(Scophthalmus maximus)幼鱼生长性状的遗传力及其相关性分析.海洋与湖沼, 39(5):499-504 |
| 王冲,姜令绪,王仁杰等, 2010.盐度骤变和渐变对三疣梭子蟹幼蟹发育和摄食的影响.水产科学, 29(9):510-514 |
| 王庆志,李琪,刘世凯等, 2012.长牡蛎成体生长性状的遗传参数估计.中国水产科学, 19(4):700-706 |
| 王庆志,李石磊,付成东等, 2014.虾夷扇贝耐高温育种家系的建立与早期筛查.水产学报, 38(3):371-377 |
| 农业部渔业局, 2013. 2013年中国渔业统计年鉴.北京:中国农业出版社, 28 |
| 严福升, 2009.牙鲆(Paralichthys olivaceus)生长相关性状遗传力估计和耐热性状初步研究.青岛:中国海洋大学硕士学位论文, 28-45 |
| 杨翠华, 2007.中国对虾与抗性相关性状的遗传学参数分析.青岛:中国海洋大学博士学位论文, 41-80 |
| 栾生,孙慧玲,孔杰, 2006.刺参耳状幼体体长遗传力的估计.中国水产科学, 13(3):378-383 |
| 高保全,刘萍,李健等, 2010.三疣梭子蟹(Portunus trituberculatus)体重遗传力的估计.海洋与湖沼, 41(3):322-326 |
| 黄付友, 2007. "黄海1号"中国对虾生长性状和对高pH、高氨氮抗性遗传力的估计.青岛:中国海洋大学硕士学位论文, 33-43 |
| 黄永春,艾华水,潘忠诚等, 2013.凡纳滨对虾抗WSSV选育家系的建立及其抗病特性.水产学报, 37(3):359-366 |
| 梁峻,郑怀平,李莉等, 2011.虾夷扇贝养殖群体的遗传力估算.海洋科学, 35(3):1-7 |
| 蒋湘,刘建勇,赖志服, 2014.九孔鲍(Haliotis diversicolor supertexta)耐低盐与生长性状的遗传参数评估.海洋与湖沼, 45(3):542-547 |
| 戴爱云,冯钟琪,宋玉枝等, 1977.三疣梭子蟹渔业生物学的初步调查.动物学杂志,(2):30-33 |
| Argue B J, Arce S M, Lotz J M et al, 2002. Selective breeding of Pacific white shrimp(Litopenaeus vannamei) for growth and resistance to Taura Syndrome Virus. Aquaculture, 204(3-4):447-460 |
| Aulstad D, Gjedrem T, Skjervold H, 1972. Genetic and environmental sources of variation in length and weight of rainbow trout(Salmo gairdneri). Journal of the Fisheries Research Board of Canada, 29(3):237-341 |
| Boomsma D I, Snieder H, de Geus E J C et al, 1998. Heritability of blood pressure increases during mental stress. Twin Research, 1(1):15-24 |
| Boyko A, Kovalchuk I, 2011. Genome instability and epigenetic modification-heritable responses to environmental stress?. Current Opinion in Plant Biology, 14(3):260-266 |
| Castillo-Juárez H, Casares J C Q, Campos-Montes G et al, 2007. Heritability for body weight at harvest size in the Pacific white shrimp, Penaeus(Litopenaeus) vannamei, from a multi-environment experiment using univariate and multivariate animal models. Aquaculture, 273(1):42-49 |
| Cock J, Gitterle T, Salazar M et al, 2009. Breeding for disease resistance of Penaeid shrimps. Aquaculture, 286(1-2):1-11 |
| Evans M R, Roberts M L, Buchanan K L et al, 2006. Heritability of corticosterone response and changes in life history traits during selection in the zebra finch. Journal of Evolutionary Biology, 19(2):343-352 |
| Fjalestad K T, Gjedrem T, Gjerde B, 1993. Genetic improvement of disease resistance in fish:an overview. Aquaculture, 111(1-4):65-74 |
| Gjedrem T, 1983. Genetic variation in quantitative traits and selective breeding in fish and shellfish. Aquaculture, 33(1-4):51-72 |
| Langdon C J, Jacobson D P, Evans F et al, 2000. The molluscan broodstock program improving Pacific oyster broodstock through genetic selection. Journal of Shellfish Research, 19(1):616 |
| Pérez-Rostro C I, Ramirez J L, Ibarra A M, 1999. Maternal and cage effects on genetic parameter estimation for Pacific white shrimp Penaeus vannamei Boone. Aquaculture Research, 30(9):681-693 |
| Thodesen J, Rye M, Wang Y X et al, 2013. Genetic improvement of tilapias in China:Genetic parameters and selection responses in growth, pond survival and cold-water tolerance of blue tilapia(Oreochromis aureus) after four generations of multi-trait selection. Aquaculture, 396-399(1):32-42 |