 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 马林, 李新正. 2015.
- MA Lin, LI Xin-Zheng. 2015.
- 我国海洋底栖猛水蚤目两个新记录种
- TWO NEW RECORDS OF BENTHIC HAPACTICOIDA FROM CHINA SEAS
- 海洋与湖沼, 46(6): 1321-1325
- Oceanologia et Limnologia Sinica, 46(6): 1321-1325.
- http://dx.doi.org/10.11693/hyhz20141200342
-
文章历史
- 收稿日期:2014-12-08
- 改回日期:2015-01-27




猛水蚤是一类小型低等甲壳动物,形态多样,个体通常小于1mm,是底栖桡足类的主要类群,分布范围极广,广泛分布于世界各大洋中,垂直分布可从潮间带到超过5000m 的深海。猛水蚤栖息生境复杂多样,可生活在各种底质的海底,以及珊瑚礁、海蚀洞、海底热液喷口和冷渗口; 或与海藻等共栖,少数种类寄生(Huys et al,1991; Huys et al,1996; Seifried,2003; Willen,2003; Corgosinho et al,2010)。我国对猛水蚤目的分类学研究还处于初级阶段,仅在渤海湾和胶州湾有少量报道(Gee et al,2000; Mu et al,2000; Ma et al,2011; 慕芳红等,2001; Mu et al,2002; Mu et al,2004; Huys et al,2008),原因在于猛水蚤个体很小、种类繁多、数量巨大、分布很广、生物多样性程度高,鉴定非常困难,研究人员相对不足。从全国海区看,以往还没有对我国猛水蚤资源进行全面系统的分类学研究。猛水蚤是继线虫之后第二丰富的小型底栖生物类群(Huys et al,1996),搞清其种类数目和生物多样性对于底栖生态学研究和环境监测具有重要的意义。
矩头猛水蚤科Tetragonicipitidae Lang,1948 世界海域共记录17 属,中国仅记录1 属,即叶足猛水蚤属 Phyllopodopsyllus T. Scott,1906此属最明显的特征是雌性第5 胸足呈叶片状(Huys et al,1996; Wells,2007; 陈清潮,2008)。目前世界海域共记录此属62 个有效种,中国曾记录西沙群岛1 浮游种,即叉叶足猛水蚤Phyllopodopsyllus furciger Sars,1907(陈清潮,2008)。 本文记录1 种,即濑户叶足猛水蚤Phyllopodopsyllus setouchiensis Kitazima,1981,这是此种第一次在中国报道。
美猛水蚤科Ameiridae Boeck,1865 就种数而言在猛水蚤目居于第三位。此科属水平的分类系统比较混乱,很多属的建立不完善、存在疑问,且属水平的分类系统太精细,因此被称为分类学上的梦魇(Conroy-Dalton et al,1996)。此科为世界性分布,栖息环境多样,在海水和淡水中均有分布。目前此科世界上共记录45 属,中国海域仅记录1 属,即美猛水蚤属 Ameira Boeck,1865(陈清潮,2008)。Boeck(1865)建立美猛水蚤属,但仅作了简单的描述,缺乏详细的描述和绘图。此后多个作者对此属进行修订,并从此属分离出很多种,建立新属。目前,世界海域记录此属66 种,中国曾记录此属1 浮游种,即长足美猛水蚤 Ameira longipes Boeck,1865(张崇洲等,1976; 陈清潮,2008),本文记录此属1 种,即扎哈美猛水蚤Ameira zahaae Karanovic & Cho,2012,此种第一次在中国报道。
1 濑户叶足猛水蚤 Phyllopodopsyllus setouchiensisKitazima,1981(图 1)Phyllopodopsyllus setouchiensis Kitazima,1981: 413—424,figs. 15—21

|
| 图 1 濑户叶足猛水蚤 Fig. 1 Phyllopodopsyllus setouchiensisKitazima,1981 |
24♂♂,MBM189234,35°08.328′N,119°54.562′E,低潮带,砂; 118♀♀,56♂♂,MBM189235,35°08.329′N,119°54.565′E,中潮带,砂。采集人: 李新正,董超,王晓晨。采集时间: 2008 年5 月14 日。
1.2 鉴别特征 1.2.1 科的鉴别特征体细长,尾叉特别。雌性第1 触角分7—9 节,雄性异形。第2 触角内外肢各分1节。大颚具外肢和内肢。第2 小颚具4—5 个内叶。 颚足执握状。第1 胸足外肢分3 节,内肢分2—3 节。 第2—4 胸足外肢分3 节,内肢分2 节。两性第5 对胸足愈合或分开。雌性具卵囊1 个。
1.2.2 属的鉴别特征体细长。尾叉特别,一般长大于宽。额角很小。雌性第1 触角分8—9 节,第1 节很长,两性第1 触角第2 节均具有1 个向外弯曲的突起。第2 触角外肢具3 根刚毛。第1 小颚具外肢和内肢。第2 小颚具5 个内叶。颚足执握状。第1—4 胸足内肢分2 节,外肢分3 节。雌性第5 胸足叶片状,内外肢愈合; 雄性第5 对胸足基节愈合,但内肢与外肢不愈合。
1.2.3 种的鉴别特征雌性: 体呈圆柱形,体长约380—460μm(额角顶端到尾叉末端的长度),宽约 65—100μm(头胸甲最宽处); 除尾节外,身体各体节背部后缘具细齿。生殖节背面和侧面分节不明显,但背外侧明显。后体部倒数第2 体节腹面末缘具长刺,尾节腹面后缘与尾叉连接处具长刺。肛板边缘具刺,不突出。尾叉近圆柱形,长约为宽的2.5 倍,末端变形; 背面内侧边缘具微刚毛,外侧具2 根尾毛,末端具2 根尾毛,内侧1 根最长。额角很小,近方形。第 1 触角分9 节,第1 节长约为第2—4 节长度之和; 第2 节具有1 大的突起; 第4 节具感觉毛; 刚毛分布为: 1[1]+2[8]+3[8]+4[4]+5[2]+6[2]+7[2]+8[2]+9[8]。第2 触角基节与内肢第1 节等长,无刚毛; 外肢分1 节,具3 根羽状刚毛; 内肢第1 节表面有刺,第2 节侧面具1 根刚毛和2 根刺,末端具7 根刚毛。大颚基节具3 根刚毛; 外肢1 节,具5 根刚毛; 内肢1 节,具7 根刚毛。第1 小颚底节具5 根刚毛; 基节具8 根刚毛; 内肢具4 根刚毛; 外肢具3 根羽状刚毛。第2 小颚底节具4 个内叶; 基节具4 根刚毛; 内肢分3 节,刚毛分布为3: 1: 3。颚足底节具3 根刚毛; 内肢分2 节,第1 节具1 根刚毛,第2 节具1 爪和2 根刚毛。
第1—4 胸足内肢分2 节,外肢分3 节; 第1 胸足内肢长超过外肢。第1—4 胸足各节刚毛和刺的分布如下:
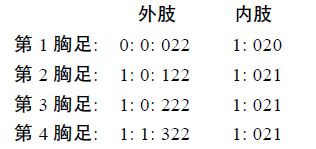
第5 胸足叶片状,形成一个卵囊状的结构,外侧具3 根刚毛,内侧具4 根刚毛。
雄性: 与雌性相似,比雌性小,体长约300— 390μm(额角顶端到尾叉末端的长度),宽约60— 80μm(头胸甲最宽处),以下特征不同。生殖节两节不愈合,第2 节及后缘相邻的体节腹面末缘具长刺。尾叉末端不变形。第1 触角近爪状,第2 节具很小的突起。第4 胸足外肢第2 节具1 个三角形的突起,末节具6 根刚毛和刺。第5 胸足内外肢不愈合,内肢具3 根刚毛; 外肢具5 根刚毛。第6 胸足具2 根刚毛。
1.3 模式标本产地日本向岛潮间带,花岗岩砂质底质,距离基准线 2.4m。
1.4 地理分布江苏平岛,日本向岛。
1.5 讨论濑户叶足猛水蚤与中国已记录的叉叶足猛水蚤的主要区别是前者第2 胸足内肢第1 节内侧具刚毛,而后者第2 胸足内肢第1 节内侧无刚毛。濑户叶足猛水蚤区别于同属其他种的主要特征是第1 触角的节数和刚毛的分布数量,胸足的刚毛分布规则,尾叉的长宽比和尾毛的分布位置。本文描述的濑户叶足猛水蚤的采集地是平岛潮间带,底质是砂。此种的模式产地是向岛,底质也为砂。本文记录的濑户叶足猛水蚤和Kitazima(1981)描述的模式标本基本一致,除1—4 胸足基节表面刺的大小和分布位置略有区别,以及雌性第5 胸足内侧具3 根裸刚毛和1 根毛状刚毛。目前,此种在中国和日本仅在潮间带记录过。
2 扎哈美猛水蚤 Ameira zahaae Karanovic& Cho,2012(图 2)Ameira zahaae Karanovic & Cho,2012: 93—101,figs. 1—4

|
| 图 2 扎哈美猛水蚤 Fig. 2 Ameira zahaae Karanovic& Cho,2012 |
29♀♀,1♂,MBM189082,35º08.328′N,119º54.562′E,低潮带,砂; 47♀♀,1♂,MBM189083,35º08.329′N,119º54.565′E,中潮带,砂。采集人: 李新正,董超,王晓晨。采集时间: 2008 年5 月14 日。
2.2 鉴别特征 2.2.1 科的鉴别特征体形粗壮或细长,额部不甚突出,头胸部与腹部无明显的分界线,生殖节由不明显的两节组成。尾叉的长度变化很大。雌性第1 触角一般为8 节,雄性第1 触角与雌性异形。第2 触角外肢分1—2 节。大颚须为单肢型。颚足一般为执握形。第1 胸足内外肢一般分3 节,内肢稍成执握形。 两性胸足一般没有明显的异形现象。第5 胸足分2 节,少数1 节。雌性卵囊1 个。
2.2.2 属的鉴别特征体细长,呈圆柱形。肛板无刺的分布。尾叉长度可变。额角很小。第1 触角分8 节,第4 节具一长感觉毛。第2 触角基节与内肢不愈合,外肢1—2 节。大颚无外肢,内肢1 节。第1 小颚内外肢仅由一些刚毛代替。第2 小颚前底节与底节愈合,最多具2 个内叶,内肢退化成几根刚毛。颚足内肢分2 节,第2 节具1 粗壮爪。第1 胸足内外肢分3 节,内肢第1 节很长,外肢第2 节内侧无刚毛。第2—4 胸足内外肢均分3 节。第5 胸足内肢与基节愈合,分 2 节。
2.2.3 种的鉴别特征雌性: 体形窄长,呈圆柱形,长约300—400μm(额角顶端到尾叉末端的长度),宽约60—70μm(头胸甲最宽处),背面具微刚毛。生殖节2 节,呈正方形,中部两侧留有节痕,卵囊1 个。 腹部除生殖节外各节腹面后缘均具刺,其中尾节中部靠近前缘也具刺。肛板光滑。尾叉略呈矩形,宽略大于长,具7 根尾毛,侧面3 根,顶端3 根,背面1 根。第1 触角分8 节,第4 节具带状感觉毛,刚毛分布为: 1[1]+2[9]+3[6]+4[4]+5[2]+6[3]+7[4]+8[7]。第2 触角外肢1 节,具刚毛3 根; 内肢第2 节内侧靠近末端具2 短刺,末端具6 根刚毛。大颚基节具1 根刚毛;无外肢; 内肢侧缘具1 根短刚毛,近末缘具1 根刚毛,顶端具3 根刚毛。第1 小颚前底节表面具2 根刚毛,顶端具5 根刺,侧缘具2 根小刺; 底节和基节各具2 根刚毛; 外肢具1 根刚毛; 内肢退化成2 根刚毛。第2 小颚前底节和底节愈合,具1 个内叶,内叶具3 根刚毛; 基节具1 粗壮爪和1 根刚毛; 无外肢; 内肢1 节,具2 根刚毛。颚足内肢第1 节外缘具长的微刚毛,第 2 节具1 粗壮的爪和1 根短的刚毛。第1—4 胸足内外肢均分3 节; 第1 胸足内肢第1 节略长于外肢; 第 2—4 胸足内肢均比外肢短。第1—4 胸足各节的刚毛和刺的分布如下:
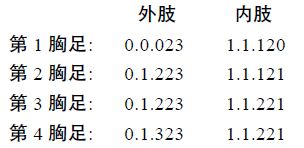
第5 胸足分2 节,内肢与基节愈合,具4 根刚毛,外侧第2 根最长,内侧2 根刚毛末端分叉。外肢具5 根刚毛,内侧2 根最长。
雄性: 长约370μm(额角顶端到尾叉末端的长度),宽约65μm(头胸甲最宽处),体形与雌性相似,除生殖节2 节不愈合,第2 节腹面末缘具长刺。第1 触角与雌性异形,第3 节和第4 节之间,第6 节和第7 节之间稍弯曲,共分8 节,第4 节具带状感觉毛。第1—4 胸足与雌性相似。第5 胸足内肢具2 根刚毛,外肢具5 根刚毛。第6 胸足具刚毛3 根。
2.3 地理分布江苏平岛,韩国西海。
2.4 讨论本文记录的中国海域发现的扎哈美猛水蚤与 Karanovic等(2012)报道的韩国海域的模式种形态基本一致,除中国海域发现的扎哈美猛水蚤第1 胸足和第4 胸足基节表面分别具3 排刺和2 排刺,而模式种第1 胸足和第4 胸足基节表面均具1 排刺。另外,扎哈美猛水蚤与Chang(2007)描述的小美猛水蚤Ameira parvula(Claus,1866)相似,两者第1 胸足形状和刚毛数,第5 胸足外肢形状及刚毛数相似,但仍有很大区别。扎哈美猛水蚤第2 触角外肢分1 节,雌性第4 胸足基节表面无刺,第5 胸足内肢内侧2 根刚毛末端分叉; Chang(2007)描述的小美猛水蚤第2 触角外肢分2 节,雌性第4 胸足基节表面分布2 排刺,第5 胸足内肢内侧2 根刚毛末端不分叉。但很多学者研究认为小美猛水蚤为世界性分布,形态具有很大的变异性,比如第2 触角外肢的节数、表面刚毛和刺的分布; 第4 胸足外肢第3 节内侧顶端有无微刚毛; 第5 胸足内肢内侧2 根刚毛顶端有无分叉,Wells等(1987)认为这些变化可能是因为一些不适当的观察造成的,而 Chang(2007)认为可能是生境和地域造成的,Karanovic等(2012)则认为现有的小美猛水蚤的报道缺乏详细细致的描述和绘图,这些变异是否是同一个种存在疑问。
扎哈美猛水蚤与微美猛水蚤Ameira parvuloides Lang,1965 非常相似,但仍有区别。扎哈美猛水蚤尾叉具7 根尾毛,大颚基节具1 根刚毛,颚足内肢第2 节具1 粗壮爪和1 根刚毛; 微美猛水蚤尾叉具5 根尾毛,大颚基节具2 根刚毛,颚足内肢第2 节具1 粗壮爪和2 根刚毛。另外两者第5 胸足外肢形状不同(扎哈美猛水蚤呈梨形,而微美猛水蚤几乎是椭圆形),刚毛之间的相对长度也不同。扎哈美猛水蚤与已报道的长足美猛水蚤的主要区别是前者第1 胸足外肢长达内肢第1 节约3/4 处; 尾节腹面具3 排刺,后者第1 胸足外肢长达内肢第1 节约2/3 处; 尾节腹面具2 排刺。
| 张崇洲, 李志英. 1976. 我国西沙群岛的猛水蚤. 动物学报,22(1): 66-70 |
| 陈清潮. 2008. 桡足亚纲. 见: 刘瑞玉主编. 中国海洋生物名录. 北京: 科学出版社, 608-634 |
| 慕芳红, 张志南, 郭玉清. 2001. 渤海底栖桡足类群落结构的研究. 海洋学报, 23(6): 120-127 |
| Boeck A. 1865. Oversigt over de ved Norges Hyster Kyster iagttagneCopepoder henhörende til Calanidernes, Cyclopidernes ogHarpacticidernes Familier. Forhandlinger i Videnskabsselskabet iKristiania. 1864: 226-282 |
| Chang C Y. 2007. Two harpacticoid species of Genera Nitokraand Ameira (Harpacticoida: Ameiridae) from brackishwaters in Korea. Integrative Biosciences, 11(2): 247-253 |
| Conroy-Dalton S, Huys R. 1996. Towards a revision of AmeiraBoeck. 1865 (Harpacticoida, Ameiridae): re-examination ofthe A. tenella-group and the establishment of Filexilia gen. n.and Glabrameira gen. n. Zoologica Scripta, 25(4): 317-339 |
| Corgosinho P H C, Martínez Arbizu P. 2010. Ameiridae Boeckand Argestidae Por revisited, with establishment ofParameiropsidae, a new family of Harpacticoida (Crustacea,Copepoda) from deep-sea sediments. Helgoland MarineResearch, 64(3): 223-255 |
| Gee J M, Mu F H. 2000. A new genus of Cletodidae (Copepoda;Harpacticoida) from the Bohai Sea, China. Journal ofNatural History, 34(6): 809-822 |
| Huys R, Boxshall G A. 1991. Copepod Evolution. London, UK:The Ray Society, 1-12 |
| Huys R, Gee J M, Moore C G et al. 1996. Synopses of the Britishfauna (New Series) No.51. In: Marine and Brackish Water Harpacticoids, Part 1. Dorchester, UK: The Dorset Press, 1,76 |
| Huys R, Mu F H. 2008. Description of a new species ofOnychostenhelia Itô (Copepoda, Harpacticoida, Miraciidae)from the Bohai Sea, China. Zootaxa. 1706: 51-68 |
| Karanovic T, Cho J L. 2012. Three new ameirid harparcticoidsfrom Korea and first record of Proameira simplex(Crustacea: Copepoda: Ameiridae). Zootaxa. 3368: 91-127 |
| Kitazima Y. 1981. Three new species of the genusPhyllopodopsyllus (Copepoda, Harpacticoida) from theInland Sea of Japan. Publications of the Seto MarineBiological Laboratory, 26(4-6): 393-424 |
| Ma L, Li X Z. 2011. Delavalia qingdaoensis sp. nov.(Harpacticoida, Miraciidae), a new copepod species fromJiaozhou Bay, Yellow Sea. Crustaceana, 84(9): 1085-1097 |
| Mu F H, Gee J M. 2000. Two new species of Bulbamphiascus(Copepoda: Harpacticoida: Diosaccidae) and a related newgenus, from the Bohai Sea, China. Cahiers de BiologieMarine, 41: 103-135 |
| Mu F H, Huys R. 2002. New species of Stenhelia (Copepoda,Harpacticoida, Diosaccidae) from the Bohai Sea (China)with notes on subgeneric division and phylogeneticrelationships. Cahiers de Biologie Marine, 43(2): 179-206 |
| Mu F H, Huys R. 2004. Canuellidae (Copepoda, Harpacticoida)from the Bohai Sea, China. Journal of Natural History, 38(1):1-36 |
| Seifried S. 2003. Phylogeny of Harpacticoida (Copepoda):Revision of “Maxillipedasphalea” and Exanechentera.Göttingen, Germany: Cuvillier Verlag, 1-2 |
| Wells J B J, Rao G C. 1987. Littoral Harpacticoida (Crustacea:Copepoda) from Andaman and Nicobar Islands. Memoirs ofthe Zoological Survey of India, 16(4): 1-385 |
| Wells J B J. 2007. An annotated checklist and keys to the speciesof Copepoda Harpacticoida (Crustacea). Zootaxa. 1568(1568):1-872 |
| Willen E. 2003. A new species of Stenhelia (Copepoda,Harpacticoida) from a hydrothermal, active, submarinevolcano in the New Ireland Fore-Arc system (Papua NewGuinea) with notes on deep sea colonization within theStenheliinae. Journal of Natural History, 37(4): 1691-1711 |