 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 焦海峰, 严巧娜, 郑丹, 尤仲杰, 严小军. 2015.
- JIAO Hai-Feng, YAN Qiao-Na, ZHENG Dan, YOU Zhong-Jie, YAN Xiao-Jun. 2015.
- 温度和盐度对埋栖性双壳类泥蚶(Tegillarca granosa) 呼吸、排泄的影响
- EFFECTS OF TEMPERATURE AND SALINITY ON OXYGEN CONSUMPTION AND AMMONIA EXCRETION OF MUDDY SHELLFISH TEGILLARCA GRANOSA
- 海洋与湖沼, 46(6): 1333-1338
- Oceanologia et Limnologia Sinica, 46(6): 1333-1338.
- http://dx.doi.org/10.11693/hyhz20141200347
-
文章历史
- 收稿日期:2014-12-12
- 改回日期:2015-04-08




2. 宁波大学海洋学院 宁波 315211;
3. 宁波市江北区海洋与渔业局 宁波 315020
2. Ocean College of Ningbo University, Ningbo 315211, China;
3. Jiangbei Ocean and Fishery Bureau, Ningbo 315020, China
呼吸、排泄是贝类新陈代谢的基本生理活动,它既反映了贝类自身的生理状态,也反映了不同环境要素对贝类基础生理活动的影响(Bayne et al,1983)。 目前,对双壳贝类呼吸与排泄的研究已有大量报道,如墨西哥湾扇贝(Argopecten irradians concentricus)(杨红生等,1998)、栉孔扇贝(Chlamys farreri)(王俊等,2002)、虾夷扇贝(Patinopecten yessoensis)(徐东等,2010)、日本扇贝(Mizuhopecten yessoensis)(Hao et al,2014)、海湾扇贝(Argopectens irradias)与太平洋牡蛎(Crassostrea gigas)(王芳等,1998)、近江牡蛎(Crassostrea hongkongensis)(王刚等,2013)、硬壳蛤(Mercenaria mercenaria)(文海翔等,2004)、西施舌(Mactra antiquata)(孟学平等,2005)、彩虹明樱蛤(Moerella iridescens)(董志国等,2006)和栉江珧(Pinna pecinata)(李金碧等,2009)等,上述研究均表明,温度是影响双壳贝类新陈代谢最重要的非生物因素,但盐度变化对双壳贝类呼吸、排泄影响的研究较少(张媛等,2007; 王刚等,2013),而温度和盐度又是决定水生生物自然分布的重要因子,它们是在贝类生理生态学研究中必须考虑的因素(Yu et al,2013)。
泥蚶(Tegillarca granosa)隶属软体动物门(Mollusca),双壳纲(Bivalvia),蚶属(Tegillarca),是泥相潮间带中重要的养殖种类。目前对泥蚶基础生理代谢的研究资料不多,焦海峰等(2013)曾对它的呼吸与排泄周年变化进行过研究,但针对不同温度和盐度条件下的呼吸与排泄的研究尚属空白。本文探讨了温度、盐度对泥蚶成贝耗氧率、排泄率的影响,旨在了解其呼吸与排泄的生理变化,丰富双壳贝类能量学的研究资料,也对进一步提高泥蚶养殖水平与保护养殖环境具有重要的意义。
1 材料与方法 1.1 实验材料泥蚶取自象山港贝类养殖区,实验前7 天采集并带回实验室。用软毛刷清除壳表面的附着物,暂养在大型水族箱中,每天上午和下午各投喂一次单胞藻,充气、换水供实验用,暂养一般超过7 天,实验前1 天停止投喂。
1.2 方法实验用海水经过滤后虹吸入水族箱中(500L),用电加热棒调节温度,每天升温1—2°C,用市售海水晶和曝气后的自来水调配盐度梯度,每天升降1—2。 当海水调制至实验条件后,稳定控温12h 以上,开始实验。温度设置6 个梯度: 13°C、18°C、23°C、28°C、 31°C、34°C,盐度设置4 个梯度: 18、23、28、33; 每个温度条件下均进行4 个盐度梯度的实验,共进行24 组实验。实验贝在各温度、盐度条件下稳定2d 以上开始实验。
停止投喂24h 后,取出泥蚶放置于锥形瓶中进行实验,将泥蚶沿瓶壁放入3L 的锥形瓶,每瓶10 只,虹吸装满过滤海水后用保鲜膜封口,以泥蚶贝壳张开为起始点进行计时,水浴控温。每组实验设置3 个重复,并设对照组(不放泥蚶)3 个。实验2h 后取锥形瓶中水样测定溶解氧(DO)、氨氮(NH4-N)的浓度。采用排水法测量泥蚶的体积。
1.3 数据处理与计算方法采用Winkler 碘量法测定溶解氧,采用次溴酸钠氧化法测定氨氮,在实验结束后用游标卡尺测定泥蚶的生物学参数,包括壳长(SL)、壳宽(SW)和壳高(SH),在60°C 下将解剖出的软体部烘干至恒重,用电子天平称量湿重(WW)和干重(DW)。
实验结束后,根据测定瓶与对照瓶内水体的DO和NH4-N 的浓度,计算泥蚶的耗氧率和排氨率
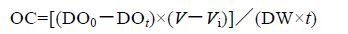
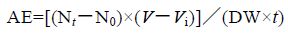
实验数据先利用Excel 预处理,方差分析和回归分析用SPSS19.0 进行,均值以Mean±SD 表示。
实验用泥蚶的生物学参数: 壳长27.14±2.15mm,壳宽20.88±1.90mm,总湿重6.96±1.78g,软体干重 0.164±0.05g。
2 结果2.1 温度、盐度对泥蚶耗氧作用的影响如图 1 所示,温度对泥蚶耗氧作用影响明显,耗氧率的高值出现在28—31°C 时,低值出现在13°C 时。在盐度为18、23、28、33 时,耗氧率均值的变化范围分别为0.66—3.15mg/(g·h)、1.02—4.53mg/(g·h)、 0.66—4.47mg/(g·h)和0.63—4.11mg/(g·h)。单因素方差分析表明,温度对耗氧率的影响极显著(F=40.909,P<0.01)。不同盐度条件下,泥蚶的耗氧率随温度变化曲线基本一致,均出现两个峰值。经回归分析,在温度13—31°C 范围内,耗氧率与温度的变化呈较显著的指数相关lnY = lna + blnT(Y=aebT)(表 1)。

|
| 图 1 温度与不同盐度条件下泥蚶耗氧作用的关系 Fig. 1 The effect of temperature on oxygen consumption of T.granosa in different salinities |
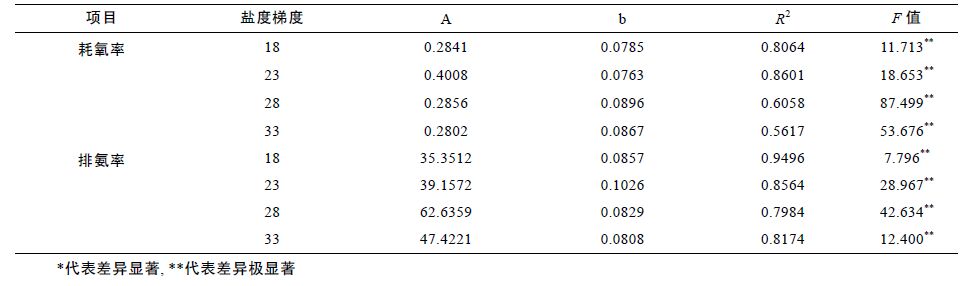 |
泥蚶耗氧率随盐度上升而上升,当盐度持续升高后,耗氧率会略有下降(图 1)。方差分析表明,温度为18°C 时,各盐度梯度下耗氧率差异极显著(F= 12.169,P<0.01),其它温度条件时,各盐度梯度下的耗氧率差异不显著(P>0.05)。
2.2 温度、盐度对泥蚶排氨作用的影响在适温范围内,排氨率随温度而上升,当温度升至一定值时,排氨率反而下降(图 2)。四个盐度梯度下,排氨率的变化范围分别为125.07—540.57、123.11—955.68、 214.55—946.97和144.30—616.91μg/(g·h)。单因素方差分析表明,不同盐度条件下,泥蚶排氨率受温度的影响均极显著(P<0.01)。经回归分析,在温度13—31°C 范围内,排氨率与温度的变化也呈较显著的指数相关(表 1)。

|
| 图 2 温度与不同盐度条件下泥蚶排氨作用的关系 Fig. 2 The effect of temperature on ammonia excretion of T.granosa in different salinity |
盐度对泥蚶排氨率的影响不明显(图 2)。单因素方差分析表明,温度为28°C 时,各盐度梯度下排氨率差异极显著(F=9.402,P<0.01),其它温度条件时,各盐度梯度下的排氨率差异不显著(P>0.05)。
2.3 温度、盐度对耗氧率与排氨率影响的交互作用在一定温度梯度(13—31°C)和盐度梯度(18—28)下,利用线性模型(Univariate)分析发现: 温度和盐度对耗氧率、排氨率的影响均存在交互作用(表 2)。 交互作用对耗氧率的影响显著(F=2.651,P<=0.025),对排氨率的影响极显著(F=3.712,P<=0.004),也就是说,温度和盐度对泥蚶基础代谢的生理活动影响明显。
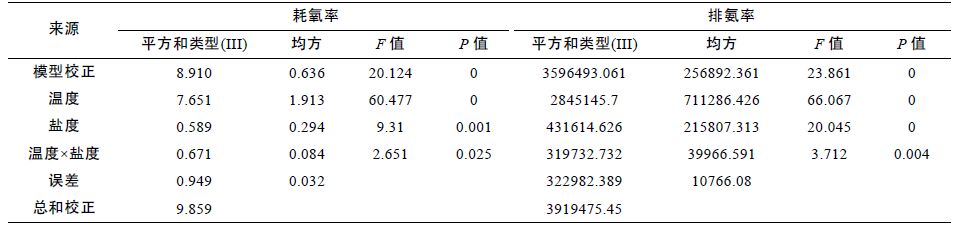 |
在上述条件下,利用线性回归(Linear Regression)分析,耗氧率(OC)、排氨率(AE)与温度(T)和盐度(S)的二元线性回归方程分别为: OC= -0.040+0.011T- 0.017S+0.002T×S(R2=0.904); AE= -25.322-0.657T- 14.333S+1.533T×S(R2=0.918)。
2.4 不同温度、盐度梯度下泥蚶呼吸排泄氧氮比(O∶N)的变化泥蚶呼吸排泄的O: N 比在4.50—13.07 之间,平均值为7.16(表 3)。方差分析表明,不同温度梯度下O: N 比有极显著差异(F=4.497,P<0.01),差异的来源是18°C 条件时O: N 比(值为10.78)。盐度对泥蚶呼吸排泄O: N 比的影响不显著(F=0.664,P>0.05)。
 |
呼吸与排泄是贝类新陈代谢基本生理活动,可以直观地反映环境条件的影响(Hammen,1968; Widdows et al,1971; Bayne et al,1983)。分析发现,泥蚶呼吸、排泄活动受温度的影响显著(P<0.01),在一定温度范围内泥蚶的耗氧率和排氨率均随温度的上升呈显著的指数相关(Y=aebT)。这与绝大多数贝类呼吸排泄的规律类似,墨西哥湾扇贝耗氧率在水温低于28°C 时随温度而升高,当温度超过30°C 时就显著下降(杨红生等,1998)。同样的拐点现象(又称为温度刺激最大响应区)在硬壳蛤中也有发现(文海翔等,2004),这种温度刺激最大响应区的现象在贻贝(Perna perna)(Mayzalld,1976)、西施舌(孟学平等,2005)、栉孔扇贝(王俊等,2002)和管角螺(罗杰等,2008)中均有发现。但研究也发现,并不是所有贝类的呼吸、排泄活动都对温度存在最大响应区,如菲律宾蛤仔(王芳等,1997)、彩虹明樱蛤(董志国等,2006)、 栉江珧(李金碧等,2009)、太平洋牡蛎(Bougrier et al,1995)和珠母贝(Saucedo et al,2004)等,它们的呼吸、 排泄率随温度而上升。值得一提的是,同样是栉孔扇贝,王俊等(2002)和王芳等(1997)研究结果也有差异: 前者表明存在温度刺激最大响应区,而后者的研究结果未发现这种最大响应区,两者差异的原因有待深入探讨。尽管不同贝类之间存在对温度刺激响应的差异,但温度是影响贝类呼吸、排泄的重要因素获得了普遍认可。本研究表明,在泥蚶的生理代谢过程中存在温度最大响应区,当温度在31°C 时,泥蚶耗氧率和排氨率显著下降。焦海峰等(2013)结果表明,在水温较低的1—7 月份,耗氧率、排氨率随温度而升高,耗氧率、排氨率的下行拐点出现在9 月份,此时水温约27°C。这与本次实验中呼吸代谢活动存在温度最大响应区的结果相互印证。但在相似的温度条件下,泥蚶耗氧率和排氨率存在差异,作者认为这种差异是由于不同实验条件造成的。Bougrier等(1995)也认为贝类耗氧率变化的差异跟环境条件密切相关,在自然条件下和在实验条件下对样品进行驯化,耗氧率会不同。
盐度也是影响贝类生理代谢的关键因子之一,它影响机体渗透压的调节。资料显示: 管角螺的排氨率存在盐度刺激最大响应区,在盐度12—27 之间,排氨率随盐度而升高,在盐度27 时升至峰值,当盐度32 时排氨率开始下降(罗杰等,2008)。青蛤(刘英杰,2005)和翡翠贻贝(杨晓新,2000)的耗氧率和排氨率随盐度的升高而升高。近江牡蛎耗氧率和排氨率与盐度呈二阶方程相关,在中盐度条件(15—20)时,耗氧率和排氨率最低; 在低盐和高盐条件下,两者均较高(王刚等,2013)。Stickle等(1982)和Tang等(2005)的研究结果与之相似。本研究中泥蚶耗氧率和排氨率虽然在中盐度时略有升高,但经方差分析,差异并不显著(P>0.05)。当不考虑极高盐条件(33)时,泥蚶的耗氧率和排氨率均随盐度而上升,这与沈伟良等(2008)对毛蚶稚贝的研究结果类似,反映出蚶科贝类具有相似的生理学特性。
温度对贝类耗氧率、排氨率的影响较为一致,但盐度的影响比较复杂,这可能与动物的种类和生长适宜盐度有关,也与贝类自身的生理特性相关。 Zurburg等(1981)研究发现某些贝类通过游离氨基酸(FAA)库中氨基酸的合成与排出来调节机体的渗透压,在合成和排出氨基酸的过程中消耗能量,造成体内耗氧和排氨升高。王刚等(2013)在近江牡蛎中也发现同样的现象,但这种现象并未在泥蚶中发现。另外,温度、盐度对泥蚶耗氧率和排氨率的影响存在交互作用,以二元线性回归方程为佳: OC(AE)=a+bT+ cS+dT×S。王芳等(1998)对海湾扇贝和太平洋牡蛎的呼吸和排泄研究也表明湿重与温度的交互作用对其耗氧率有影响。因此,环境因素对贝类呼吸代谢的影响机制较为复杂,贝类的耗氧率和排氨率不仅与生物体大小有关,更与其生理状态和所处环境相关,要探讨贝类呼吸代谢的生理学特性需要综合考虑多个因子,这也对实验生态学的理论研究提出新的要求。
3.2 氧氮比变化氧氮比(O∶N)是表征动物呼吸底物的重要参数,反映了动物在特定状态下体内的脂肪、糖类和蛋白质代谢的比例关系(Person et al,2004)。Mayzalld(1976)指出,若O∶N 比低于7 完全由蛋白质的消耗提供能量; Ikeda等(1974)认为O: N 比大于24 时是由蛋白质和脂肪联合供能,若比值无限大则由碳水化合物或脂肪供能(Conover et al,1968)。本研究中,泥蚶的 O∶N 比均值为7.16,推测机体主要依赖消耗蛋白质维持基础代谢。在泥蚶耗氧率、排氨率的周年变化中发现,O∶N 比在冬季明显较高,其中11 月份和1 月份均超过20(焦海峰等,2013)。本文的O∶N比与5—7 月份较为相近,这可能与本实验在春季开展有关。墨西哥湾扇贝的O∶N比在不同的调节实验条件下差异也不显著(杨红生等,1998),但近江牡蛎呼吸排泄的 O∶N 比随盐度变化差异显著(王刚等,2013),西施舌处于适温状态的O∶N 比高于低温和高温状态(孟学平等,2005),环境因素变化对贝类呼吸排泄O∶N 比的差异原因,需要深入探讨。
目前,O∶N 比高低与环境对有机体的压力密切相关得到普遍认可,可以作为生物体适应环境压力的指标之一(Widdows et al,1971; Mukadam et al,2014)。Montagna等(2008)也认为O∶N 比可以作为评估生理学调节的工具,因为它对毒性反应的免疫应答具有敏感性。同时Pillai等(2002)研究发现沙虾(Meta)在运输过程中和适应到新环境后O∶N 比发生了显著的改变,利用O∶N 比反映环境改变对机体的影响在泥蚶中也得到了体现(焦海峰等,2013)。但本实验环境的变化对泥蚶O∶N比的影响没有明显的规律,因此短期胁迫对泥蚶生理活动的影响程度尚待研究,而且环境胁迫对贝类的影响程度以及对贝类生理活动规律的影响机制也不清晰。
4 结论本文研究了温度、盐度对泥蚶呼吸、排泄的影响。 结果表明,温度对泥蚶耗氧率、排氨率的影响均极显著,耗氧率和排氨率与温度的变化呈显著的指数相关; 盐度对泥蚶耗氧率、排氨率的影响不明显; 而温度与盐度对耗氧率、排氨率的影响均存在交互作用,且以二次线性方程拟合最优。
| 王刚, 张其中, 喻子牛等. 2013. 盐度对近江牡蛎耗氧率、排氨率、O: N 和吸收率的影响. 海洋科学, 37(2): 101-106 |
| 王芳, 董双林, 李德尚. 1997. 菲律宾蛤仔和栉孔扇贝的呼吸与排泄研究. 水产学报, 21(3): 252-257 |
| 王芳, 董双林, 张硕等. 1998. 海湾扇贝(Argopectenirradians)和太平洋牡蛎(Crasostrea gigas)呼吸和排泄的研究. 青岛海洋大学学报, 28(2): 233-239 |
| 王俊, 姜祖辉, 唐启升等. 2002. 栉孔扇贝耗氧率和排氨率的研究. 应用生态学报, 13(9): 1157-1160 |
| 文海翔, 张涛, 杨红生等. 2004. 温度对硬壳蛤Mercenariamercenaria (Linnaeus. 1758)呼吸排泄的影响. 海洋与湖沼,35(6): 549-554 |
| 刘英杰. 2005. 青蛤摄食生理和代谢生理以及能量收支的基础研究. 青岛: 中国海洋大学硕士学位论文, 32-45 |
| 李金碧, 龚世园, 喻达辉. 2009. 温度和盐度对栉江珧耗氧率和排氨率的影响. 安徽农业科学, 37(5): 2016-2018 |
| 杨红生, 张涛, 王萍等. 1998. 温度对墨西哥湾扇贝耗氧率及排泄率的影响. 海洋学报, 20(4): 91-96 |
| 杨晓新, 林小涛, 计新丽等. 2000. 温度、盐度和光照条件对翡翠贻贝滤水率的影响. 海洋科学, 24(6): 36-39 |
| 沈伟良, 尤仲杰, 施祥元. 2008. 不同规格及不同盐度下毛蚶稚贝耗氧率和排氨率的研究. 海洋水产研究, 29(2):53-56 |
| 张媛, 方建光, 毛玉泽等. 2007. 温度和盐度对橄榄蚶耗氧率和排氨率的影响. 中国水产科学, 14(4): 690-694 |
| 罗杰, 刘楚吾, 李锋等. 2008. 盐度及规格对管角螺耗氧率和排氨率的影响. 海洋科学, 32(5): 46-50 |
| 孟学平, 董志国, 程汉良等. 2005. 西施舌的耗氧率与排氨率研究. 应用生态学报, 16(12): 2435-2438 |
| 徐东, 张继红, 王文琪等. 2010. 温度变化对虾夷扇贝耗氧率和排氨率的影响. 中国水产科学, 17(5): 1101-1106 |
| 董志国, 李晓英, 孟学平等. 2006. 彩虹明樱蛤的耗氧率与排氨率研究. 台湾海峡, 25(4): 503- 508 |
| 焦海峰, 项翔, 尤仲杰等. 2013. 泥蚶、缢蛏和僧帽牡蛎呼吸与排泄的周年变化. 海洋学报, 35(6): 147- 153 |
| Bayne B L, Newell R C, Saleuddin A S M et al. 1983.Physiological Energetics of Marine Molluscs. New York:Academic Press, 407-515 |
| Bougrier S, Geairon P, Dealous-Paoli J M et al. 1995. Allometricrelationships and effects of temperature on clearance andoxygen consumption rates of Crassostrea gigas (Thunberg).Aquaculture, 134(1-2): 143-154 |
| Conover R J, Corner E D S. 1968. Respiration and nitrogenexcretion by some marine zooplankton in relation to theirlife cycles. Journal of Marine Biological Association of theUnited Kingdom, 48(1): 49-75 |
| Hammen C S. 1968. Aminotransferase activities and amino acidexcretion of bivalve mollusks and brachiopods. ComparativeBiochemistry and Physiology, 26(2): 697-705 |
| Hao Z L, Tang X J, Ding J et al. 2014. Effect of high temperatureon survival, oxygen consumption, behavior, ammonia-N excretion, and related immune indicators of the Japanesescallop Mizuhopecten yessoensis. Aquaculture International,22(6): 1863-1876 |
| Ikeda T. 1974. Nutrition ecology of marine zooplankton.Memoirs of the Faculty of Fisheries Hokkaido University,22(1): 1-97 |
| Mayzalld P. 1976. Respiration and nitrogen excretion ofzooplankton. Ⅳ . The influence of starvation on themetabolism and the biochemical composition of somespecies. Marine Biology, 37(1): 47-58 |
| Montagna M C, Collins P A. 2008. Oxygen consumption andammonia excretion of the freshwater crab Trichodactylusborellianus exposed to chlorpyrifos and endosulfaninsecticides. Pesticide Biochemistry and Physiology, 92(3):150-155 |
| Mukadam M, Kulkarni A. 2014. Acute toxicity of cypermethrin,a synthetic pyrethroid to estuarine clam Katelysia opima(Gmelin) and its effect on oxygen consumption. Journal ofAgricultural Chemistry and Environment, 3(4): 139-143 |
| Person-Le Ruyet J, Mahé K, Le Bayon N et al. 2004. Effects oftemperature on growth and metabolism in a Mediterraneanpopulation of European sea bass, Dicentrarchus labrax.Aquaculture, 237(1-4): 269-280 |
| Pillai B R, Diwan A D. 2002. Effects of acute salinity stress onoxygen consumption and ammonia excretion rates of themarine shrimp Metapenaeus Monoceros. Journal ofCrustacean Biology, 22(1): 45-52 |
| Saucedo P E, Ocampo L, Monteforte M et al. 2004. Effect oftemperature on oxygen consumption and ammonia excretionin the Calafia mother-of-pearl oyster, Pinctada mazatlanica(Hanley. 1856). Aquaculture, 229(1-4): 377-387 |
| Stickle W B, Bayne B L. 1982. Effects of temperature andsalinity on oxygen consumption and nitrogen excretion inThais (Nucella) lapillus (L.). Journal of ExperimentalMarine Biology and Ecology, 58(1): 1-17 |
| Tang B J, Liu B Z, Yang H S et al. 2005. Oxygen consumptionand ammonia-N excretion of Meretrix meretrix in differenttemperature and salinity. Chinese Journal of Oceanology andLimnology, 23(4): 469-474 |
| Widdows J, Bayne B L. 1971. Temperature acclimation ofMytilus edulis with reference to its energy budget. Journal ofthe Marine Biological Association of the United Kingdom,51(4): 827-843 |
| Yu Z H, Qi Z H, Hu C Q et al. 2013. Effects of salinity oningestion, oxygen consumption and ammonium excretionrates of the sea cucumber Holothuria leucospilota.Aquaculture Research, 44(11): 1760-1767 |
| Zurburg W, De Zwaan A. 1981. The role of amino acids inanaerobiosis and osmoregulation in bivalves. Journal ofExperimental Zoology, 215(3): 315-325 |