 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 徐安乐, 蒋鹏达, 黎中宝, 黄永春, 陈强, 李文静. 2015.
- XU An-Le, JIANG Peng-Da, LI Zhong-Bao, HUANG Yong-Chun, CHEN Qiang, LI Wen-Jing. 2015.
- 超微粉碎浒苔(Enteromorpha prolifera)对花鲈生长、非特异性免疫及消化酶活性的影响
- EFFECTS OF ENTEROMORPHA PROLIFERA ULTRAFINE POWDERS ON GROWTH PERFORMANCE, NON-SPECIFIC IMMUNITY AND DIGESTIVE ENZYME ACTIVITY OF LATEOLABRAX JAPONICUS
- 海洋与湖沼, 46(6): 1494-1501
- Oceanologia et Limnologia Sinica, 46(6): 1494-1501.
- http://dx.doi.org/10.11693/hyhz20151000268




2. 福建省海洋渔业资源与生态环境重点实验室 厦门 361021
2. Fujian Provincial Key Laboratory of Marine Fishery Resources and Eco-environment, Xiamen 361021, China
近年来水产养殖业集约化程度不断提高,随之而来的病害与渔药残留问题也日益突出,不断恶化的生态问题严重阻碍了水产养殖业的健康可持续发展,积极开发出更高效的绿色环保型、抗病促长添加剂,调控水产动物营养和免疫,成为解决这一问题的重要途径。
浒苔(Enteromorpha prolifera)俗称苔条、青海苔等,隶属于绿藻门(Chlorophyta)、绿藻纲(Chlorophyceae)、石莼目(Ulvales)、石莼科(Ulvaceae)、浒苔属(Enteromorpha)(曾呈奎等,1962)。浒苔属广温、广盐性的藻类,其抗逆性强,广泛分布于世界海洋、河口、海陆结合部的咸淡水生境中,是一种资源丰富、容易获得的高蛋白、低脂肪的优质海藻(杨欢等,2013)。浒苔不仅含有丰富的营养物质,而且还含有多糖、萜类和甾体等多种生物活性物质,具有一定的药用价值。国内外有关浒苔在养殖业中的应用已经有诸多研究报道,浒苔作为饲料添加剂添加于畜禽和水产动物饲料中具有明显的促进动物生长、增强免疫力、改善肉质的作用,在畜禽以及水产动物,如梭鱼(Liza haematocheila)(闫杰,2012)、大黄鱼(Pseudosciaena crocea)(Asino,2010)、黄斑蓝子鱼(Siganus oramin)(周胜强等,2013)等水生动物上均有研究。
花鲈(Lateolabrax japonicus)属鱼纲(Pisces)、鲈形目(Perciformes)、鮨科(Servanidae)、花鲈属(Lateolabrax),主要分布于西北太平洋沿岸的日本、朝鲜和我国沿岸海域,属广温、广盐性浅海近岸中下层鱼类(冯昭信,1998),是我国沿海和江河的重要经济鱼类之一。目前国内外有关花鲈的研究,主要围绕免疫调节剂(Yu et al,2014)、营养调控机制(Thilagam et al,2014)、饲料原料开发(Zhang et al,2014)等方面,而浒苔在花鲈饲养中的应用还未发现有相关报道。由于浒苔具有坚韧的纤维素细胞壁,细胞内的营养物质被动物体充分吸收利用过程中存在一定障碍,本研究试将浒苔经过超微粉碎处理后添加到花鲈的饲料中,探究其对花鲈生长性能、非特异性免疫及消化酶活性的影响,为浒苔成为新型抗病促长环保型水产饲料添加剂提供理论依据。
超微粉碎是指将物料颗粒粉碎至10—25 μm,是粉体工程中的一项技术,固形物质经过超微粉碎后,颗粒的表面积和孔隙率会增加,从而获得一些独特的物理化学性质(刘树立等,2007)。将天然植物进行超微粉碎可以增强其有效成分在动物体内的吸收,因其细胞破壁率提高,而且微分粒径减小,更容易吸附于小肠壁上被吸收。超微粉碎技术在众多领域,如化妆品、食品、化工、医药特别是中草药加工等领域中均有应用(李铁梁等,2013)。
1 材料与方法 1.1 实验材料2014 年3—4 月从厦门周边各个滩涂池采集浒苔原料,用现场海水洗净,装入样品袋中,带回实验室后继续用自来水冲洗干净。沥干,晾晒,放入60°C 烘箱烘干,利用冷冻粉碎干燥机(山东三清不锈钢设备有限公司制造)将浒苔制成浒苔粉,过200 目筛网(即粉末粒径≤0.074 mm),保存备用。
花鲈苗购自福建省漳浦县锦兴育苗场,经暂养驯化后,于纯淡水中养殖。暂养期间投喂基础饲料,每天定时投喂2 次,持续14 d 后,选择体格健壮、无病无伤、规格一致的花鲈,随机分配到实验缸展开实验。共6 组,每组3 个平行共18 个水族缸,每缸30 尾,实验鱼平均体重为(5.16±0.25)g,体长为(6.63±0.56)cm。
1.2 饲料处理及实验设计将超微粉碎浒苔按照0%、1%、2%、3%、4%、5%的配比加入常规水产饲料原料(购于厦门市嘉康饲料有限公司,原料成分及基本营养成分见表 1)中,混合均匀,经制粒机(华南理工大学科技实业总厂制造)制成粒径为2.5mm 的颗粒型饲料,分别标记为对照组、Diet 1、Diet 2、Diet 3、Diet 4、Diet 5 共6 组。饲料投喂量按鱼体质量的3%左右投喂,并根据摄食情况调整,每天定时(8: 00—8: 30,17: 00—17: 30)投喂2 次,用虹吸管清理水族缸底部污物并换水,每次换水量为30%—50%,24 h 充气增氧,实验期间水温25—27.8°C,溶氧>6.0 mg/L,pH 为7.5—8.1,氨氮浓度<0.2 mg/L。实验周期28 d,于2014 年5—6 月在集美大学海水养殖实验场进行。
饲喂满28 d 后对实验鱼饥饿24 h,麻醉后对每缸鱼称重并计数,从每缸随机捞取9 尾,尾静脉处采血,将收集到的血样转移至实验室,4°C 静置过夜(约18 h),4°C、3500 r/min 条件下离心10min,收集上清液,做好标记,即为待测血清,置于–80°C 冰箱保存待测。每缸取4 尾鱼进行解剖,取肠、肝脏组织,称取肝脏重,并将肠和肝脏组织立即投入–196°C 液氮中,转移至实验室放置于–80°C 冰箱中保存待测。每缸另取 5 尾全鱼保存于–20°C 冰箱,用于鱼体成分分析。
1.4 样品分析检测方法 1.4.1 生长指标分析
特定生长率(SGR,%/d)=(lnWt-lnWo)/t×100;
饲料系数(FCR,%)= F/(Wt-Wo);
存活率(SR,%)= Nt/No×100;
肝指数(HIS,%)= WL/WB×100;
肥满度(CF,%)= Wt/L3×100;
实验鲈鱼全体成分测定方法: 粗蛋白含量测定采用凯氏定氮法(总N×6.25); 索氏抽提法(参照GB/T6433-2006)测粗脂肪; 采用马福炉灼烧法(550°C,参照GB/T6438-2007)测定粗灰分; 测水分是在75°C烘箱中将鱼体烘至恒重,计算风干状态下鱼体失去水分的比率。
1.4.2 消化酶指标测定测定消化酶活力时,根据不同消化酶的要求,用温度4°C,浓度为0.7%的生理盐水与待测样品按比例混合,用组织匀浆器在冰水浴条件下约12000 r/min 匀浆,分别制成相应浓度的匀浆液,之后用低温离心机2500 r/min,离心10 min,取上清液待测,AMS、LPS、蛋白酶及组织TP 含量均由南京建成生物工程研究所提供相应试剂盒测定。
1.4.3 血清非特异性免疫指标测定血清中T-SOD活力测定是根据黄嘌呤氧化酶法,酶单位定义是每毫升反应液中SOD 抑制率达50%时所对应的SOD 量为一个活力单位(U); LSZ 活力采用比浊法进行检测,单位为μg/mL; AKP 单位定义为100 mL 血清在37°C与基质作用15 min 产生1 mg 酚为1 个活力单位。T-SOD、LAM、AKP 及血清蛋白均由南京建成生物工程研究所提供相应试剂盒测定。
1.5 数据统计与分析实验数据均采用SPSS19.0 数据软件进行统计学分析,用单因素方差分析,Duncan 比较,数据全部用平均值±标准误(Mean±SE)表示,取显著水平P<0.05。
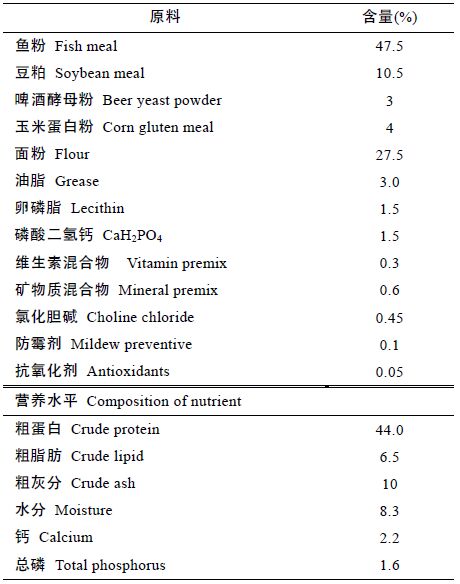
2 结果2.1 超微粉碎浒苔对花鲈鱼体基本营养成分的影响由图 1 可知,添加了超微粉碎浒苔之后,花鲈鱼体的水分有所降低,在Diet 3(3%)明显下降(P<0.05),同时,粗蛋白含量得到提升,在Diet 4(4%)含量最高,较对照组差异显著(P<0.05),而粗脂肪和灰分则没有明显变化(P<0.05)。

|
|
图 1 花鲈鱼体基本营养成分(风干样)
Fig. 1 The basic nutrient composition of L. japonicus(dry sample) a、b、c: 对于花鲈水分含量指标,上标字母不同表示不同组之间差异显著(P<0.05); 1、2、3: 对于花鲈粗蛋白指标,上标字母不同表示不同组之间差异显著(P<0.05)。A: 对于花鲈粗脂肪指标,上标字母相同表示不同组之间差异不显著(P>0.05); B: 对于花鲈灰分指标,上标字母相同表示不同组之间差异不显著(P>0.05) |
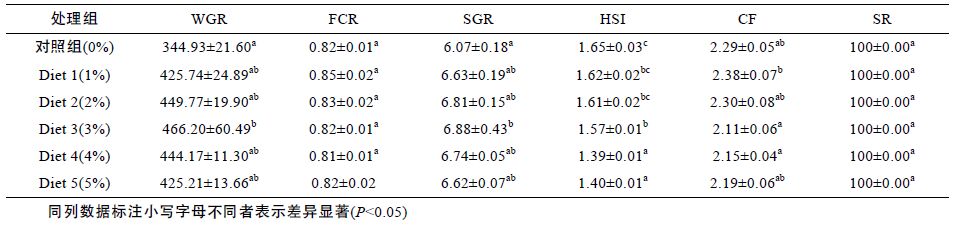
由表 2 可知,花鲈的WGR、SGR 随着浒苔添加量的增加有明显提高,其中在Diet 3 时,与对照组相比,WGR、SGR 均有明显提高,差异显著(P<0.05),但FCR 基本稳定(P>0.05),随着浒苔添加量的提高,HSI下降趋势明显,其数值在Diet 4 最低,较对照组差异显著(P<0.05)。而CF 在Diet 1 时较大,随着添加量的增多肥满度有变小趋势,但与对照组相比差异不显著(P>0.05)。
 |
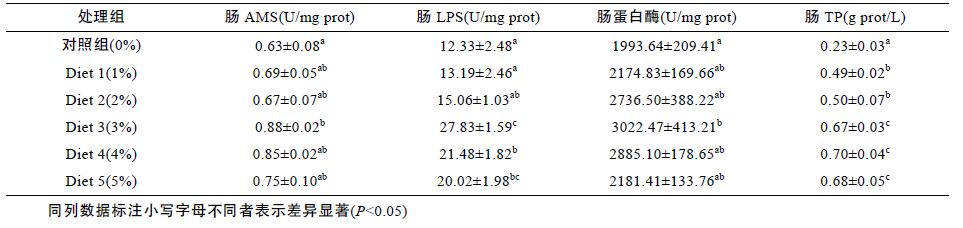
由表 3、图 2 可知,随着饲料中浒苔含量的增加,肠AMS、肠LPS、肠蛋白酶和胃LPS 的变化趋势基本一致,均在浒苔添加量为3%时达到最大,且与对照组相比有显著差异(P<0.05),随着浒苔含量的继续增加,以上各酶活力呈现减小趋势。而胃AMS和胃蛋白酶却在对照组活力最大,随着浒苔添加量的增加呈现不断降低的趋势。其中胃AMS 的降低趋势最明显,对照组的酶活力明显高于其余各组(P<0.05),且其余各组间也有明显差异。胃蛋白酶活力也在对照组最大,Diet 1 较对照组略有降低,其余四组较对照组均有显著降低(P<0.05),且其余四组之间也有明显差异。肠TP 含量、LPS 含量和胃TP 含量、LPS 含量变化趋势基本一致,均在高添加量时较对照组有显著提高(P<0.05)。
 |

|
|
图 2 超微粉碎浒苔对花鲈胃消化酶的影响
Fig. 2 Effects of the E. prolifera ultrafine powders on activities of stomach digestive enzyme of L. japonicus a—d: 表示花鲈胃AMS 指标,上标字母不同表示不同组之间差异显著(P<0.05); A—D: 表示花鲈胃蛋白酶指标,上标字母不同表示不同组之间差异显著(P<0.05); 1—5: 表示花鲈胃LPS 指标,上标字母不同表示不同组之间差异显著(P<0.05); m—n: 表示花鲈胃TP 指标,上标字母不同表示不同组之间差异显著(P<0.05) |
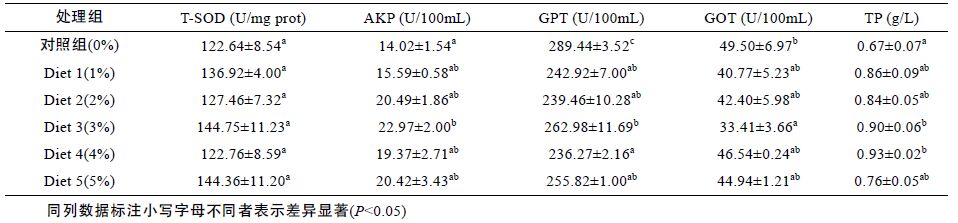
由表 4 可知浒苔添加剂对花鲈肝脏中的几种非特异性免疫酶有比较明显的影响,其中AKP 最小值为对照组,最大值为Diet 3,且差异显著(P<0.05),较对照组提高了63.8%; 而GPT 跟GOT 的变化趋势一致,对照组活力最大,分别在Diet 4和Diet 3 呈现最小,TP 最小值为对照组,随着添加量的增多其活力逐渐增大,直至Diet 3和Diet 4 达到最大,显著高于对照组(P<0.05),但在Diet 5 又有所下降; 而T-SOD 活力在各组之间没有明显变化(P>0.05)。
 |
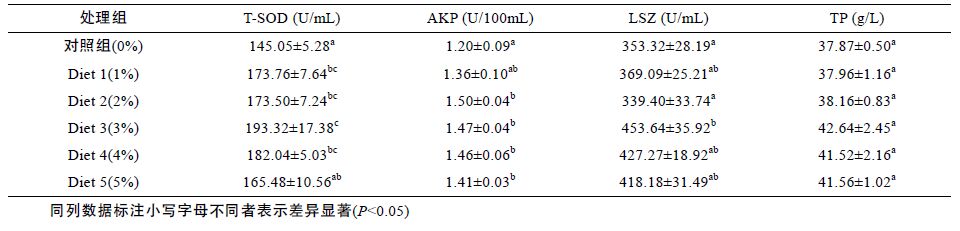
由表 5 可知浒苔添加剂对花鲈血清中几种非特异性免疫酶的影响,随着饲料中浒苔含量的增加,T-SOD 活力呈现先增大后减小的趋势,Diet 1 较对照组有显著的增大(P<0.05),且Diet 1 与Diet 2 基本一致,分别较对照组提高了17.8%和19.6%,其活力在Diet 3 达到最大(P<0.05),比对照组提高了33.3%,随着浒苔添加量的继续增多,酶活力逐渐减小; AKP 活力呈现先增大,后保持稳定的趋势,在Diet 2 及以上添加组较对照组有显著提高(P<0.05),最大值为Diet2,比对照组提高了25%; LSZ 在diet 3 较对照组有显著提高(P<0.05),比对照组提高了28.4%,而血清TP并无显著变化(P>0.05)。
 |
浒苔富含维生素和矿物质,具有天然来源、营养丰富、高蛋白、低脂肪、无毒害等特点(刘智禹,2012),已经较多地应用到了畜禽和水生动物的养殖中,本研究显示,添加了超微粉碎浒苔的实验组,花鲈鱼体的水分有所降低,而粗蛋白含量得到提升,且在Diet 4(4%)含量最高,有研究表明,鱼体肌肉水分含量与剪切力成正相关(Hagen et al,2007),而随着剪切力增加,肌肉嫩度则相应的减小,硬度提高,说明浒苔具有改善花鲈鱼肉质构特性的作用。这与Asino(2010)在大黄鱼中的研究结果有异,可能由于养殖环境和试验时间不同所致。周胜强等(2013)对黄斑蓝子鱼的研究显示,浒苔对鱼体各项体成分含量均没有显著差异(P>0.05),5%以上含量的浒苔添加组,实验鱼的生长指标有明显降低(P<0.05),但加入非淀粉多糖酶后,10%和15%浒苔组生长性能才恢复至对照组水平。说明浒苔的提前处理对提高其利用率有很大作用,而超微粉碎工艺正是为了能够让浒苔更充分的被鱼体消化,避免因利用率低下造成浪费,本实验中,蛋白和水分含量均有变化,粗脂肪和灰分没有明显变化,可能由于浒苔添加量以及花鲈与黄斑蓝子鱼食性不同等诸多差异造成的。本实验花鲈的粗蛋白在Diet 4 明显升高,水分含量在Diet3(3%)最低,说明了适量的浒苔能够提高鱼体蛋白含量,降低水分含量,而浒苔对鱼体脂肪含量及灰分并无明显影响。
3.2 超微粉碎浒苔对花鲈生长性能的影响浒苔富含价值较高的营养物质,如氨基酸、海藻多糖、蛋白质、维生素和大量微量元素等(林建云等,2011),王晓兰等(2014)将复方中草药提取液、L-甘氨酸和干浒苔粉分别加入大菱鲆(Scophthalmus maximus L.)配合饲料中进行试验,结果发现浒苔组WGR 最高,说明了浒苔能够促进大菱鲆生长,柠檬酸处理后的浒苔能够显著提高梭鱼增重率(解康等,2012),Yousif等(2004)研究了浒苔对黄斑蓝子鱼的效果,认为饲料中不应添加多于10%的浒苔。黄斑蓝子鱼属于植食性鱼类,而且在自然环境中最喜欢摄食的海藻就是浒苔(曾凡归,2010),它应该较肉食性鱼类如鲈鱼、大黄鱼等更适应饲料中的浒苔,但上述实验结果却并非如此,因为Asino(2010)对大黄鱼的研究表明饲料中浒苔含量至少为15%对其生长无明显影响,分析其原因可能是因为大黄鱼跟黄斑蓝子鱼饲料配方的差异以及养殖环境的不同所致,大黄鱼养殖在网箱中,而黄斑蓝子鱼养殖在水族缸中,相对而言,网箱养殖损失的饲料会多一点,所以饲料中浒苔含量较多也属正常,说明实际生产中在向饲料中添加浒苔时要考虑到养殖对象的习性和摄食偏好。
本实验通过超微粉碎浒苔,使浒苔藻粉的粒径更小,更容易被花鲈消化吸收,结果显示,超微粉碎浒苔对花鲈的WGR和SGR 均有显著影响,添加量为3%时达到最大,且比对照组有显著提高(P<0.05),这与Asino(2010)的研究结果一致,但随着浒苔含量的继续增多,本实验中WGR和SGR 却有下降的趋势,表明浒苔在饲料中要适量添加,过量的浒苔对花鲈的生长可能有害,这与Yıldırım等(2009)对虹鳟(Oncorhynchus mykiss)的研究结果一致。赵瑞祯(2012)对鲈鱼的研究同样表明饲料中添加过多海藻会抑制鱼体生长,分析原因可能因为海藻有特殊的鲜味,添加量过多超出了花鲈的喜好范围,降低了饲料适口性所致,另外由于浒苔经过了超微粉碎工艺的加工,大大提高了鱼体的吸收效率,所以其适宜添加量(3%)低于石莼的添加量(15%)是正常的。本实验结果显示SR和FCR均无明显变化(P>0.05),在鱼体病理状态下,HSI 可以用来反应肝损伤的程度(杨毅雯,2013),而在正常情况下,HSI 则可以在一定程度上判断鱼体是否有脂肪肝等症状,由于现代养殖为了追求高产,饲料中往往会添加过量的营养,但过量的营养往往会促进鱼体肝脏脂肪的形成(Du et al,2006),导致HSI 增大,严重的还会形成营养性脂肪肝,本实验结果显示HSI 在Diet 4(4%)显著低于对照组(P<0.05),说明了添加浒苔能够促进肝脏代谢,降低脂肪肝发生的几率。
3.3 超微粉碎浒苔对花鲈消化酶活力的影响浒苔中含有的海藻多糖能够影响酶和蛋白质表达进而促进机体生长代谢。研究表明(郝凤奇等,2008)海藻提取物能够显著提高蛋白质消化率,原因可能是一方面由于海藻中含有的海藻酸通过与饲料中的含氮物质反应,使蛋白质分解为小肽,利于吸收; 另一方面通过调节动物肠道酸碱平衡,起到抑制病原菌的作用,促进消化道中消化酶的分泌。适量的浒苔可以显著增强肉鸡的消化酶活性(孙建凤等,2010; 王述柏等,2013a),与本实验结果一致,本实验中添加浒苔之后花鲈的肠AMS、肠LPS、肠蛋白酶和胃LPS的变化趋势均为先升高后降低,均在Diet 3(3%)达到最大,且与对照组相比有显著差异(P<0.05),说明浒苔能够明显提高花鲈肠道消化酶活性,而实验的生长指标也说明了Diet 3 的花鲈生长最好,分析原因为浒苔中含有丰富的多糖及其复合物,这些物质能够有效地促进花鲈机体代谢,促进消化酶活性的提高,但消化酶的活性具有负反馈调节机制,过量的浒苔对其起到一定的抑制作用。而实验中胃蛋白酶以及胃AMS 却呈现出对照组活性最高,并逐渐降低的趋势,这可能由于浒苔中的一些活性物质在胃和肠道处具有不同的吸收机制,导致胃肠消化酶活性并不同步,也可能由于蛋白酶和LPS 在花鲈的胃和肠道本身是就具有不同甚至相反的活性变化规律,这有待继续研究。对蛋鸡的研究表明浒苔可以提高肠道有益菌的数量(王述柏等,2013b),而鱼肠道菌群情况与普通哺乳动物肠道中厌氧菌占绝对优势的种类并不相同,但乳酸杆菌和双歧杆菌仍然是水生生物的生理性有益菌,对维持肠道平衡起到重要作用(王红宁等,1994),推测浒苔在水生动物中同样能够调节肠道健康,促进消化吸收,这与本实验结果一致。
3.4 超微粉碎浒苔对花鲈血清非特性免疫指标的影响浒苔中含有酚类、萜类、多糖类、卤化物、含硫化合物等活性物质,具有较强的抗菌活性和抗病毒性。研究表明(Castro et al,2004)浒苔多糖能够提高大菱鲆的呼吸活性,通过增强中性粒细胞和巨噬细胞的活力进而提高机体免疫力,肠浒苔对多种真菌具有抑制活性(Rizvi et al,2000),扁浒苔能够抑制细菌感染以及消除炎症(Venkata et al,2004),浒苔多糖能够提高小鼠的体液免疫、细胞免疫以及单核吞噬系统功能(Wei et al,2014),也有研究表明浒苔多糖可以明显地促进T、B 淋巴细胞的体外增殖,对诱导IFN-γ的产生有非常明显的增强作用,具有增强机体免疫反应的作用(徐大伦等,2005a)。研究发现,大黄鱼肌肉中的3 种抗氧化酶活性、谷胱甘肽(GSH)和MDA(丙二醛)含量以及总抗氧化能力(T-AOC)均明显低于肝脏中,反映了肝脏对清除自由基具有重要作用,在抗氧化调节方面起到主要作用(张晓丽,2013)。本实验中,浒苔对花鲈肝脏中的几种非特异性免疫酶活性有比较明显的影响,其中AKP 活性最小值为对照组,最大值为Diet 3(3%),且差异显著(P<0.05),当肝功能受到损伤时,GOT和GPT 活性就会升高,本实验中,GPT 跟GOT 的均在对照组活力最大,分别在Diet 4和Diet 3 呈现最小,说明超微粉碎浒苔具有保护肝脏的作用,而TP 随着添加量的增多其活性逐渐增大,直至Diet 3和Diet 4 达到最大,肝脏中T-SOD 活性没有显著变化,但血清T-SOD 活性有显著升高,在Diet 3 达到最大,血清AKP 活性在Diet 2 及以上添加组有显著升高,血清LSZ 在Diet 3 及以上添加组有显著升高。T-SOD 是重要的抗氧化酶之一,在清除活性氧自由基,防止生物分子损伤方面有重要的作用,本实验中浒苔使花鲈血清T-SOD 活性显著提高,原因可能为浒苔多糖能够与受体结合,刺激机体产生免疫反应,从而有效地清除羟自由基(郭子叶等,2014)。AKP是溶酶体的一种含金属的标志性酶,催化磷酸单酯水解成乙醇和磷酸,是动物体内参与免疫防御等活动的重要的水解酶(Pipe,1990)。LSZ 是一种碱性蛋白,能够水解细菌细胞壁上的肽聚糖,担负起机体防御的功能,徐大伦等(2005b)研究发现向扇贝体内注射浒苔多糖后,扇贝体内SOD 酶和LSZ 活性均有显著的提高作用,说明浒苔多糖能够促进扇贝的免疫活性,本实验中花鲈的AKP 及LSZ 均有明显提高,同样说明浒苔能够明显促进花鲈的非特异性免疫功能。
综上所述: 花鲈饲料中添加超微粉碎浒苔可以明显促进其生长性能,提高非特异性免疫力及肠道中三种消化酶活力,降低花鲈肥满度并对胃消化酶产生一定影响,建议超微粉碎浒苔在饲料中最适添加量为3%。
| 王红宁, 何明清, 柳 苹等. 1994. 鲤肠道正常菌群的研究.水生生物学报, 18(4): 354-359 |
| 王述柏, 史雪萍, 周传凤等. 2013a. 浒苔添加水平对肉鸡生产性能、胴体品质及小肠消化酶活性的影响. 动物营养学报,25(6): 1332-1337 |
| 王述柏, 贾玉辉, 王利华等. 2013b. 浒苔添加水平对蛋鸡产蛋性能、蛋品质、免疫功能及粪便微生物区系的影响. 动物营养学报, 25(6): 1346-1352 |
| 王晓兰, 关洪斌, 田 甜. 2014. 3 种新型配方饲料对大菱鲆诱食和生长的影响. 饲料研究, (1): 50-54 |
| 冯昭信. 1998. 花鲈研究. 北京: 海洋出版社, 1 |
| 刘树立, 王 华. 2007. 超微粉碎技术的优势及应用进展. 干燥技术与设备, 5(1): 35-38 |
| 刘智禹. 2012. 浒苔对海水中重金属的富集研究及食用安全风险评估. 福建水产, 34(1): 71-75 |
| 闫杰, 张 欣, 孔晓静等. 2012. 饵料中浒苔添加量对梭鱼生长影响效果研究. 饲料研究, (3): 64-65, 71 |
| 孙建凤, 宋宏立, 赵 军等. 2010. 饲粮中不同浒苔水平对肉鸡养分利用率和消化酶活性的影响. 动物营养学报, 22(6):1658-1664 |
| 李铁梁, 罗 琳, 马志宏等. 2013. 超微粉碎技术对中草药制剂的增效作用研究. 饲料工业, 34(21): 47-51 |
| 杨欢, 黎中宝, 李元跃等. 2013. 厦门海域浒苔种类鉴定及其营养成分分析. 中国渔业质量与标准, 3(4): 70-75 |
| 杨毅雯. 2013. 紫茉莉对酒精性肝损伤大鼠ALT、AST、T-SOD影响的研究. 大理: 大理学院硕士学位论文 |
| 张晓丽, 胡玉珍, 李明云等. 2013. 养殖大黄鱼在自然海区降温不同阶段抗氧化水平及血清酶活性的变化. 海洋科学,37(11): 27-34 |
| 林建云, 陈维芬, 贺 青等. 2011. 福建沿岸海域浒苔藻类的营养成分含量与食用安全. 台湾海峡, 30(4): 570-576 |
| 周胜强, 游翠红, 王树启等. 2013. 饲料中添加浒苔对黄斑蓝子鱼生长性能与生理生化指标的影响. 中国水产科学,20(6): 1257-1265 |
| 赵瑞祯. 2012. 石莼饲料对鲈鱼生长影响的研究. 饲料研究,(8): 58-60 |
| 郝凤奇, 尹 航, 李冠桥等. 2008. 添加海藻提取物对育肥牛蛋白质消化率的影响. 饲料与畜牧, (1): 44-45 |
| 徐大伦, 黄晓春, 欧昌荣等. 2005a. 浒苔多糖对非特异性免疫功能的体外实验研究. 食品科学, 26(10): 232-235 |
| 徐大伦, 黄晓春, 欧昌荣等. 2005b. 浒苔多糖对扇贝SOD 酶和溶菌酶活力的影响. 水利渔业, 25(3): 22-23 |
| 郭子叶, 蔡春尔, 耿中雷等. 2014. 浒苔粗多糖化妆品开发潜力研究. 中国海洋药物, 33(2): 57-62 |
| 曾凡归. 2010. 黄斑蓝子鱼对大型海藻的选择性及其影响因素.汕头: 汕头大学硕士学位论文 |
| 曾呈奎, 张德瑞, 张俊甫. 1962. 中国经济海藻志. 北京: 科学出版社, 43-50 |
| 解康, 关洪斌, 冯文利等. 2012. 浒苔在鱼类养殖中深加工工艺的研究. 饲料工业, 33(10): 22-24 |
| Asino H M. 2010. 饲料中添加浒苔Enteromorpha prolifera 对大黄鱼生长性能的影响. 青岛: 中国海洋大学硕士学位论文 |
| Castro R, Zarra I, Lamas J. 2004. Water-soluble seaweed extractsmodulate the respiratory burst activity of turbot phagocytes.Aquaculture, 229(1-4): 67-78 |
| Du Z Y, Liu Y J, Tian L X et al. 2006. The influence of feeding rate on growth, feed efficiency and body composition ofjuvenile grass carp (Ctenopharyngodon idella). AquacultureInternational, 14(3): 247-257 |
| Hagen Ø, Solberg C, Sirnes E et al. 2007. Biochemical andstructural factors contributing to seasonal variation in thetexture of farmed Atlantic Halibut (Hippoglossushippoglossus L.) flesh. Journal of Agricultural and FoodChemistry, 55(14): 5803-5808 |
| Pipe R K. 1990. Hydrolytic enzymes associated with the granularhaemocytes of the marine mussel Mytilus edulis. TheHistochemical Journal, 22(11): 595-603 |
| Rizvi M A, Farooqui S, Shameel M. 2000. Bioactivity andelemental composition of certain seaweeds from Karachicoast. Pakistan Journal of Marine Biology, 6(2): 207-218 |
| Thilagam H, Gopalakrishnan S, Bo J et al. 2014. Comparativestudy of 17 β-estradiol on endocrine disruption andbiotransformation in fingerlings and juveniles of Japanesesea bass Lateolabrax japonicus. Marine Pollution Bulletin,85(2): 332-337 |
| Venkata Raman B, Rao D N, Radhakrishnan T M. 2004.Enteromorpha compressa (L.) Greville an edible green algaas a source of antiallergic principle (S). Indian Journal ofClinical Biochemistry, 19(1): 105-109 |
| Wei J T, Wang S X, Liu G et al. 2014. Polysaccharides fromEnteromorpha prolifera enhance the immunity of normal mice.International Journal of Biological Macromolecules, 64: 1-5 |
| Yıldırım Ö, Ergün S, Yaman S et al. 2009. Effects of twoseaweeds (Ulva lactuca and Enteromorpha linza) as a feedadditive in diets on growth performance, feed utilization,and body composition of rainbow trout (Oncorhynchusmykiss). Kafkas Üniversitesi Veteriner Fakültesi Dergisi,15(3): 455-460 |
| Yousif O M, Osman M F, Anawhi A R et al. 2004. Growthresponse and carcass composition of rabbitfish, Siganuscanaliculatus (Park) fed diets supplemented with dehydratedseaweed, Enteromorpha sp. Emirates Journal of Food andAgriculture, 16(2): 18-26 |
| Yu H H, Han F, Xue M et al. 2014. Efficacy and tolerance of yeastcell wall as an immunostimulant in the diet of Japaneseseabass (Lateolabrax japonicus). Aquaculture, 432: 217-224 |
| Zhang Y Q, Wu Y B, Jiang D L et al. 2014. Gamma-irradiatedsoybean meal replaced more fish meal in the diets ofJapanese seabass (Lateolabrax japonicus). Animal FeedScience and Technology, 197: 155-163 |