 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 江兴龙, 林国清, 廖碧钗, 郑伟刚, 黄健, 邓来富, 尼逸伦. 2015.
- JIANG Xing-Long, LIN Guo-Qing, LIAO Bi-Chai, ZHENG Wei-Gang, HUANG Jian, DENG Lai-Fu, NI Yi-Lun. 2015.
- 缢蛏(Sinonovacula constricta)规模增殖放流对三沙湾滩涂底质环境改良的效果
- EVALUATION ON JUVENILE SINONOVACULA CONSTRICTA RELEASE FOR STOCK ENHANCEMENT AND SEDIMENT IMPROVEMENT IN SANSHA BAY
- 海洋与湖沼, 46(6): 1524-1530
- Oceanologia et Limnologia Sinica, 46(6): 1524-1530.
- http://dx.doi.org/10.11693/hyhz20150700193
-
文章历史
- 收稿日期:2015-07-22
- 改回日期:2015-08-19


2. 福建省水产技术推广总站 福州 350003;
3. 鳗鲡现代产业技术教育部工程研究中心 厦门 361021
2. Fujian Fishery Technical Extension Center, Fuzhou 350003, China;
3. Engineering Research Center of the Modern Industry Technology for Eel Ministry of Education, Xiamen 361021, China
渔业资源增殖放流作为一项通过向自然水域投放鱼虾蟹贝类亲体、人工繁育苗种或经暂养的野生苗种来恢复渔业资源,实现渔业可持续发展的管理手段,为当前国内外在水生生物资源养护和水域生态修复领域普遍采用(Bartley et al,2008; Lorenzen,2008)。近年来,由于经济建设发展的需要,近海及江河、湖泊成为开发建设的重要区域,密集频繁的水上施工作业、大面积的围海围湖造田、过度的渔业捕捞以及工业污染物、废弃物、生活垃圾的大量排放,使我国近海和内陆水域(江河、湖泊等)的渔业资源严重衰退,水域环境恶化(隋然等,2014)。针对水生生物资源与水域生态环境所面临的问题,近年来我国各地加大了增殖放流工作力度,资源养护管理工作呈现出新的良好局面,增殖放流己在全国逐渐成为转变渔业发展方式、提高渔民收入、维护渔区社会稳定的重要手段(李继龙等,2009)。贝类人工增殖是人工集约化开发浅海滩涂的重要途径,也是促进浅海滩涂生态平衡的措施(黄东万,2004)。缢蛏(Sinonovacula constricta Lamarck)属于软体动物门、瓣鳃纲、帘蛤目、竹蛏科,是一种广温性埋栖型滩涂贝类,广泛分布于我国沿海,是我国四大贝类养殖品种之一(王如才等,1993)。三沙湾位于福建省东北部沿海,它由盐田港、白马港、卢门港、东吾洋、官井洋和福鼎洋等组成,是一个近封闭型海湾,仅在东南方向有一个宽约 3km的东冲口与东海相通。其潮汐形态为非正规半日潮,属于大潮差海区(刘修德,2009)。盐田港畔的福安市溪尾镇溪邳村,拥有海域面积约333 hm2和滩涂面积约133 hm2,近年来随着海洋开发利用强度加大,周边滩涂湿地的生态服务功能受到一定的影响(杨妙峰等,2015),特别随着陆源污染的加剧,滩涂底质日益老化,滩涂贝类养殖环境日益恶化。为修复溪邳村滩涂底质环境,恢复原有贝类种类缢蛏的种群数量和渔业资源量,达到恢复滩涂生物多样性的目的,于2013—2014 年开展了缢蛏规模增殖放流。本研究通过开展对放流缢蛏生长、生物量动态变化和滩涂底质环境主要因子的跟踪监测,探讨缢蛏增殖放流与修复滩涂底质环境间的关系,评估缢蛏增殖放流的综合效果,为今后科学开展滩涂贝类增殖放流提供指导。
1 材料与方法1.1 放流苗种来源缢蛏放流苗种为购自福建泉州的海区培育苗种。
1.2 放流地点和方法于2013 年4 月24 日和2014 年3 月17 日,分别在三沙湾的福安市溪尾镇溪邳村滩涂开展缢蛏增殖放流,选择中潮带中线以下滩涂,滩涂放流增殖区面积约59810m2,分为二个增殖片区(增殖区Ⅰ区和增殖区Ⅱ区,见图 1),每区面积约3 公顷。缢蛏苗用塑料框装运到现场放流,放流前进行总量称重并随机抽取30 个样品,进行生物学测定,测定体重、壳长、壳宽等指标。放流方法采用抛播缢蛏苗。

|
| 图 1 缢蛏增殖放流区位置 Fig. 1 The areas of artificial release for stock enhancement of S. constricta |
缢蛏增殖放流后每隔3 个月开展1 次跟踪监测,其中,第1 批次缢蛏增殖放流的最后一次跟踪监测抽采样时间为2014 年1 月22 日,放流天数273 天; 第2 批次缢蛏增殖放流的最后一次跟踪监测抽采样时间为2015 年1 月30 日,放流天数为319 天。对增殖放流滩涂区域,设24 个采样位点,在增殖区Ⅰ区、增殖区Ⅱ区及对照区内,各设8 个面积均为1m2 的采样点,开展缢蛏采样,采样时用小铲将底泥沙连同缢蛏取出,厚度约20cm,随着缢蛏的生长所取泥沙的厚度逐渐增加,将取得的海泥装入水桶运回,用80 目或40 目筛绢网将泥沙滤出,从冲净的剩余物中将缢蛏捡出,然后进行生物学测定,测定体重、壳长、壳宽等指标; 在增殖区Ⅰ区、增殖区Ⅱ区及对照区内,各设4 个站点,采集泥样,监测滩涂底泥表层的氧化还原电位、有机碳及硫化物含量。氧化还原电位的现场测定方法、硫化物和有机碳样品的采集和检测方法等都按《海洋监测规范》(GB 17378-2007)规定的方法进行(国家海洋局,2008)。
1.4 数据处理与分析应用SAS 9.1 统计分析软件(SAS Institute Inc.,Cary,NC,USA),对对照区与增殖区的试验数据进行显著水平ɑ=0.05 的t-test 检验,以P<0.05 为显著差异,P<0.01 为极显著差异。
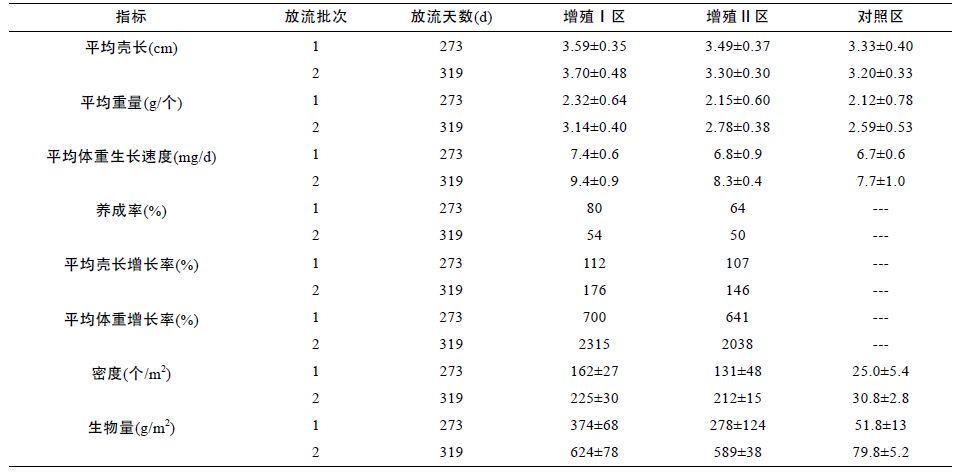
2 结果2.1 缢蛏增殖放流批次、规格和数量于2013 年4 月24 日,开展了第一批次的缢蛏增殖放流,放流缢蛏苗种111 框,总重量3607.5kg,计1244 万粒缢蛏苗; 缢蛏体重0.14—0.53g,平均体重0.29g; 壳长1.2—2.3cm,平均壳长1.69cm; 壳宽0.4—0.8cm,平均壳宽0.6cm。于2014 年3 月17 日,开展了第二批次的缢蛏增殖放流,放流缢蛏苗种95 框,总重量3254kg,计2503 万粒缢蛏苗; 缢蛏体重0.03—0.43g,平均体重0.13g; 壳长0.6—2.0cm,平均壳长1.34cm。数据见表 1。

第1 批次的缢蛏增殖放流结果,根据2014 年1 月22 日的随机抽采样检测结果,放流273 天,增殖放流区Ⅰ区的缢蛏密度和缢蛏生物量(g/m2)分别比对照区的极显著提高了约548%和622%(P<0.01),缢蛏平均壳长、平均个体重量分别比对照区的显著提高了约8%、9%(P<0.05),缢蛏生长速度比对照区的显著提高了11%(P<0.05),缢蛏的养成成活率约80%; 增殖放流区Ⅱ区的缢蛏密度和缢蛏生物量(g/m2)分别比对照区的极显著提高了约424%和437%(P<0.01),缢蛏平均壳长比对照区的显著提高了约5%(P<0.05),平均个体重量比对照区的提高了约1%(P>0.05),缢蛏生长速度与对照区的无显著差异(P>0.05),缢蛏的增殖养成率约64%(表 2,图 2,图 3)。在开展第二批次的缢蛏增殖放流前,增殖Ⅰ区、Ⅱ区总收获缢蛏中苗的重量计28800kg,产值约32 万元,第1 批次的增殖放流产出比为1 : 8.0。
 |

|
| 图 2 第一批次螠蛏放流生长情况 Fig. 2 The growth curve of S. constricta in the first batch of release |

|
| 图 3 第一批次螠蛏放流生物量情况 Fig. 3 The biomass curve of S. constricta in the first batch of release |
第2 批次的缢蛏增殖放流结果,根据2015 年1月30 日的随机抽采样检测结果,放流319 天,增殖放流区Ⅰ区的缢蛏密度和缢蛏生物量(g/m2)分别比对照区的极显著提高了约631%和682%(P<0.01),缢蛏平均壳长和平均个体重量分别比对照区的显著提高了约16%和21%(P<0.05),缢蛏生长速度比对照区的显著提高了22%(P<0.05),缢蛏的养成成活率约54%; Ⅱ区的缢蛏密度和缢蛏生物量(g/m2)分别比对照区的极显著提高了约588%和638%(P<0.01),平均壳长和平均个体重量分别比对照区的提高了约3%和7%(P>0.05),缢蛏生长速度与对照区的无显著差异(P>0.05),缢蛏的增殖养成率约50%(表 2,图 4,图 5)。增殖示范区Ⅰ区、Ⅱ区总收获的缢蛏中苗的重量计36245kg,产值约41 万元,第2 批次的增殖放流产出比为1 : 9.0。

|
| 图 4 第 2 批次螠蛏放流生长情况 Fig. 4 S. constricta growth curve of the second batch of release |

|
| 图 5 第 2 批次螠蛏放流生物量情况 Fig. 5 S. constricta biomass curve of the second batch of release |
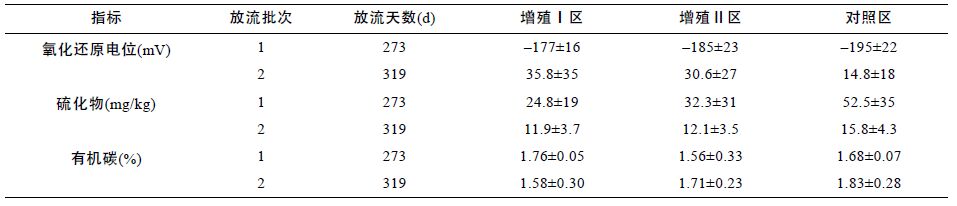
第 1 批次螠蛏放流前,底泥中的氧化还原电位、硫化物和有机碳等的本底浓度在增殖区Ⅰ区、Ⅱ区和对照区之间都无显著差异(P>0.05)(表 3)。从表 4 可知,在第1 批次的放流期间,底泥中的氧化还原电位、硫化物和有机碳等浓度在增殖区Ⅰ区、Ⅱ区和对照区之间都无显著差异(P>0.05),但,底泥中Ⅰ区、Ⅱ区的氧化还原电位平均值分别比对照区提高了9.0%和5.1%,硫化物浓度平均值分别比对照区降低了53%和38%,有机碳浓度平均值Ⅰ区比对照区提高了5.0%,Ⅱ区比对照区下降了7.2%(P>0.05); 在第2 批次的放流期间,底泥中的氧化还原电位、硫化物和有机碳等浓度在增殖区Ⅰ区、Ⅱ区和对照区之间都无显著差异(P>0.05),但,底泥中Ⅰ区、Ⅱ区的氧化还原电位平均值分别比对照区提高了141%和107%,硫化物浓度平均值分别比对照区降低了24%和23%,有机碳浓度平均值分别比对照区下降了13%和6.6%(P>0.05); 在增殖区Ⅰ区、Ⅱ区和对照区,第2 批次放流期间底泥中的氧化还原电位值比第1 批次放流期间的值分别极显著提高了120%、117%和108%(P<0.01)(变化趋势见图 6)。在Ⅰ区和Ⅱ区,第2 批次放流期间底泥中的硫化物浓度比第1 批次放流期间的分别降低了52%和62%(P>0.05),对照区中,第2 批次放流期间底泥中的硫化物浓度比第1 批次放流期间的显著降低了70%(P<0.05)(变化趋势见图 7)。在Ⅰ区第2 批次放流期间底泥中的有机碳浓度比第1 批次放流期间的降低了10%(P>0.05),在Ⅱ区和对照区,第2 批次放流期间底泥中的有机碳浓度比第1 批次放流期间的分别提高了9.5%和8.9%(P>0.05)(变化趋势见图 8)。
 |
 |

|
| 图 6 放流期间增殖区与对照区底质氧化还原电位的月际变化 Fig. 6 The monthly variation in oxidation-reduction potential in the artificial release and control areas |

|
| 图 7 放流期间增殖区与对照区底质硫化物的月际变化 Fig. 7 The monthly variation in sulfide concentration of the sediment in the artificial release and control areas |

|
| 图 8 放流期间增殖区与对照区底质有机碳的月际变化 Fig. 8 The monthly variation in organic carbon concentration in the artificial release and control areas |
从表 5 可知,在第 1 批次和第 2 批次的螠蛏放流期间,放流天数全程为644 天,底泥中的氧化还原电位、硫化物和有机碳等浓度在增殖区Ⅰ区、Ⅱ区和对照区之间都无显著差异(P>0.05),但,底泥中Ⅰ区、Ⅱ区的氧化还原电位平均值分别比对照区提高了26%和18%,硫化物浓度平均值分别比对照区降低了45%和34%,有机碳浓度平均值分别比对照区下降了5.8%和6.8%(P>0.05)。此外,底泥中Ⅰ区的氧化还原电位平均值比Ⅱ区提高了10%,硫化物浓度平均值比Ⅱ区降低了16%,有机碳浓度平均值比Ⅱ区提高了1%(P>0.05)。

从第1 批次和第2 批次的缢蛏增殖放流结果可知,增殖放流区Ⅰ区和Ⅱ区的缢蛏密度和缢蛏生物量(g/m2)都比对照区有极显著提高,并且第2 批次的增幅都高于第1 批次,Ⅰ区的增幅都高于Ⅱ区; 第1 批次和第2 批次中均仅Ⅰ区的缢蛏生长速度分别比对照区的显著提高了11%和22%,而Ⅱ区的缢蛏生长速度与对照区均无显著差异。表明,在该海区的缢蛏增殖放流,第2 批次的缢蛏资源增殖效果优于第1 批次,增殖放流区Ⅰ区的缢蛏资源增殖效果优于Ⅱ区。从实际获得的增殖放流投入产出比中,第1 批次的增殖放流产出比为1 : 8.0,而第2 批次的增殖放流产出比为1 : 9.0,明显高于第1 批次,表明第2 批次的增殖放流资源增殖效果和经济效益优于第1 批次。原因认为主要是第2 批次的缢蛏增殖放流的生物密度达418.5 个/m2,高于第1 批次的207.3 个/m2 导致; 此外,虽然第2 批次的缢蛏放流规格低于第1 批次,以及放流天数多于第1 批次,导致第2 批次放流的养成成活率低于第1批次。但综合作用的结果,表现为第2 批次的增殖放流资源增殖效果和经济效益优于第1 批次,也表明缢蛏增殖放流合理密度的确定和放流苗种规格的选择都是影响资源增殖效果的重要因素。合理确定放流数量至关重要,若放流数量过少,将达不到放流效果;若放流的数量过多,不仅会增加放流的成本,而且还造成了放流水域环境的压力,也会影响苗种的生长(梁君,2013)。放流苗种规格对苗种的放流效果有很大影响,放流最适规格的苗种,苗种成活率高、培育时间相对较短,可以节省放流成本,从而提升经济效益(徐炳庆,2011)。
3.2 底质环境改良效果分析由于投放的贝类、多毛类的摄食与活动,使无机盐从底质到水体加速释放,对促进滩涂水域浮游植物增长、提高初级生产力及降低有机质、腐植质都十分有利,对改善底质结构、增加底质透气性、调节氧化还原电位、最终改善沉积物的理化性质、提高环境容纳量,均起到良好的重要作用(陈惠彬,2005)。本研究中,通过增殖放流缢蛏,由于缢蛏的摄食与潜沙活动等,增加了对底质的扰动和透气性,有利于底泥氧化还原电位的提高,降低硫化物浓度和有机碳浓度。监测结果,缢蛏总体放流期间,底泥中增殖放流Ⅰ区、Ⅱ区的氧化还原电位平均值分别比对照区提高了26%和18%,硫化物浓度平均值分别比对照区降低了45%和34%,有机碳浓度平均值分别比对照区下降了5.8%和6.8%。这也表明,缢蛏放流对滩涂底质中氧化还原电位的提高和硫化物浓度的降低具有较明显改良作用,而对有机碳浓度的降低作用较小。此外,第2 批次放流期间增殖放流Ⅰ区、Ⅱ区底泥中的氧化还原电位值比第1 批次放流期间的值分别极显著提高了120%和117%,硫化物浓度比第1 批次放流期间的分别降低了52%和62%,也表明,随着缢蛏放流活动的持续进行,在第1 批次放流对滩涂底质环境因子改良的基础上,第2 批次放流期间,产生了改良的叠加效应,表现为对底质环境的改良效果获得更大幅度的递增。利用生物的方法对受损的生态系统进行修复,国内外学者都做了大量的工作(Prichard et al,1992;Sandhya et al,1992; 李秋芬等,2000; 江兴龙,2012;江兴龙等,2013),但修复的对象主要是受污水体,应用贝类增殖放流的手段对滩涂底质环境开展修复的研究较少报道,本研究虽然取得了一些对滩涂底质环境因子改良的初步效果,但尚未获得显著的底质环境修复效果,今后还需要深入开展相关研究。另一方面,滩涂底质环境的改良也影响放流缢蛏的生命活动、种群增长和资源增殖效果。缢蛏属埋栖类贝类,其生长和存活除受到水质状况的制约外,与底质环境的质量密切相关,滩涂底质影响放流缢蛏的产量,缢蛏产量与底质中硫化物和有机碳的平均含量大致呈线性负相关关系,底质中硫化物及有机碳含量过高会使缢蛏产量下降(陈聚法等,2005)。本研究放流期间,增殖放流Ⅰ区底泥中的氧化还原电位平均值比Ⅱ区提高了10%,硫化物浓度平均值比Ⅱ区降低了16%,而有机碳浓度平均值仅比Ⅱ区高1%,表明放流期间Ⅰ区的总体底质环境优于Ⅱ区。这也促使增殖放流区Ⅰ区的缢蛏资源增殖效果优于Ⅱ区。
| 王如才, 王昭平, 张建中. 1993. 海水贝类养殖学. 青岛: 青岛海洋大学出版社, 72-75 |
| 刘修德. 2009. 福建省海湾数模与环境研究--三沙湾. 北京:海洋出版社 |
| 江兴龙. 2012. 日本鳗鲡(Anguilla japonica)土池生物膜原位修复低碳养殖技术的研究. 海洋与湖沼, 43(6): 1134-1140 |
| 江兴龙, 邓来富. 2013. 凡纳滨对虾(Litopenaeus vannamei)池塘生物膜低碳养殖技术研究. 海洋与湖沼, 44(6):1536-1543 |
| 李秋芬, 袁有宪. 2000. 海水养殖环境生物修复技术研究展望.中国水产科学, 7(2): 90-92 |
| 李继龙, 王国伟, 杨文波等. 2009. 国外渔业资源增殖放流状况及其对我国的启示. 中国渔业经济, 27(3): 111-123 |
| 杨妙峰, 郑盛华, 林永青等. 2015. 三沙湾溪邳村滩涂养殖区沉积物质量状况及风险评价. 福建水产, 37(3): 202-210 |
| 陈惠彬. 2005. 渤海典型海岸带滩涂生境、生物资源修复技术研究与示范. 海洋环境保护, (3): 20-23 |
| 陈聚法, 张东杰, 宋建中等. 2005. 乳山湾缢蛏养殖老化滩涂的修复研究. 海洋水产研究, 26(5): 57-61 |
| 国家海洋局. 2008. GB 17378.1-7-2007 海洋监测规范. 北京:中国标准出版社 |
| 徐炳庆. 2011. 山东近海中国对虾增殖放流的研究. 上海: 上海海洋大学硕士学位论文 |
| 黄东万. 2004. 贝类护养增殖高产技术. 广西水产科技, (4):44-46 |
| 梁君. 2013. 海洋渔业资源增殖放流效果的主要影响因素及对策研究. 中国渔业经济, 31(5): 122-134 |
| 隋 然, 马 欣, 张 勇等. 2014. 目前我国渔业资源增殖放流中存在的问题及建议. 中国水产, (4): 29-30 |
| Bartley D M, Bell J D. 2008. Restocking, stock enhancement,and sea ranching: arenas of progress. Rev Fish Sci, 16(1-3):357-365 |
| Lorenzen K. 2008. Understanding and managing enhancementfisheries systems. Rev Fish Sci, 16(1-3): 10-23 |
| Prichard P H, Mueller J G, Rogers J C et al. 1992. Oil spillbioremediation: experiences, lessons and results from theExxon Valdex oil spill in Alaska. Biodegradation, 3(2-3):315-335 |
| Sandhya G, Singhal P K, Hasija S K. 1992. Relative contributionsof bacteria and fungi to water hyacinth decomposition.Aquatic Botany, 43(1): 1-15 |