 2015, Vol. 46
2015, Vol. 46中国海洋湖沼学会主办。
文章信息
- 高瞻, 陈强, 黎中宝, 李文静, 黄永春, 徐安乐, 杨敏. 2015.
- GAO Zhan, CHEN Qiang, LI Zhong-Bao, LI Wen-Jing, HUANG Yong-Chun, XU An-Le, YANG Min. 2015.
- 饲料添加发酵浒苔对花鲈(Lateolabrax japonicus) 生长性能、消化酶活性和免疫的影响
- EFFECTS OF FERMENTED ENTEROMORPHA PROLIFERA ON GROWTH PERFORMANCE, DIGESTIVE ENZYME ACTIVITIES AND SERUM NON-SPECIFIC IMMUNITY OF LATEOLABRAX JAPONICUS
- 海洋与湖沼, 46(6): 1549-1556
- Oceanologia et Limnologia Sinica, 46(6): 1549-1556.
- http://dx.doi.org/10.11693/hyhz20150200057
-
文章历史
- 收稿日期:2015-02-14
- 改回日期:2015-10-31




2. 福建省海洋渔业资源与生态环境重点实验室 厦门 361021
2. Fujian Provincial Key Laboratory of Marine Fishery Resources and Eco-environment, Xiamen 361021, China
花鲈(Lateolabrax japonicus)又称鲈鱼、七星鲈等,属硬骨鱼纲(Osteichsthyes)、鲈形目(Perciformes)、鮨科(Servanidae)、花鲈属(Lateolabrax)。花鲈是广温广盐性鱼类,在日本、朝鲜沿海以及我国沿岸海域均有分布(张春丹等,2005)。花鲈肉质爽滑鲜美,味道可口。其生长快,适应能力强,淡海水均能养殖,是一种经济、营养价值极高的鱼类,也是我国重要的养殖鱼类。迄今关于花鲈的营养研究主要有鱼粉替代(Hu et al,2013)、维生素需求量(张璐等,2015)以及添加玉米蛋白粉(Men et al,2014)等,但关于浒苔对花鲈的营养研究还未见报道。
近几十年来,我国作为世界水产大国,随着集约化水平的不断提高,水产品的产量大大超过了全球生产总量的1/3,长达60 年之久的海水鱼人工养殖已取得巨大成就(Hong et al,2003; Cao et al,2015)。但渔业的过度开发与利用已经引发了一系列问题,如鱼类病害,药物残留以及环境污染等。研究新型绿色抗病促长添加剂开始日益得到关注,也已经成为调节鱼体营养、增强鱼体免疫性的有效途径(黎中宝,2004)。
浒苔(Enteromorpha prolifera)俗称苔条、青海苔等,属于大型绿藻,隶属于绿藻门(Chlorophyta)、石莼目(Ulvales)、石莼科(Ulvaceae)、浒苔属(Enteromorpha)(Hiraoka et al,2003; 杨欢等,2013)。浒苔营养物质极其丰富,富含氨基酸、矿质元素等,具有极高的药用和食用价值。近年来,海洋环境污染加剧,浒苔大量繁殖,绿潮频频暴发(Nelson et al,2008)。绿潮的频发不仅会改变海洋生态环境和海洋生物多样性,还会严重污染水质。因此,如何使得浒苔资源被合理有效利用,改善生态环境问题,非常值得研究。而国内外也已经有一系列关于浒苔作为饲料添加剂在养殖业中的报道,如虹鳟(Oncorhynchus mykiss)(Onder et al,2009),大黄鱼(Larimichthys crocea)幼鱼(Asino et al,2011),大菱鲆(Scophthalmus maximus)(卢青等,2015)以及梭鱼(Liza haematocheila)(闫杰等,2012)等。以上研究均表明,饲料添加浒苔对水生动物具有显著促长,增强免疫力的作用。而浒苔在花鲈中的应用研究未见报道。故本试验利用发酵浒苔作为添加剂,探求其对花鲈生长、消化以及免疫性能的影响,为发酵浒苔在鲈鱼饲料中的应用提供理论依据。
发酵技术是指利用工程技术手段,直接用微生物参与,或者利用菌体细胞或酶的某些功能,通过其代谢活动将底料转化成我们所需要的产物的一项技术(祝玉洪等,2013)。我国利用微生物发酵来酿酒早有应用,后在中草药也有大量研究。随着发酵技术的日益成熟,发酵技术在饲料行业中应用越来越广泛。因其可以增加原料风味、改善适口性、提高营养价值而受到欢迎。本试验初步尝试开发利用发酵浒苔,这对使浒苔成为具有经济价值的海藻类饲料也有着重要意义。
1 材料与方法1.1 试验用鱼及驯养试验所用花鲈购于福建省漳浦县锦兴育苗场。置于集美大学海水试验场循环过滤桶(1200L)中暂养14d。期间每天两次定时(7: 30—8: 00,16: 30—17: 00)投喂基础饲料,并逐渐将其淡化至纯淡水养殖。
1.2 试验设计及试验饲料花鲈所用基础饲料购于厦门市嘉康饲料厂,其主要成分包括鱼粉、豆粕、面粉、卵磷脂、维生素和矿物质等(主要营养成分见表 1)。所选用的浒苔于2014 年3—5 月从厦门龙舟池采集,将采集到的浒苔用海水冲洗干净,自然晾干。直至浒苔表面无水分后置于60°C 烘箱中进行烘干,之后在冷冻超微粉碎机中进行粉碎,过200 目筛制得粉碎好的浒苔粉。将粉碎好的浒苔粉加入2‰的二型饲料发酵助剂(购于北京华夏康源科技有限公司),1%的玉米粉作为能量饲料,同时加入65%的蒸馏水,装入规格为350*450mm,厚度为10 丝的发酵袋内,密封发酵3—4 d。得到的发酵浒苔闭封保存备用。将发酵浒苔与基础饲料按等差数列设计成比例,按逐级扩大法混匀,利用催化成型机配制成对照组(0%)、Diet 1(1%)、Diet 2(2%)、Diet 3(3%)、Diet 4(4%)、Diet 5(5%)六种不同发酵浒苔添加水平的颗粒型饲料,风干2—3 d,于自封袋内–20°C 冰箱保存备用。
在暂养结束后挑选出体格健壮、规格一致的花鲈苗540 尾[初始体重为(6.40±0.14)g],随机分配到18 个设有循环过滤水系统的玻璃水族缸(40cm × 40cm × 70cm)中,共分为6 个处理组,每处理组3 个重复。在试验期间每天同样两次定时(7: 30—8: 00,16: 30—17: 00)饱食投喂。喂食30min 后收集残饵,烘干,称量,确保得到准确饲料摄入量。然后吸粪换水,保持24h 不间断充气增氧。试验期间水温(26±1)°C、pH(8.0±0.2)、溶氧≥7 mg/L、氨氮浓度<0.2 mg/L。每天观察记录花鲈摄食与生长状况,记录水温以及死亡情况。
1.4 样品采集及处理饲喂28d 后,将花鲈饥饿24h,采用丁香酚麻醉,对各试验缸鱼进行整体称重和计数。每缸随机捞取5尾鱼测体质量和体长,采用1mL 无菌注射器对其进行尾部静脉取血。将收集得到的血样静置12h 后在3000r/min 条件下离心10min,收集上层血清置–80°C冰箱保存用于分析血清生化指标。同时将其进行解剖,快速分离肝脏,胃,肠道。将肝脏称重以计算肝体指数,然后分装入冻存管投入液氮后转移至–80°C 冰箱中保存备用。每缸随机选取3 只花鲈保存于–80°C 冰箱用于全鱼体成分的测定。
1.5 测定指标 1.5.1 生长指标分析对试验鱼生长性能指标测定时所用公式为:
增重率WG(%)= 100×(鱼体末重–鱼体初重)/鱼体初重;
饲料系数FCR(%)=均摄食量/(终末体质量–初始体质量);
存活率SR(%)= 100×(收获尾数/放养尾数);
肝体指数HSI(%)= 100×(肝胰脏重/鱼体质量);
肥满度CF(%)= 100×体质量(g)/体长3(cm3)
鱼体称湿重后于70°C 烘至恒重,再次称重。水分的测定采用105°C 常压烘箱干燥恒重法。将烘干的鱼体绞碎,采用半微量凯氏定氮法(总氮×6.25)测粗蛋白; 索氏抽提法(乙醚为抽提液)测粗脂肪; 马福炉灼烧法(550°C)测灰分。
1.5.3 肠、胃消化酶活性测定将胃和肠道称重,按1 : 9(V/W)加入4°C 的0.85%生理盐水,在冰水浴中匀浆5min。采用低温高速离心机(4°C,3000r/min,15min)对匀浆液进行离心,吸取上清于–80°C 冰箱保存待测消化酶。测定指标有胰蛋白酶、胃蛋白酶、肠、胃道淀粉酶(AMS)、脂肪酶(LPS)活性,测定所用相应试剂盒均由南京建成生物工程研究所生产,操作步骤按照说明书进行。
1.5.4 血清、肝脏免疫酶活性测定将肝脏称重,加入9 倍体积(V/W)的85%生理盐水(4°C),用匀浆器冰水浴匀浆5 min,再用低温高速冷冻离心机3000 r/min离心15 min,取上清液制成20%的组织匀浆。测定指标有血清、肝脏总超氧化物歧化酶(T-SOD)、溶菌酶(LZM)和碱性磷酸酶(AKP)活性均采用南京建成生物工程研究所生产的相应试剂盒测定,操作步骤按照说明书进行。
1.5.5 统计方法试验所得数据均用SPSS19.0 统计软件进行单因素方差分析(One-way ANOVA),利用Duncan 法进行多重比较,取显著水平P<0.05。试验所得数据均以平均数±标准误进行表示。
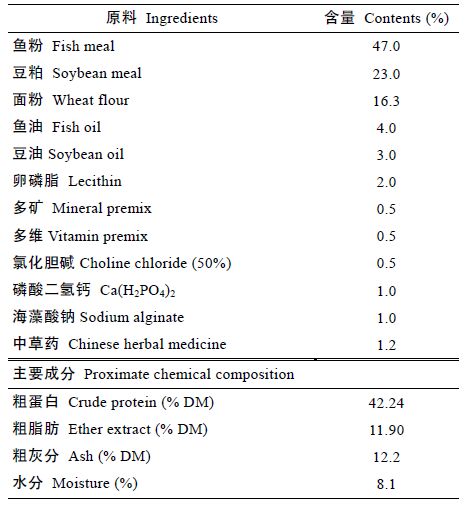
2 结果2.1 发酵浒苔对花鲈生长性能的影响发酵浒苔对花鲈生长性能影响见表 2,饲料中添加发酵浒苔显著提高了花鲈的增重率和特定生长率(P<0.05)(表 3)。生长实验结果表明添加水平在Diet2(2%)时增重率和特定生长率达到最大,比对照组分别显著提高了45.2%、18.2%(P<0.05),但添加组与对照组相比不存在显著差异(P>0.05)。随发酵浒苔添加浓度的上升,各实验组花鲈饲料系数呈现先降低后升高的趋势,Diet 3(3%)组花鲈的饲料系数显著低于对照组16.6%(P<0.05)。各组肥满度均无显著差异(P>0.05),各组存活率均为100%。
 |
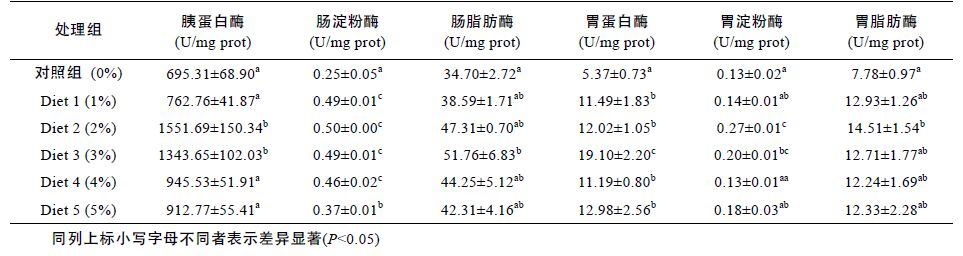
由表 3 可知,以特定生长率、饲料系数为指标作最佳拟合曲线,花鲈鱼饲料中发酵浒苔的最佳添加水平分别为2.91%、2.71%。

由表 4 可知: 添加发酵浒苔可适当降低鱼体中的水分,提高灰分含量但并不显著(P>0.05)。同时各添加组的粗脂肪和粗蛋白含量均高于对照组,各添加组之间并无显著性差异(P>0.05)。
 |
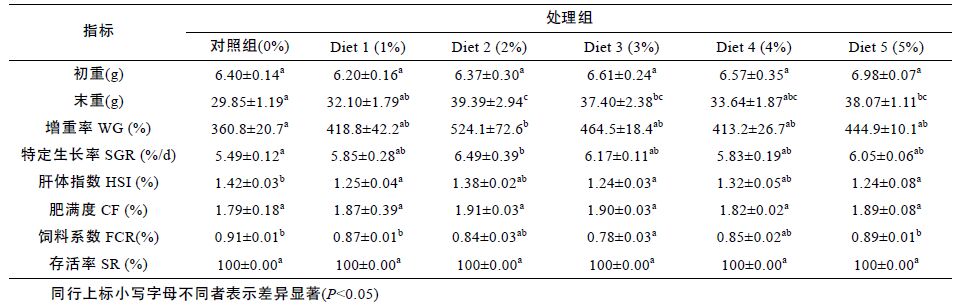
由表 5 可以看出,在发酵浒苔添加浓度为2%—3%时胰蛋白酶,肠淀粉酶,肠脂肪酶活性明显高于其余各组(P<0.05),随着添加浓度的继续上升,呈现出下降的趋势。胰蛋白酶在添加水平Diet 2(2%)时活性达到最高。肠淀粉酶和脂肪酶则在Diet 3(3%)时达到最高,处理组各组差异不显著(P<0.05),且均高于对照组(P>0.05)。
 |
花鲈摄食试验料后对胃中消化酶活性影响见表 5,添加组中花鲈胃蛋白酶、淀粉酶、脂肪酶均得到显著的提高(P<0.05),且随着发酵浒苔添加浓度的上升呈现先上升后下降的趋在添加水平为Diet 3(3%)时胃蛋白酶活性最高,且各处理组与对照组差异性显著(P<0.05)。各组定粉酶,脂肪酶在添加水平为Diet 2(2%)时活性最高,各添加组均显著高于对照组(P<0.05),但各添加组之间的脂肪酶并无显著差异(P>0.05)。在发酵浒苔添加量为2%—3%浓度范围时花鲈的胃消化酶活性达到最高且显著高于对照组(P<0.05)。
表 6 为以各消化酶为指标所得出的拟合曲线方程,由表可知花鲈鱼饲料中发酵浒苔的最佳添加水平为2.50%—3.00%。
发酵浒苔对花鲈血清免疫酶的活性影响见表 7。由表可知,饲料中添加发酵浒苔可提高处理组鲈鱼血清的碱性磷酸酶活性,在Diet 2(2%)组活性达到最大,但差异不显著(P>0.05)。随着添加发酵浒苔浓度的不断增高,鲈鱼血清中溶菌酶的活性呈现先上升后逐渐下降的趋势,在Diet 2(2%)组活性达到最大,并且显著高于其余各组(P<0.05)。添加组血清中总超氧化物歧化酶活性显著高于对照组(P<0.05),但添加组各组之间差异不显著(P<0.05)。
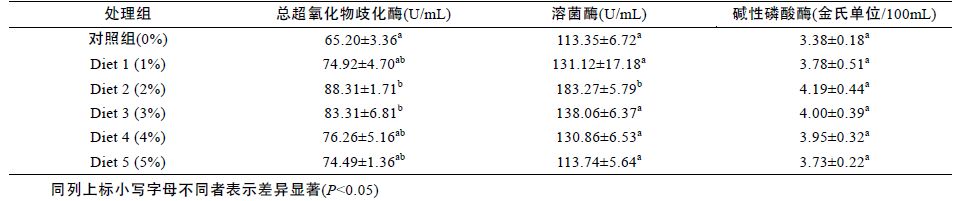 |
由表 8 可知,饲料中添加发酵浒苔可显著提高添加组鲈鱼肝的碱性磷酸酶活性(P<0.05),在Diet 2(2%)组活性达到最大,但各添加组之间差异不显著(P>0.05)。在添加浓度为2%—3%之间花鲈肝的溶菌酶活性显著高于其余各组(P<0.05),其余添加组与对照组之间不存在差异性(P>0.05)。随着添加发酵浒苔浓度的不断增高,鲈鱼肝中总超氧化氢歧化酶的活性呈现先上升后逐渐下降的趋势,在Diet 2(2%)组活性达到最大,并且显著高于其余各组(P<0.05)。
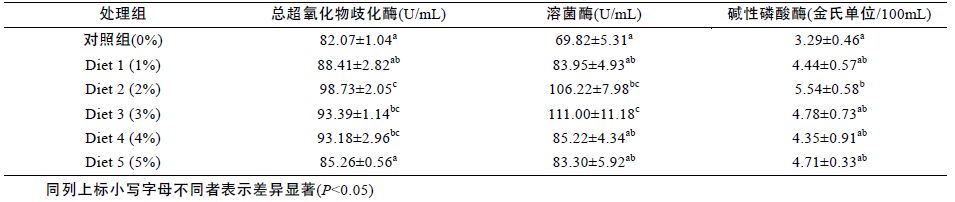 |
表 9 为以各免疫酶为指标所得出的拟合曲线方程,由表可知花鲈鱼饲料中发酵浒苔的最佳添加水平为2.41%—3.00%。
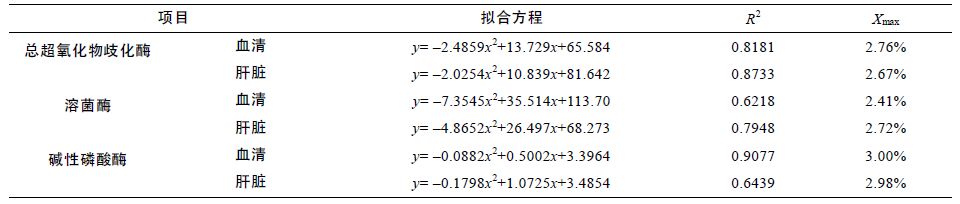
浒苔是一种优质的大型天然海藻,含有丰富的蛋白质、矿物质、维生素等,营养价值及其利用率都非常高。Asino等(2011)研究了饲料中添加浒苔对大黄鱼幼鱼生长的促进作用,研究结果表明添加浒苔可以显著提高其特定生长率,存活率均达到97%以上。闫杰等(2012)在饵料中添加浒苔探究对梭鱼(Liza haematocheila)生长效果影响,结果表明添加浒苔组可以显著提高梭鱼的特定生长率,降低饲料系数。王晓兰等(2014)在大菱鲆(Scophthalmus maximus)配合饲料中添加复方中草药提取液、L-甘氨酸和干浒苔粉,结果表明浒苔添加组的大菱鲆的增重率得到了提高。在本试验中,浒苔添加组显著提高了鲈鱼的生长性能,并且在一定添加浓度内,鲈鱼的增重率和特定生长率逐渐上升,其原因可能是由于浒苔蛋白质、不饱和脂肪酸含量高,脂肪含量低,且含有生物活性物质如多糖、萜类和甾体等(李晓等,2013),具有增强免疫力,调节机体代谢,促进鲈鱼生长的作用。Yıldırım等(2009)探究了将10%的浒苔粉作为配合饲料添加对虹鳟(Oncorhynchus mykiss)生长效果的影响,结果表明添加10%浒苔粉会降低鱼的生长速度和饲料利用率。在Yousif等(2004)对黄斑蓝子鱼饲料中添加浒苔的研究中也发现在添加量为10%时可以提高鱼体增重率。但随着添加量由10%、20%到30%的增加,鱼体的增重率、生长效率会逐渐降低,同时提高饲料系数。这些结果与本试验中得到的一致。虽然在本试验中发酵浒苔的添加量最高只达到5%,但随着浒苔添加量的增多而降低原因可能是由于添加量过大而降低了饲料适口性所致,但具体研究机制仍需进一步探索。杨春花等(2012)对海藻粉进行发酵后再添加到基础日粮中饲喂1 日龄樱桃谷肉鸭(Cherry Valley Ducks),研究结果表明其大大高肉鸭采食量、日增重以及降低饲料系数。本试验对浒苔进行发酵后对花鲈进行研究同样提高了花鲈的增重率,降低了饲料系数。分析其原因可能是在发酵时破坏了浒苔所含的胶质所致,使营养物质更容易被吸收,避免了因利用率低而造成的浪费。
另外,在本试验中添加浒苔可以降低鲈鱼的肝体指数、鱼体水分含量,提高粗蛋白和粗灰分含量。从而在一定程度上改善鱼肉品质,提高产品质量。在Asino等(2011)的研究结果中添加浒苔可以提高其粗脂肪、粗灰分含量,结果基本一致。
3.2 发酵浒苔对花鲈胃、肠道消化酶的影响鱼类消化酶活性反应的是鱼类对营养物质的吸收利用情况,同一种消化酶在不同消化器官中分布将存在差异。微生物发酵不仅可以改变饲料理化性质,还可以降解底物中的大分子有机物,从而使消化利用率得到提高(李猛等,2015)。通过微生物发酵产物作为饲料添加剂研究花鲈的消化酶活性的试验并不多见,本试验在一定程度上弥补了类似研究的不足。本试验证明对浒苔进行发酵后,不但提高了饲料的适口性,而且也显著提高花鲈的消化性能。Wang等(2006)、Suzer等(2008)研究表明,消化性能的增强可能是提高鱼类生长率一个原因。杨春花等(2012)的研究中,将海藻粉发酵后饲喂1 日龄樱桃谷肉鸭(Cherry Valley Ducks)后发现,由于发酵工艺降低了浒苔的纤维素含量,使得浒苔所含营养物质更加利于肉鸭的消化吸收,进而提高了肉鸭的消化性能,因此显著提高了其生长速度。孙建凤等(2010)在肉鸡饲粮中添加浒苔,结果发现肉鸡的胃蛋白酶、胰蛋白酶以及淀粉酶活性都得到了显著提高。王述柏等(2013)研究表明饲料中添加4%的浒苔可以提高肉鸡肠淀粉酶、脂肪酶以及蛋白酶活性。这均与本试验结果较为一致。夏素银等(2010)利用发酵蛋白饲料替代豆粕研究其对肉仔鸡消化吸收的影响,发现提高了其养分消化率。本试验所研究的浒苔就是一种高蛋白的海藻,故该研究结果对本试验具有一定的参考价值。秦搏等(2015)发现添加浒苔粉可以显著提高幼刺参(Apostichopus japonicus Selenka)的淀粉酶和胃蛋白酶,其中以添加水平25%含量为最优。这与本试验中2%含量为最佳相差较大,究其原因一方面可能是因为研究对象不同而差异,另一方面可能是因为浒苔经过发酵后大大提高了其利用率,故使其最适添加水平得以降低。
3.3 发酵浒苔对花鲈血清、肝脏免疫指标的影响鱼类生存环境具有特殊性,因此在抵抗病害方面非特异性免疫机制比特异性免疫显得更为重要(Anderson,1992)。本研究选取超氧化物歧化酶(SOD)、溶菌酶(LZM)和碱性磷酸酶(AKP)这三种具有代表性的非特异性免疫酶指标对花鲈非特异性免疫性能进行说明。超氧化物歧化酶反应了机体清除多余自由基的能力,是一种重要的抗氧化酶。溶菌酶(LZM)具有较强杀菌作用,可促进机体细胞的吞噬活性,可在机体启动免疫保护时发挥作用。本试验在基础饲料中添加不同水平的发酵浒苔进行投喂后,结果表明添加发酵浒苔可以明显提高花鲈血清、肝脏的免疫性能。徐大伦等(2005)研究发现向扇贝(Chlamys farreri))体内注射浒苔多糖后,扇贝体内超氧化物歧化酶和溶菌酶活性得到明显提高,进一步论证了浒苔具有促进机体免疫活性的作用。Castro等(2004)研究证实绿藻类的浒苔能够提高大菱鲆的呼吸活性,从而加强噬菌细胞中巨噬细胞杀死细菌病原体的能力。唐薇等(2014)在研究中发现浒苔粉能促进刺参组织的免疫性能。王述柏等(2013)在日粮中添加浒苔粉显著提高了肉鸡的免疫力。这些结果都说明了添加浒苔具有提高机体免疫性能的作用,与本试验结果较为吻合。卢青等(2015)采用浒苔粉作为饲料添加剂对大菱鲆(Scophthalmus maximus)的试验研究得出,当添加量在5%时并不会提高大菱鲆的免疫力,且随着添加量继续增高反而抑制其免疫活性。在本试验中,在发酵浒苔在添加量为5%仍可增加机体免疫力,且在添加量为3%时达到最佳。同时也随着添加量的增高呈降低趋势,这点与卢青的研究相符合。但就添加量5%能否提高鱼体免疫力上与卢青的研究不一致,原因可能是卢青等采用的是浒苔粉,而本试验采用的是发酵浒苔,发酵浒苔中的发酵菌随底物进入机体环境,并作为外源微生物激发免疫,增加了具有杀菌作用的体液因子,从而有效提高机体免疫力。同样在姜燕(2014)研究实验中发现,与未发酵的饲料相比发酵饲料能提高刺参(Apostichopus japonicus Selenka)的超氧化物歧化酶、溶菌酶等非特异性免疫酶的活性。由此可以推知,对浒苔进行发酵处理所得到的效果可能好于未经过发酵处理的浒苔。
4 结论在本试验条件下,饲料中添加发酵浒苔能够促进花鲈的生长,降低花鲈鱼体水分含量,在一定程度改善花鲈鱼肉品质。还可以提高饲料利用率,增强花鲈胃、肠道消化酶活性从而改善花鲈鱼的生长状况。并且对花鲈免疫力也有显著提高,增强花鲈抗病能力。
通过生长指标,消化指标以及免疫指标等重要指标的最佳拟合曲线可得知,在添加浓度为2%—3%时能显著提高其生长性能,消化性能和免疫性能。
| 王述柏, 史雪萍, 周传凤等. 2013. 浒苔添加水平对肉鸡生产性能、胴体品质及小肠消化酶活性的影响. 动物营养学报,25(6): 1332-1337 |
| 王晓兰, 关洪斌, 田甜. 2014. 3 种新型配方饲料对大菱鲆诱食和生长的影响. 饲料研究, (1): 50-54 |
| 卢青, 杨宁, 王正丽等. 2015. 饲料中添加浒苔对大菱鲆生长和非特异性免疫力的影响. 青岛农业大学学报(自然科学版), 32(1): 62-66 |
| 闫杰, 张欣, 孔晓静等. 2012. 饵料中浒苔添加量对梭鱼生长影响效果研究. 饲料研究, (3): 64-65, 71 |
| 孙建凤, 宋宏立, 赵军等. 2010. 饲粮中不同浒苔水平对肉鸡养分利用率和消化酶活性的影响. 动物营养学报,22(6): 1658-1664 |
| 李晓, 王颖, 吴志宏等. 2013. 浒苔对刺参幼参生长影响的初步研究. 中国水产科学, 20(5): 1092-1099 |
| 李猛, 廖梅杰, 王印庚等. 2015. 浒苔添加比例与微生物发酵对幼刺参生长、消化和非特异性免疫的影响. 动物营养学报, 27(10): 3270-3278 |
| 杨欢, 黎中宝, 李元跃等. 2013. 厦门海域浒苔种类鉴定及其营养成分分析. 中国渔业质量与标准, 3(4): 70-75 |
| 杨春花, 方希修. 2012. 海藻发酵饲料对樱桃谷肉鸭生长性能的影响. 黑龙江畜牧兽医, (4): 81-82 |
| 张璐, 李静, 谭芳芳等. 2015, 饲料中不同维生素A 含量对花鲈生长和血清生化指标的影响. 水产学报, 1: 88-96 |
| 张春丹, 李明云. 2005. 花鲈繁殖生物学及繁育技术研究进展.宁波大学学报(理工版), 18(3): 400-403 |
| 姜燕. 2014. 刺参(Apostichopus japonicus)发酵饲料的制作工艺与应用效果研究. 青岛: 中国海洋大学博士学位论文,43-72 |
| 祝玉洪, 马立周. 2013. 发酵技术在饲料行业中的应用. 饲料工业, 34(18): 49-54 |
| 秦搏, 常青, 陈四清等. 2015. 饲料中浒苔添加量以及处理方法对幼刺参生长、消化率、消化酶和非特异性免疫酶的影响. 水产学报, 39(4): 547-556 |
| 夏素银, 王成章, 严学兵等. 2010. 发酵蛋白饲料替代豆粕对肉仔鸡生长性能、养分消化率及肠道菌群的影响. 动物营养学报, 22(2): 352-357 |
| 徐大伦, 黄晓春, 欧昌荣等. 2005. 浒苔多糖对扇贝SOD 酶和溶菌酶活力的影响. 水利渔业, 25(3): 22-23 |
| 唐薇, 王庆吉, 张蕾等. 2014. 不同藻粉对刺参组织免疫性能和体壁成分的影响. 资源开发与市场, 30(8): 905-907, 920 |
| 黎中宝. 2004. 饲料添加剂. 厦门: 厦门大学出版社, 160-164 |
| Anderson D P. 1992. Immunostimulants, adjuvants, and vaccinecarriers in fish: applications to aquaculture. Annual Reviewof Fish Diseases, 2: 281-307 |
| Asino H, Ai Q H, Mai K S et al. 2011. Evaluation ofEnteromorpha prolifera as a feed component in large yellowcroaker (Pseudosciaena crocea, Richardson. 1846) diets.Aquaculture Research, 42(4): 525-533 |
| Cao L, Naylor R, Henriksson P et al. 2015. China's aquacultureand the world's wild fisheries. Science, 347(6218):133-135 |
| Castro R, Zarra I, Larras J. 2004. Water-soluble seaweed extractsmodulate the respiratory burst activity of turbot phagocytes.Aquaculture, 229(1-4): 67-68 |
| Hiraoka M, Dan A, Shimada S et al. 2003. Different life historiesof Enteromorpha prolifera (Ulvales, Chlorophyta) from fourrivers on Shikoku Island, Japan. Phycologia, 42(3): 275-284 |
| Hong W S, Zhang Q Y. 2003. Review of captive bred speciesand fry production of marine fish in China. Aquaculture,227(1-4): 305-318 |
| Hu L, Yun B, Xue M et al. 2013. Effects of fish meal quality andfish meal substitution by animal protein blend on growthperformance, flesh quality and liver histology of Japaneseseabass (Lateolabrax japonicus). Aquaculture, 372: 52-61 |
| Men K K, Ai Q H, Mai K S et al. 2014. effects of dietary corngluten meal on growth, digestion and rotein metabolism inrelation to IGF-I gene expression of Japanese seabass,Lateolabrax japonicus. Aquaculture, 428: 303-309 |
| Nelson T A, Haberlin K, Nelson A V et al. 2008. Ecological andphysiological controls of species composition in greenmacroalgal blooms. Ecology, 89(5): 1287-1298/a> |
| Suzer C, Çoban D, Kamaci H O et al. 2008. Lactobacillus spp.bacteria as probiotics in gilthead sea bream (Sparus aurat L.)larvae: Effects on growth performance and digestive enzymeactivities. Aquaculture, 280(1-4): 140-145 |
| Wang Y B, Xu Z R. 2006. Effect of probiotics for common carp(Cyprinus carpio) based on growth performance anddigestive enzyme activities. Animal Feed Science andTechnology, 127(3-4): 283-292 |
| Yıldırım Ö, Ergün S, Yaman S et al. 2009. Effects of twoseaweeds (Ulva lactuca and Enteromorpha linza) as a feedadditive in diets on growth performance, feed utilization,and body composition of rainbow trout (Oncorhynchusmykiss). Kafkas Üniversitesi Veteriner Fakültesi Dergisi,15(3): 455-460 |
| Yousif O M, Osman M F, Anawhi A R et al. 2004. Growthresponse and carcass composition of rabbitfish, Siganuscanaliculatus (Park) fed diets supplemented with dehydratedseaweed, Enteromorpha sp. Emirates Journal of Food andAgriculture, 16(2): 18-26 |