 2016, Vol. 47
2016, Vol. 47

十溴联苯醚(BDE-209)胁迫对褶皱臂尾轮虫(Brachionus plicatilis)摄食与选食的影响

http://dx.doi.org/10.11693/hyhz20150500152
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 陈红梅, 唐学玺, 房宽, 王悠. 2016.
- CHEN Hong-Mei, TANG Xue-Xi, FANG Kuan, WANG You. 2016.
- 十溴联苯醚(BDE-209)胁迫对褶皱臂尾轮虫(Brachionus plicatilis)摄食与选食的影响
- ACUTE TOXIC EFFECT OF BDE-209 ON FEEDING BEHAVIOR OF ROTIFER BRACHIONUS PLICATILIS
- 海洋与湖沼, 47(2): 438-446
- Oceanologia et Limnologia Sinica, 47(2): 438-446.
- http://dx.doi.org/10.11693/hyhz20150500152
-
文章历史
- 收稿日期:2015-05-28
- 改回日期:2015-08-27
引用本文
陈红梅,
唐学玺,
房宽,
王悠. 2016. 十溴联苯醚(BDE-209)胁迫对褶皱臂尾轮虫(Brachionus plicatilis)摄食与选食的影响[J]. 海洋与湖沼, 47(2): 438-446
CHEN Hong-Mei,
TANG Xue-Xi,
FANG Kuan,
WANG You. 2016. ACUTE TOXIC EFFECT OF BDE-209 ON FEEDING BEHAVIOR OF ROTIFER BRACHIONUS PLICATILIS[J]. Oceanologia et Limnologia Sinica, 47(2): 438-446.
十溴联苯醚(BDE-209)胁迫对褶皱臂尾轮虫(Brachionus plicatilis)摄食与选食的影响
中国海洋大学海洋生命学院 青岛 266003
摘要:
实验生态条件下研究十溴联苯醚(BDE-209)的急性毒性胁迫对褶皱臂尾轮虫(Branchionus plicatilis)运动行为与摄食行为的影响, 探讨可用以表征胁迫-响应的生物标志物。结果表明:(1) BDE-209胁迫24h显著性增加了褶皱臂尾轮虫的运动抑制率, 由此得到24h半数有效抑制浓度(24h-EC50)为9.084mg/L, 但未达到半数致死作用;(2) 当分别以四种海洋微藻: 小球藻(Chlorella vulgaris)、青岛大扁藻(Platymonas helgolandica)、等鞭金藻8701(Isochrysis galbana)和小新月菱形藻(Cylindrotheca closterium)为单一饵料来源时, 褶皱臂尾轮虫对四种饵料单胞藻的滤水率(F)和摄食率(G)均随着胁迫时间和胁迫浓度的增加而不断下降, 呈现出明显的浓度依赖性, 说明BDE-209胁迫能显著抑制褶皱臂尾轮虫的摄食作用。(3) 将其中三种微藻等生物量混合投喂时, 褶皱臂尾轮虫对三种微藻的滤水率和摄食率与单一饵料投喂条件相比均发生变化, 且对各种微藻的选食率(Sr)和摄食总生物量均随着胁迫时间和浓度的增加呈下降趋势, 选择性指数(Si)发生变化, 对小球藻的摄食选择性最高。滤水率的变化较其它指标更敏感, 可用以指示浮游动物在个体水平上响应外源环境变化的标志物。结果表明, BDE-209胁迫能有效改变褶皱臂尾轮虫的摄食与选食行为, 且其最适宜的饵料是小球藻。
关键词:
BDE-209
褶皱臂尾轮虫
饵料微藻
急性毒性
摄食
选食
ACUTE TOXIC EFFECT OF BDE-209 ON FEEDING BEHAVIOR OF ROTIFER BRACHIONUS PLICATILIS
College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China
Abstract:
Under controlled laboratory conditions, the acute toxic effect of high brominated Deca-BDE (BDE-209) on Brachionus plicatilis and the response of feeding behavior to the stress were studied. Experiment was designed at five concentrations of BDE-209 (2, 6, 10, 14, and 18mg/L) against a blank and a control groups in triplication. The results indicated that: (1) BDE-209 significantly inhibited the motile behavior of B. plicatilis and the 24 h-EC50 was 9.084mg/L, but the half lethal effects were not observed; (2) When four species of marine microalgae bait: Chlorella vulgaris, Platymonas helgolandica, Isochrysis galbana, and Cylindrotheca closterium were fed as the single food source, the filtering rate (F) and the feeding rate (G) of B. plicatilis decreased obviously as the increase of BDE-209 concentrations and exposure time, exhibiting a does-dependent manner, which indicated the feeding behavior of B. plicatilis was significantly inhibited by BDE-209; (3) when C. vulgaris, P. helgolandica, and I. Galbana were fed to B. plicatilis together as mixed food, the values of F, G, feeding selectivity ratio (Sr) and total biomass of B. plicatilis decrsased obviously in the same manner, and the food selectivity index (Si) changed simultaneously. C. vulgaris was the optimal food source for B. plicatilis with and without BDE-209 exposure. Moreover, the filtering rate was the most sensitive parameter to the stress and could be used as an indicator of zooplankton to the changes of ambient environment. We concluded that the stress of BDE-209 significantly changed the feeding and selecting behavior of B. plicatilis, and the optimal food source for it was C. vulgaris.
Key words:
BDE-209
Brachionus plicatilis
bait marine microalgae
acute toxicity
feeding activity
feeding selectivity
多溴联苯醚(polybrominated diphenyl ethers,PBDEs)作为溴化阻燃剂被广泛应用于多种消费产品中(Talsness,2008)。它化学性质稳定,亲脂性较高,可在生物体内蓄积放大并随食物链传递而产生生物毒性效应,是一类具有高生态风险性的新型持久性有机污染物(persistent organic pollutants,POPs)(Tanabe,2004)。近年来,PBDEs陆续在海水、海洋沉积物和海洋生物体等各种介质中被检出且呈持续增长趋势(孟范平等,2009),且在深海与两极的海洋环境中也发现有PBDEs的分布。PBDEs对海洋生态环境的潜在威胁引起人们的高度重视。有关PBDEs对海洋生物的毒性效应已有较多的研究,主要集中于哺乳动物或脊椎动物如鱼类的研究中,发现其具有明显的生殖毒性、发育毒性以及内分泌干扰毒性 (Chen et al,2012; McClain et al,2012),对低等海洋浮游生物的研究较少。海洋浮游动物是影响海洋生态系统物质循环与能量流动的关键生物类群,而摄食作用能直接影响浮游动物的种群动态变化; 且有研究表明海洋浮游动物是PBDEs进入海洋生态环境的起点(Chiuchiolo et al,2004),因此研究PBDEs对海洋浮游动物摄食行为的影响对阐明PBDEs的海洋生态学效应具有重要意义。
海洋浮游动物褶皱臂尾轮虫(Branchionus plicatilis)是我国沿海浮游动物的广布种,常被用作环境监测和生态毒理学研究的受试生物(Suatoni et al,2006)。本实验室前期的研究结果表明,褶皱臂尾轮虫幼虫对饵料微藻具有明显的摄食选择性(房宽等,2013)。但是外源污染物是否会影响其摄食行为,改变其摄食选择性尚不清楚。因此,本研究开展高溴代多溴联苯醚——十溴联苯醚(BDE-209)胁迫对褶皱臂尾轮虫摄食与选食行为的影响,以期为阐明 BDE-209的海洋生态学潜在影响提供一定的科学依据,探讨建立基于浮游动物的、能灵敏指示环境胁迫变化的生物标志物。
1 材料与方法 1.1 实验动物培养取适量褶皱臂尾轮虫(B. plicatilis)休眠卵(由中国海洋大学水产学院惠赠)进行孵化。孵化条件: 温度(20 ± 1)°C,海水盐度30,pH 8.6,光照60μmol/(m2·s),光暗比12L︰12D,3—7d即可孵化。孵化期间投喂密度为1×106cell/mL的小球藻作为饵料。孵化后挑选一活泼健壮的非混交卵雌体于恒温光照培养箱内按上述条件培养2个月待用。
收集12—14h的幼虫(龄期<14h)进行实验,期间不投喂其它饵料。实验选取绿藻门的小球藻(Chlorella vulgaris)、青岛大扁藻(Platymonas helgolandica)、金藻门的等鞭金藻8701(Isochrysis galbana)和硅藻门的小新月菱形藻(Cylindrotheca closterium)四种单胞藻为饵料。微藻培养于f/2培养基中,培养条件同褶皱臂尾轮虫,每隔12h摇动一次。
1.2 实验试剂实验用多溴联苯醚——十溴联苯醚(BDE-209,纯度99.05%),购自美国AccuStandard公司,为难溶于水的白色粉末。在预实验中选择了三种助溶剂: 丙酮、异辛烷和二甲亚砜(DMSO),分别测定三种助溶剂对褶皱臂尾轮虫幼虫24h的急性毒性致死实验的最小无明显效应浓度(no observable effect concentration,NOEC),其NOEC值的大小为异辛烷<丙酮<DMSO(表 1); 同时发现DMSO的助溶效果最佳,因此选为本实验的助溶剂。
表 1 三种助溶剂对褶皱臂尾轮虫24h的NOEC值实验结果
Tab. 1 Determination of NOEC of three cosolvents on B. plicatilis at 24h
| 助溶剂 | 回归方程 | 95%置信区间(%) | R2 | NOEC(%) |
| 丙酮 | y=9.05x-6.43 | 1.32—4.50 | 0.98 | 3.71 |
| 异辛烷 | y=3.54x-0.01 | 0.30—0.55 | 0.99 | 0.44 |
| DMSO | y=18.82x-22.57 | 12.64—14.10 | 0.97 | 13.52 |
| NOEC: 最小无明显效应浓度 | ||||
用DMSO(A.R.)配制成浓度为2×103mg/L的BDE-209-DMSO母液,4°C下避光保存备用。实验时用DMSO稀释母液至中间浓度后加入至实验体系中。
1.3 实验设计 1.3.1 BDE-209对褶皱臂尾轮虫的急性毒性效应如1.1所述的筛选方法选取轮虫并随机分组。实验共分5个BDE-209浓度组: 2、6、10、14和18mg/L,同时设空白对照组(海水对照组)和助溶剂对照组(0.9% DMSO,V/V),每组设3个平行。实验选取10mL的烧杯,总体积为5mL,其中含有20只褶皱臂尾轮虫。胁迫12h及24h后在解剖镜下观察,同时记录褶皱臂尾轮虫的致死率和运动抑制率。通过概率单位-对数浓度法计算BDE-209的24h-LC50或EC50。轮虫死亡判定标准: ①轮虫停止运动,②内部组织出现皱缩,③咀嚼囊和足均不运动,④轮盘收回则认为其死亡; 轮虫运动抑制判定标准: 若轻摇烧杯15s之内轮虫不运动则认为其运动能力受到抑制(房宽等,2013)。
1.3.2 BDE-209胁迫对褶皱臂尾轮虫摄食的影响(1) 对滤水率(F)和摄食率(G)的影响 将轮虫分别置于小球藻(密度为10×105cell/mL)、青岛大扁藻(密度为2×105cell/mL)、等鞭金藻8701(密度为15×105cell/mL)、小新月菱形藻(密度为4.5×105cell/mL)中驯养48h,其中4株海洋微藻的投喂密度均选取各饵料微藻的饱和密度(周文礼,2005)。实验用轮虫收集活泼健壮且龄期<2h的带卵雌体,饥饿12h后进行实验。在急性毒性实验24h-EC50的基础上设定摄食实验的浓度: 实验共设低、中、高3个浓度组,分别为: 0.0375、0.15和0.6mg/L,并设空白对照组(海水对照组)和DMSO助溶剂(0.3‰,V/V)的阴性对照组,每组设3个平行。实验在10mL烧杯中进行,总体积为5mL,每实验体系中放入50只轮虫幼虫,然后在黑暗条件下置于转速为100r/min的恒温震荡器上孵育,确保饵料藻处于均匀状态的同时防止藻类迅速增殖。胁迫12h及24h后用鲁哥氏碘液固定,显微镜下血球计数板计数,根据Frost公式(Frost,1972)采用饵料密度差减法计算其F和G:
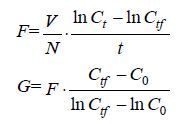
其中,F表示滤水率[mL/(ind.·h)]; G表示摄食率[cell/(ind.·h)]; V表示实验溶液体积; N表示轮虫个体数(ind.); C0表示起始饵料密度(×104/mL); Ct表示无轮虫组中的最终饵料密度(×104cell/mL); Ctf表示实验组中的最终饵料密度(×104cell/mL); t表示摄食时间。
(2) 对选食作用的影响 在前期实验的基础上,选择小球藻、青岛大扁藻和小新月菱形藻3种微藻,这三种微藻形态差异明显且易在显微镜下甄别,将其按等生物量混合作为饵料用以培养褶皱臂尾轮虫。由于三种微藻的个体大小不同,因此根据本实验室前期研究结果(房宽等,2013),混合饵料中微藻起始密度分别设定为: 小球藻33.33×104cell/mL、青岛大扁藻4.80×104cell/mL、等鞭金藻8701为14.14× 104cell/mL。轮虫的选择、培养条件及实验方法同上。
根据混合饵料中各种微藻密度的变化计算褶皱臂尾轮虫的选食率(Sr)和选择性指数(Si)(Frost,1972):
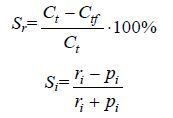
其中,Sr: 轮虫对某种单胞藻选食率; Si: 轮虫对某种单胞藻摄食的选择性指数(Si,0< Si <1表示喜食,Si =0表示对食物无选择性; -1< Si <0表示不喜食; Ct: 无轮虫组中最终饵料密度(×104cell/mL); Ctf: 实验组中最终饵料密度(×104cell/mL); ri: 轮虫所摄食的全部藻中某种单胞藻所占的比例; pi: 实验最初某种单胞藻在全部单胞藻中的比例。
1.4 数据分析采用SPSS 19.0软件通过单因素方差分析(one- way ANOVA)各作用组和对照组间差异,P<0.05表示差异显著。采用Sigmaplot 10.0软件作图。
2 结果 2.1 BDE-209的急性毒性胁迫对褶皱臂尾轮虫致死率与运动抑制率的影响随着BDE-209浓度的增加褶皱臂尾轮虫的死亡率增加,但24h时高浓度组(18mg/L)的死亡率为33.33%,未达到半数致死作用(表 2)。与死亡率不同,BDE-209胁迫能有效抑制褶皱臂尾轮虫的运动,并且随着胁迫浓度的增高运动抑制作用不断增强。根据胁迫组中个体运动抑制率的变化,通过概率单位法得到BDE-209对褶皱臂尾轮虫的24h-EC50为9.084mg/L(y=3.22x-3.08,R2=0.98)(表 2)。
表 2 BDE-209的急性毒性胁迫对褶皱臂尾轮虫致死率与运动抑制率的影响
Tab. 2 Acute toxic effect of BDE-209 on lethality and motile inhibition of B. plicatilis
| 浓度(mg/L) | 致死率(%) | 运动抑制率(%) |
| 0 | 0 | 0 |
| 0.9% DMSO | 0 | 0 |
| 2 | 0 | 3.33 |
| 6 | 3.33 | 23.33 |
| 10 | 16.67 | 53.33 |
| 14 | 23.33 | 76.67 |
| 18 | 33.33 | 83.33 |
根据国家环保总局《中华人民共和国环境保护行业标准—新化学物质危害评估导则》(国家环保总局,2004)中的毒性分级标准规定: LC50/EC50在1.0—10mg/L之间的为高毒性物质,据此标准BDE-209属高毒性物质。
2.2 单一饵料投喂条件下BDE-209胁迫对褶皱臂尾轮虫滤水率(F)和摄食率(G)的影响当投喂四种单一饵料,BDE-209胁迫12h、24h时对褶皱臂尾轮虫滤水率(F)均随胁迫浓度的升高呈现出不断下降的趋势,与对照组相比差异显著(P<0.05),且24h时F的下降程度明显高于12h时的影响(图 1)。通过概率单位法计算BDE-209胁迫24h时褶皱臂尾轮虫对四种饵料微藻滤水率的EC50分别为0.54、0.51、0.33、0.46mg/L,说明BDE-209胁迫对褶皱臂尾轮虫摄食上述四种微藻的影响程度为: 小球藻<青岛大扁藻<小新月菱形藻<等鞭金藻8701。但胁迫前后褶皱臂尾轮虫对各种饵料藻F的大小顺序未发生变化,均为等鞭金藻8701<青岛大扁藻<小球藻<小新月菱形藻。实验结果说明BDE- 209胁迫影响褶皱臂尾轮虫对不同种类饵料微藻的摄食行为。
 |
| 图 1 单一饵料微藻投喂条件下BDE-209胁迫对褶皱臂尾轮虫滤水率的影响 Fig. 1 Effect of BDE-209 on the filtering rate(F)of B. plicatilis with mono-food feeding of microalga |
BDE-209胁迫对褶皱臂尾轮虫摄食率(G)的作用表现出与F类似的影响。在胁迫组中褶皱臂尾轮虫G随胁迫浓度的升高而不断下降,与对照组相比差异显著(P<0.01)。与F变化类似,24h胁迫组中的下降幅度要明显高于12h时的下降幅度,说明随着胁迫时间的延长BDE-209对褶皱臂尾轮虫摄食的抑制作用增强(图 2)。以0.6mg/L的高浓度组为例,BDE-209胁迫24h褶皱臂尾轮虫对小球藻、青岛大扁藻、等鞭金藻8701、小新月菱形藻的G分别下降至海水对照组的73.62%、67.40%、57.26%、78.33%,与对照组相比,BDE-209胁迫未改变投喂单一饵料条件下褶皱臂尾轮虫对各种饵料藻G的变化顺序,均表现为青岛大扁藻<小新月菱形藻<等鞭金藻8701<小球藻的变化。
 |
| 图 2 单一饵料微藻投喂条件下BDE-209胁迫对褶皱臂尾轮虫摄食率的影响 Fig. 2 Effect of BDE-209 on the feeding rate(G)of B. plicatilis with mono-food feeding of microalga |
投喂等生物量混合的饵料微藻时,无BDE-209胁迫的对照组中褶皱臂尾轮虫对混合饵料中各微藻滤水率F的大小顺序为: 小新月菱形藻<青岛大扁藻<小球藻; BDE-209胁迫条件下对混合饵料中各组分的滤水率影响与单一饵料结果一致,均随胁迫浓度的升高呈现出逐渐下降的趋势(P<0.05)(图 3),但F变化呈现出青岛大扁藻<小新月菱形藻<小球藻的变化趋势,与单一饵料投喂条件下的青岛大扁藻<小球藻<小新月菱形藻的变化顺序有所变化。
 |
| 图 3 BDE-209胁迫对褶皱臂尾轮虫对混合饵料中各种微藻滤水率的变化 Fig. 3 Effect of BDE-209 on the filtering rate(F)of B. plicatilis with mixed food feeding of microalga |
与相同条件下滤水率F的变化趋势类似,BDE- 209胁迫条件下褶皱臂尾轮虫对混合饵料中各种微藻摄食率G仍随胁迫浓度的升高呈现出不断下降的趋势,与对照组相比差异显著(P<0.05)(图 4)。受到BDE-209胁迫后,褶皱臂尾轮虫对混合饵料中各种微藻G大小顺序的影响与单一饵料微藻投喂结果一致,均未发生变化,为青岛大扁藻<小新月菱形藻<小球藻。
 |
| 图 4 BDE-209胁迫对褶皱臂尾轮虫对混合饵料中各种微藻摄食率的变化 Fig. 4 Effect of BDE-209 on the feeding rate(G)of B. plicatilis with mixed food feeding of microalga |
为了进一步阐明褶皱臂尾轮虫具有摄食选择性,我们对混合饵料投喂条件下褶皱臂尾轮虫的选食率(Sr)和选择性指数(Si)的变化进行研究。结果表明,如图 5所示,褶皱臂尾轮虫对混合饵料中微藻的选食率Sr随BDE-209胁迫浓度的升高呈现出逐渐下降的趋势,与对照组相比差异显著(P<0.05),且24h时Sr的下降程度明显高于12h时的影响。0.037mg/L组与对照组相比,BDE-209胁迫12h与24h时,褶皱臂尾轮虫对混合饵料中各微藻组分的Sr均有所下降,但未达到显著水平(P>0.05)。BDE-209胁迫条件下褶皱臂尾轮虫对混合饵料中各种饵料藻Sr的变化顺序与滤水率F的变化一致,均为青岛大扁藻<小新月菱形藻<小球藻,但与对照组中“小新月菱形藻<青岛大扁藻<小球藻”的变化顺序不同。研究结果进一步说明BDE-209胁迫能改变褶皱臂尾轮虫对饵料单胞藻的摄食选择性。
 |
| 图 5 BDE-209胁迫对褶皱臂尾轮虫对混合饵料中各种微藻选食率的变化 Fig. 5 Effect of BDE-209 on the food selectivity rate(Sr)of B. plicatilis with mixed food feeding of microalga |
褶皱臂尾轮虫对混合饵料的选择性指数与选食率的变化完全一致: 随着BDE-209胁迫浓度的升高,褶皱臂尾轮虫对小球藻的Si呈现不断增加的趋势,且总体呈现出青岛大扁藻<小新月菱形藻<小球藻的变化顺序,但不同于对照组中“小新月菱形藻<青岛大扁藻<小球藻”的变化(图 6)。研究结果说明BDE-209胁迫能改变褶皱臂尾轮虫正常的选食规律。
 |
| 图 6 BDE-209胁迫对褶皱臂尾轮虫对混合饵料中各种微藻选择性指数的变化 Fig. 6 Effect of BDE-209 on the selectivity index(Si)of B. plicatilis with mixed food feeding of microalgae |
BDE-209胁迫12h和24h条件下褶皱臂尾轮虫摄食混合微藻总生物量的变化随着胁迫浓度增加均呈现出不断下降的趋势(P<0.05)。以0.6mg/L的高浓度组为例,BDE-209胁迫12h和24h时褶皱臂尾轮虫摄食饵料藻总生物量分别下降至对照组的78.98%和73.54%,差异极显著(P<0.01),说明BDE-209既能改变褶皱臂尾轮虫的摄食选择性,又能抑制褶皱臂尾轮虫摄食饵料藻总生物量(图 7)。
 |
| 图 7 BDE-209胁迫对褶皱臂尾轮虫摄食混合饵料中各组分微藻总生物量的影响 Fig. 7 Feeding changes of B.plicatilis on microalgal biomass when fed with mixed bait microalgae |
已有研究证明,低溴代多溴联苯醚对海洋浮游生物有一定的危害,不但产生急性毒性作用,而且还会影响浮游动物的行为(Breitholtz et al,2008; 李卓娜等,2009)。那么高溴代多溴联苯醚是否也存在这种危害呢?本研究结果显示,BDE-209的急性毒性胁迫能有效抑制褶皱臂尾轮虫的运动,并具有一定的致死作用,但在实验的浓度范围内并未出现半数致死现象。该结果与Nakari等(2008)研究PBDEs的急性毒性结果相似,即在实验所能允许的最高浓度仍未达到对大型蚤的半数致死效应。可能原因在于BDE-209的分子量大,空间位阻大,难以全部通过生物膜进入浮游动物褶皱臂尾轮虫的体内,却会附着在轮虫表面而影响其生命活动。轮虫通过头冠纤毛运动,其运动能力受到激动神经肌肉的非限制性收缩和纤毛摆动的影响,因此我们分析PBDEs对轮虫运动能力的干扰可能因为以下两个方面:(1)PBDEs是一种神经毒性物质。少量进入轮虫体内的BDE-209可能通过干扰轮虫神经系统的正常功能降低纤毛运动的协调性而产生运动抑制;(2)大分子的BDE-209能够与海水中的微藻或碎屑结合成较大的颗粒粘合附在轮虫头冠部的纤毛上,干扰纤毛的摆动从而产生运动抑制作用。
浮游动物的摄食行为通常被认为是生物体受到环境干扰时的第一响应者,可以作为评价毒物对水生生物个体毒性的一个重要指标(Fernandezcasalderrey et al,1994)。在本研究中,BDE-209胁迫能显著抑制褶皱臂尾轮虫对四种海洋微藻的摄食作用,具有明显的浓度、时间依赖性,且滤水率下降程度要高于摄食率的变化。类似的研究结果在重金属以及有机磷农药对浮游动物的研究中均有报道(Ferrando et al,1993),说明浮游动物的摄食活动极易受到外源胁迫的影响,其中滤水率较摄食率的变化更敏感。这种敏感性的差异可能与轮虫的被动滤食方式有关(Kersting et al,1981)。由于轮虫只能从滤入的海水中获取食物,滤水的变化必然对摄食造成影响,因此与摄食率相比轮虫的滤水率更敏感。已经有研究表明鱼的行为变化可以作为检测毒物急性毒性较为快速而敏感的指标(de Paiva Magalhães et al,2007),但是在浮游动物中的应用较少。本研究结果说明浮游动物的滤水率可以作为灵敏指示外源胁迫的指标之一,但尚需开展进一步的研究以证明其有效性。
BDE-209胁迫不仅改变了褶皱臂尾轮虫的运动能力与摄食行为,而且改变其摄食选择性。从研究结果来看,在单一和混合饵料供给条件下,BDE-209胁迫均能改变褶皱臂尾轮虫对四种微藻滤水率的大小顺序,说明BDE-209胁迫影响褶皱臂尾轮虫摄食不同种类饵料微藻的选择性。同时,随着BDE-209胁迫时间的延长和暴露浓度的增加,褶皱臂尾轮虫对混合饵料中各种饵料单胞藻的选食率和摄食总生物量均呈现不同程度的下降,选择性指数呈现出不同的变化趋势,说明BDE-209可能影响褶皱臂尾轮虫的选食行为,其中无论有无BDE-209胁迫轮虫对混合饵料中小球藻的滤水率、摄食率、选食率和选择性指数均最大,这与小球藻的细胞结构有密切关系。小球藻个体最小,细胞壁纤维化程度较低,有利于褶皱臂尾轮虫幼虫的摄食与消化。小新月菱形藻的个体较小球藻大,且具有难消化的硅质外壳,而青岛大扁藻则由于个体较大而不宜为褶皱臂尾轮虫幼虫摄食。这种微藻结构与细胞组成上的差异与褶皱臂尾轮虫幼虫的摄食选择性密切相关。
4 结论(1) BDE-209的急性毒性作用能对褶皱臂尾轮虫产生致死作用,并有效抑制其运动能力;
(2) 在单一饵料培养时,BDE-209胁迫降低褶皱臂尾轮虫对各种饵料单胞藻的滤水率和摄食率;
(3) BDE-209胁迫改变褶皱臂尾轮虫对混合饵料的选食作用,其中滤水率的变化能在个体水平上灵敏指示褶皱臂尾轮虫对外源胁迫的响应;
(4) BDE-209胁迫与否,褶皱臂尾轮虫最适宜的饵料都是小球藻。
参考文献