 2016, Vol. 47
2016, Vol. 47

熊本牡蛎(Crassostrea sikamea)多嵴和无嵴品系子二代生长性状的选择效应

http://dx.doi.org/10.11693/hyhz20151100288
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 张跃环, 秦艳平, 张扬, 李军, 肖述, 向志明, 马海涛, 喻子牛. 2016.
- ZHANG Yue-Huan, QIN Yan-Ping, ZHANG Yang, LI Jun, XIAO Shu, XIANG Zhi-Ming, MA Hai-Tao, YU Zi-Niu. 2016.
- 熊本牡蛎(Crassostrea sikamea)多嵴和无嵴品系子二代生长性状的选择效应
- SELECTIVE EFFECT OF THE SECOND GENERATION FOR FAST GROWTH IN TWO STRAINS OF THE KUMAMOTO OYSTER CRASSOSTREA SIKAMEA
- 海洋与湖沼, 47(3): 581-586
- Oceanologia et Limnologia Sinica, 47(3): 581-586.
- http://dx.doi.org/10.11693/hyhz20151100288
-
文章历史
- 收稿日期:2015-11-19
- 改回日期:2016-03-09
引用本文
张跃环,
秦艳平,
张扬,
李军,
肖述,
向志明,
马海涛,
喻子牛. 2016. 熊本牡蛎(Crassostrea sikamea)多嵴和无嵴品系子二代生长性状的选择效应[J]. 海洋与湖沼, 47(3): 581-586
ZHANG Yue-Huan,
QIN Yan-Ping,
ZHANG Yang,
LI Jun,
XIAO Shu,
XIANG Zhi-Ming,
MA Hai-Tao,
YU Zi-Niu. 2016. SELECTIVE EFFECT OF THE SECOND GENERATION FOR FAST GROWTH IN TWO STRAINS OF THE KUMAMOTO OYSTER CRASSOSTREA SIKAMEA[J]. Oceanologia et Limnologia Sinica, 47(3): 581-586.
熊本牡蛎(Crassostrea sikamea)多嵴和无嵴品系子二代生长性状的选择效应
1.
中国科学院南海海洋研究所热带海洋生物资源与生态重点实验室 广东省应用海洋生物学重点实验室 广州 510301;
2. 南海生物资源开发与利用协同创新中心 广州 510275
2. 南海生物资源开发与利用协同创新中心 广州 510275
摘要:
为了进一步检查熊本牡蛎(Crassostrea sikamea)多嵴品系及无嵴品系的连续选择反应能力,以两个品系子一代上选组为材料,以壳高为指标,按照10%选择强度,开展了两个品系子二代混合上选研究,评估了两个品系子二代选择反应、现实遗传力,计算了两个品系的遗传改进量。结果表明:两个品系仍具有较高的现实遗传力,多嵴品系遗传参数均大于无嵴品系,表现出较好的遗传改良效果。多嵴和无嵴品系幼虫期的选择反应分别为0.47、0.34,稚贝期为0.65、0.40,养成期为0.82、0.56;幼虫期的现实遗传力分别为0.27、0.19,稚贝期为0.37、0.23,养成期为0.47、0.32;幼虫期的遗传改进量分别为4.83%、3.61%,稚贝期为7.55%、5.76%,养成期为9.95%、6.47%。经过一周年的养成,多嵴品系壳高显著大于无嵴品系,两个品系现实遗传力分别为0.50、0.35,遗传改进量分别为10.13%、6.61%。由此可见,多嵴品系具有较快的生长速度,遗传改良潜力较大。本研究为熊本牡蛎遗传改良和新品系培育奠定了坚实的基础。
关键词:
熊本牡蛎
连续选择
选择反应
现实遗传力
遗传改进量
SELECTIVE EFFECT OF THE SECOND GENERATION FOR FAST GROWTH IN TWO STRAINS OF THE KUMAMOTO OYSTER CRASSOSTREA SIKAMEA
1.
Key Laboratory of Tropical Marine Bio-resources and Ecology, Guangdong Provincial Key Laboratory of Applied Marine Biology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China;
2. South China Sea Bio-Resource Exploitation and Utilization Collaborative Innovation Center, Guangzhou 510275, China
2. South China Sea Bio-Resource Exploitation and Utilization Collaborative Innovation Center, Guangzhou 510275, China
Abstract:
To examine response to selection, realized heritability, and genetic gain for in shell height of the cultchless Kumamoto oyster Crassostrea sikamea taken from the second generation in southern China, the selective effect was evaluated using multi-ridge and non-ridge strains by selecting the largest 10% oyster from two parents for of selected groups, while the equal number of the parents were randomly chosen as the control groups before the removal of parents for truncation selection. Results showed that the genetic parameters of the multi-ridge strain was were larger than those of the non-ridge strain, while two strain still existed and both were in relatively high genetic level. For multi-ridge strain, the response to selection and realized heritability were 0.47, 0.27 for larvae, 0.65, 0.37 for spat, and 0.82, 0.47 for grow out adult stage, respectively. For non-ridge strain, response to selection and realized heritability, they were 0.34, 0.19 for larvae, 0.40, 0.23 for spat, and 0.56, 0.32 for adult grow out stage, respectively. Genetic gain of multi-ridge and non-ridge strains was 4.83%, 3.61% for larvae, 7.55%, 5.76% for spat, 9.95%, 6.47% for adult, respectively. At the end of 360 day, the realized heritability and current genetic gain was 0.50, 10.13% for multi-ridge strain 0.35, 6.61% for non-ridge strain, respectively. This study shows that the multi-ridge strain has a potential of rapid growth rate and genetic improvement potential, which also and providesd a solid base for new strain cultivation as a solid foundation.
Key words:
Crassostrea sikamea
successive selection
Selected reaction
realized heritability
genetic gain
对于贝类选择育种而言,连续几代的选择均可以获得积极的选择反应(张国范等,2009)。Haskin等(1979)对美洲牡蛎(Crassostrea virginica)进行了连续四代的抗单孢子虫病选择,使其抗病力比野生群体提高了8—9倍; Newkirk(1982)认为欧洲牡蛎(Ostrea edulis)生长选择每代提高10%—20%是合理的; Paynter等(1990)报道了养殖人员经过18代选择的美洲牡蛎,其生长速度显著快于野生群体; Nell等(1999)对悉尼岩牡蛎(Saccostrea commercialis)体重进行了连续2代选择,获得了17.8%的遗传改进量; Nell等(2006)进行悉尼 岩牡蛎三代的抗单孢子虫病(QX疾病,Queensland Unknown Disease)选育,获得了较强的抗病系; Li等(2011)和Wang等(2012)对长牡蛎(C. gigas)进行了连续6代的混合选择,获得了积极的改良效果,培育出“海大1号”长牡蛎新品种; Dégremont等(2015)对长牡蛎进行了连续4代的抗疱疹病毒选育,获得了抗性较强的牡蛎新品系,存活力提高了61.8%。
至于其它常见经济贝类,Zheng等(2004,2006)对海湾扇贝(Argopecten irradians)进行了连续4代的混合选择,发现基础群体对其遗传效果具有显著影响,培育出“中科红”海湾扇贝; Zheng等(2013)还对华贵栉孔扇贝(Chlamys nobilis)进行了橘红色闭壳肌品系连续4代的选育,培育出“南澳1号”华贵栉孔扇贝新品种; 闫喜武等(2010)和刘振等(2014)对菲律宾蛤仔(Ruditapes philippinarum)进行了连续8代混合选择,培育出“斑马蛤”菲律宾蛤仔新品种; You等(2009)对九孔鲍(Haliotis diversicolor)进行了连续4代混合选择,培育出“东优1号”杂色鲍新品种。
先前,我们对熊本牡蛎(C. sikamea)的多嵴、无嵴品系进行了一次上选,经过一周年的养成,获得了比较好的选择反应,并且获得了8.41%、7.71%遗传改进量(张跃环等,2016)。本研究是以两个品系子一代选择组为材料,继续观测两个品系的选择反应、现实遗传力,计算二次选择的遗传改进量,为熊本牡蛎遗传育种提供理论基础和实践经验。
1 材料与方法 1.1 亲本来源如图 1所示,本研究以2012年5月采集的湛江野生群体熊本牡蛎为材料,构建了核心基础群。于2013年5月,从核心基础群中筛选出多嵴和无嵴品系,
 |
| 图 1 两个品系熊本牡蛎子二代选择育种技术路线图 Fig. 1 Pedigree of selected strains indicating founder strain and selection intensity(i)for a successive two-generation selection |
按照10%留种率,进行了两个品系熊本牡蛎子一代选择反应及其现实遗传力研究。在此基础上,于2014年5月,以多嵴、无嵴品系熊本牡蛎上选的子一代作为材料,随机筛选每个品系若干个个体(剔除掉外部形态不整齐个体)作为育种材料进行子二代连续上选。
1.2 实验处理利用截头法分别对多嵴和无嵴品系子一代进行混合选择,留种率均为10%,选择强度(i)均为1.755。从两种类型中各随机抽取1200个个体,用游标卡尺进行测量(精确至0.02mm),绘制个体壳高大小的频率分布图(图 1)。从每个群体中选择10%的最大个体(120个),作为选择组2(selected group 2,SG2),在剩余的个体中随机抽取120个个体作为对照组2(control group2,CG2)。其中,多嵴品系亲本的截点为62.37mm,雌雄比例为87:33; 无嵴品系亲本的截点为57.46mm,雌雄比例为81:39(图 1)。
1.3 子代培育采用解剖方式获取配子,为了保证所有雄性个体的繁殖机会,每个实验组均采用单个雄性个体与所有卵子配对的原则进行(1♂:n♀)。实验中为了防止发生组间污染,利用300目筛绢网进行幼虫选育,各实验组均保留D型幼虫约1.2亿个,多余幼虫全部放掉; 每个实验组放置在3个10m3水泥池中培养,幼虫培养密度控制在4ind./mL。
幼虫培育期间,盐度控制在20—25; 稚贝培养的盐度控制在15—20。室内培育期间,6日龄以前,投喂云微藻(Chlorella sp.),之后混合投喂云微藻和牟氏角毛藻(Chaetoceros muelleri)(体积比=1:1),投饵量视摄食情况而定,日投喂量在(5—10)×104cell/mL。当幼虫出现鳃原基、眼点、足时,利用波纹板采苗。当稚贝壳高在10mm左右时装入5mm网孔塑料袋在竹林盐厂生态池中进行中间育成,每袋密度控制在240—300个。随着稚贝生长,定期调整密度,最后移至孔径10mm的扇贝笼中养成,密度控制在每层50—60个。
1.4 测定指标壳高小于400μm的幼虫在显微镜下用目微尺(10×)测量,壳长大于3.0mm的子代和亲本用游标卡尺测量,精确值0.02mm。每个实验组设3个重复,每个重复随机测量30个个体。
1.5 数据分析为了保证方差齐性,所有的壳高进行以10为底的对数转化。用SPSS19.0统计软件对数据进行分析处理,不同实验组间数据的比较采用双因素方差分析方法(Two-way ANOVA),用Turkey法进行组间多重比较,差异显著性设置为P<0.05。
参照Zheng等(2006)使用的方法,将两个品系熊本牡蛎子二代的选择反应(Selected reaction,SR)、现实遗传力(hR2)及遗传改进量(Genetic gain,GG)用以下公式计算
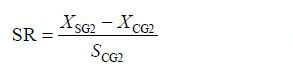 (1)
(1) (2)
(2)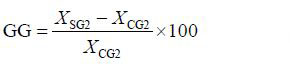 (3)
(3)公式(1)、(2)和(3)中: XSG2和XCG2分别是选择组(SG2)和对照组(CG2)个体的平均壳高,SCG2是对照组(CG2)的标准差,i是选择强度。
为了确定品系来源及其选择效应对生长性状的影响,参照Zhang等(2007)方法,利用双因子分析模型检测品系来源(strain origin,SO)及选择效应(selective effect,SE)对各实验组子代壳高的影响
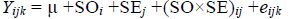
式中: Yijk为k个重复i个品系j种选择下的壳高(存活率); μ为常数; SOi为壳高的品系来源(i=1,2); SEj为壳高的选择效应(j=1,2);(SO×SE)ij为品系与选择效应的交互作用; eijk为随机误差(k=1,2,3)。
2 结果 2.1 壳高大小幼虫期,3日龄时,多嵴品系选择组壳高显著大于其它实验组(P<0.05); 9—15日龄期间,多嵴品系选择组壳高显著大于无嵴品系对照组,而多嵴品系对照组与无嵴品系无显著差异(P>0.05)。经过双因子分析模型分析,品系来源及其选择效应均对子二代幼虫壳高造成了显著影响,15日龄时二者间的交互作用也造成了显著影响(P<0.05)。稚贝期(30—60日龄),多嵴品系选择组壳高显著大于无嵴品系对照组,而其它两个实验组间无显著差异(P>0.05)。在此期间,选择效应对子二代壳高具有显著影响。养成期,90—180日龄期间,多嵴品系选择组壳高一直显著大于无嵴品系对照组(P<0.05); 360日龄时,多嵴品系选择组显著大于其它三个实验组,无嵴品系选择组显著大于相应对照组(P<0.05),表现出明显的选择效果。180—360日龄期间,品系来源及其选择效应均对子二代壳高具有显著影响(P<0.05),二者间交互作用尚未发生显著影响(表 1,2)。
表 1 两个品系子二代选择组和对照组壳高
Tab. 1 Shell height of the control and selective groups in different ages
| 牡蛎日龄 | 多嵴品系 | 无嵴品系 | ||
| CG2 | SG2 | CG2 | SG2 | |
| 幼虫期 | ||||
| 3 | 89.63±9.17b | 93.15±9.08a | 86.14±8.72b | 88.62±8.41b |
| 9 | 220.36±23.46ab | 230.57±22.98a | 212.88±23.73b | 220.65±21.65ab |
| 15 | 313.73±30.66ab | 332.29±30.87a | 296.77±31.16b | 309.54±29.14ab |
| 稚贝期 | ||||
| 30 | 2.17±0.24ab | 2.33±0.21a | 1.98±0.32b | 2.09±0.25ab |
| 45 | 5.64±0.59ab | 6.05±0.55a | 5.22±0.71b | 5.52±0.61ab |
| 60 | 12.36±1.66ab | 13.35±1.45a | 11.35±1.58b | 12.03±1.65ab |
| 养成期 | ||||
| 90 | 27.61±3.64ab | 30.31±3.27a | 25.66±3.31b | 27.29±2.87ab |
| 180 | 41.28±4.82ab | 45.38±4.62a | 36.77±4.17b | 39.14±3.96ab |
| 360 | 56.96±6.57b | 62.73±6.35a | 54.45±5.78c | 58.05±5.72b |
| 幼虫期壳高单位为μm,稚贝期和养成期壳高单位为mm; CG2、SG2分别表示对照组和选择组; 每一行不同字母表示彼此间差异显著(P<0.05) | ||||
表 2 品系来源(SO)及选择效应(SE)对其子二代生长性状影响的方差分析
Tab. 2 Analysis of variance showing the strain origin(SO)and selective effects(SE)on growth for each experimental group
| 类别 | 来源 | 自由度 | 均方 | P值 |
| SO | 1 | 0.011 | <0.001*** | |
| 第3天 | SE | 1 | 0.007 | <0.001*** |
| SO×SE | 1 | <0.001 | 0.410 | |
| SO | 1 | 0.009 | 0.002** | |
| 第9天 | SE | 1 | 0.008 | 0.004** |
| SO×SE | 1 | <0.001 | 0.686 | |
| SO | 1 | 0.039 | <0.001*** | |
| 第15天 | SE | 1 | 0.005 | 0.018* |
| SO×SE | 1 | 0.006 | 0.023* | |
| SO | 1 | 0.015 | 0.332 | |
| 第30天 | SE | 1 | 0.004 | 0.014* |
| SO×SE | 1 | 0.017 | 0.376 | |
| SO | 1 | 0.012 | 0.275 | |
| 第45天 | SE | 1 | 0.043 | 0.009** |
| SO×SE | 1 | <0.001 | 0.673 | |
| SO | 1 | 0.048 | 0.174 | |
| 第60天 | SE | 1 | 0.003 | 0.016* |
| SO×SE | 1 | 0.022 | 0.354 | |
| SO | 1 | <0.001 | 0.842 | |
| 第90天 | SE | 1 | 0.189 | <0.001*** |
| SO×SE | 1 | 0.032 | 0.065 | |
| SO | 1 | 0.090 | <0.001*** | |
| 第180天 | SE | 1 | 0.036 | 0.001** |
| SO×SE | 1 | 0.002 | 0.429 | |
| SO | 1 | 0.050 | <0.001*** | |
| 第360天 | SE | 1 | 0.016 | 0.001** |
| SO×SE | 1 | <0.001 | 0.896 | |
| *: P<0.05; **: P<0.01; ***: P<0.001 | ||||
从遗传参数上看,两个品系选择反应、现实遗传力均表现为幼虫期<稚贝期<养成期。多嵴、无嵴品系幼虫期的选择反应分别为0.47±0.12、0.34±0.06,稚贝期为0.65±0.05、0.40±0.05,养成期为0.82±0.07、0.56±0.07; 幼虫期的现实遗传力为0.27±0.07、0.19± 0.04,稚贝期为0.37±0.03、0.23±0.03,养成期为0.47± 0.04、0.32±0.04。从其遗传改进量上看,多嵴、无嵴品系品系幼虫期为4.83%±1.01%、3.61%±0.71%,稚贝期为7.55%±0.40%、5.76%±0.22%,养成期为9.95%±0.18%、6.47%±0.13%。经过一周年的养成,多嵴、无嵴品系的选择反应、现实遗传力和遗传改进量分别为0.88、0.62,0.50、0.35,10.13%、6.61%(表 3,图 2)。
表 3 两个品系子二代的选择反应(SR)、现实遗传力(hR2)及遗传改进量(GG)
Tab. 3 Response to selection(SR),realized heritability(hR2)and genetic gain(GG)of the two oyster strains
| 牡蛎日龄 | SR | hR2 | GG | |||
| MS | NS | MS | NS | MS | NS | |
| 幼虫期 | ||||||
| 3 | 0.38 | 0.28 | 0.22 | 0.16 | 3.93 | 2.88 |
| 9 | 0.44 | 0.33 | 0.25 | 0.19 | 4.63 | 3.65 |
| 15 | 0.61 | 0.41 | 0.34 | 0.23 | 5.92 | 4.30 |
| 稚贝期 | ||||||
| 30 | 0.67 | 0.34 | 0.38 | 0.20 | 7.37 | 5.56 |
| 45 | 0.69 | 0.42 | 0.40 | 0.24 | 7.27 | 5.75 |
| 60 | 0.60 | 0.43 | 0.34 | 0.25 | 8.01 | 5.99 |
| 养成期 | ||||||
| 90 | 0.74 | 0.49 | 0.42 | 0.28 | 9.78 | 6.35 |
| 180 | 0.85 | 0.57 | 0.48 | 0.32 | 9.93 | 6.45 |
| 360 | 0.88 | 0.62 | 0.50 | 0.35 | 10.13 | 6.61 |
| MS: 多嵴品系; NS: 无嵴品系 | ||||||
 |
| 图 2 两个品系不同阶段平均的选择反应(SR)、现实遗传力(hR2)及遗传改进量(GG) Fig. 2 Response to selection(SS),realized heritability(hR2)and genetic gain(GG)of the two oyster strains in different stages |
本研究中,通过设置亲本子代对照组来计算遗传参数,非用亲本本身,这主要是因为贝类的生长经常受到环境效应影响,如水质、饵料、水流等影响着贝类的生长性能(Zheng et al,2004,2006; 李冬春等,2013)。研究中发现,两个品系子一代的壳高就存在着显著差异,即多嵴品系子一代显著大于无嵴品系,且其选择反应均较高(张跃环等,2016),所以子二代还存在着一定程度的遗传改良效应。为此,我们进行了两个品系的子二代连续上选。
3.2 遗传改进量经过两代连续上选,两个品系遗传改进量存在着分歧,多嵴品系遗传改进量一直大于无嵴品系,尤其是到了360日龄时,多嵴品系和无嵴品系遗传改进量分别为10.13%、6.61%。这种分歧可能与品系内遗传多样性变化水平有关,也就是说多嵴品系内遗传多样性水平较高,而无嵴品系内遗传多样性水平较低(Li et al,2011; Wang et al,2012)。在贝类连续选择中,Newkirk等(1982)认为欧洲牡蛎生长选择每代提高10%—20%是合理的; Nell等(1999)对悉尼岩牡蛎体重进行了连续两代选择,获得了17.8%的遗传改进量; Zheng等(2006)对海湾扇贝进行了两代连续上选,发现新引进的海湾扇贝群体可以获得10%左右的遗传改进量; 张国范等(2010)对菲律宾蛤仔进行中日韩三个群体的连续两代的混合选择,获得了10%左右遗传改进量; 王庆志(2011)对长牡蛎进行了中日韩三个地理群体的三代混合选择,每代均获得了10%的遗传改进量。由此可见,本研究中子二代获得遗传改进量处于较低水平,这可能与熊本牡蛎自身个体较小,生长缓慢有着直接关系。
3.3 遗传参数从遗传参数看,在整个生活史中,多嵴品系的选择反应及现实遗传力均高于无嵴品系,尤其是经过周年养成的成体,二者间差异明显。我们分析,这可能是两个品系间的遗传多样性水平差异不同,多嵴品系中仍存在着较大的遗传方差,大的遗传方差暗示蕴藏着大的遗传力,遗传力大,遗传改进量相对较多,这也是多嵴品系遗传改进量较多的原因(Zheng et al,2004,2006; 马爱军等,2012)。此外,在熊本牡蛎野生群体中,多嵴类型占据大多数,无嵴类型相对较少,可能导致无嵴品系遗传多样性水平较低。
与其子一代选择相比,多嵴品系的现实遗传力较高,说明还具有较大的选择空间; 而无嵴品系的现实遗传力相对居中,较一代选择时结果略有下降,这可能是遗传多样性水平变化小,或者不存在较大的效应基因导致的(张国范等,2009; 张跃环等,2014)。经过双因子模型分析发现,两个品系经过一周年的养成,仍存在着显著的生长分歧,这也说明熊本牡蛎多嵴品系子二代具有较好的遗传改良效应,连续上选的效果是显著的。
4 结论本文发现经过连续两次的上选,两个品系生长性状仍具有较高的选择反应,且多嵴品系的遗传参数及遗传改进量均明显大于无嵴品系,出现了遗传分歧,这也说明多嵴品系仍具有较好的遗传改良前景。本文为熊本牡蛎遗传改良和新品种培育奠定了理论与实践基础。
参考文献