 2016, Vol. 47
2016, Vol. 47

小黄鱼(Pseudosciaena polyactis)形态性状与体质量的相关性及通径分析

http://dx.doi.org/10.11693/hyhz20151200294
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 刘峰, 陈琳, 楼宝, 詹炜, 陈睿毅, 徐冬冬, 王立改, 徐麒翔, 马涛, 毛国民. 2016.
- LIU Feng, CHEN Lin, LOU Bao, ZHAN Wei, CHEN Rui-Yi, XU Dong-Dong, WANG Li-Gai, XU Qi-Xiang, MA Tao, MAO Guo-Min. 2016.
- 小黄鱼(Pseudosciaena polyactis)形态性状与体质量的相关性及通径分析
- Correlation and path coefficient analysis ON body WEIGHT and morphometric traits of small yellow croaker Pseudosciaena polyactis
- 海洋与湖沼, 47(3): 655-662
- Oceanologia et Limnologia Sinica, 47(3): 655-662.
- http://dx.doi.org/10.11693/hyhz20151200294
-
文章历史
- 收稿日期:2015-12-03
- 改回日期:2016-01-31
引用本文
刘峰,
陈琳,
楼宝,
詹炜,
陈睿毅,
徐冬冬,
王立改,
徐麒翔,
马涛,
毛国民. 2016. 小黄鱼(Pseudosciaena polyactis)形态性状与体质量的相关性及通径分析[J]. 海洋与湖沼, 47(3): 655-662
LIU Feng,
CHEN Lin,
LOU Bao,
ZHAN Wei,
CHEN Rui-Yi,
XU Dong-Dong,
WANG Li-Gai,
XU Qi-Xiang,
MA Tao,
MAO Guo-Min. 2016. Correlation and path coefficient analysis ON body WEIGHT and morphometric traits of small yellow croaker Pseudosciaena polyactis[J]. Oceanologia et Limnologia Sinica, 47(3): 655-662.
小黄鱼(Pseudosciaena polyactis)形态性状与体质量的相关性及通径分析
1.
浙江省海洋水产研究所 舟山 316021;
2. 浙江海洋大学 舟山 316022;
3. 象山县水产技术推广站 宁波 315700;
4. 浙江省海水增养殖重点实验室 舟山 316021
2. 浙江海洋大学 舟山 316022;
3. 象山县水产技术推广站 宁波 315700;
4. 浙江省海水增养殖重点实验室 舟山 316021
摘要:
为了研究野生小黄鱼(Pseudosciaena polyactis)繁殖后代的形态性状对体质量的影响作用,以随机采集的729尾4.5月龄小黄鱼为研究对象,对每尾个体的体质量(g)和8个形态性状(cm):全长(X1)、体长(X2)、头长(X3)、躯干长(X4)、尾部长(X5)、尾柄长(X6)、尾柄高(X7)和体高(X8)进行准确测定,运用相关分析、多元回归分析和通径分析方法,分别计算性状间的相关系数、构建形态性状与体质量的多元回归方程,获得了通径系数和决定系数,进行了养殖小黄鱼的形态性状对体质量的影响研究。结果显示,8个形态性状与体质量的相关系数均达到极显著水平(P <0.01);通径分析中,4个形态性状对体质量的通径系数达到极显著水平(P<0.01),它们是影响体质量的重要指标,其中体长(通径系数:0.589)对体质量的直接影响最大。应用逐步多元回归分析建立了以体质量为因变量(Y),体长(X2)、躯干长(X4)、尾柄高(X7)和体高(X8)为自变量的回归方程:Y=-32.377 + 3.064X2-1.511X4+14.285X7+4.438X8。模型等式中形态性状与体质量的相关指数R2=0.927,说明所选性状是影响体质量的主要形态性状。以上分析结果为小黄鱼的选育测量指标的确定提供了理论依据。
关键词:
小黄鱼
养殖群体
形态性状
相关分析
通径分析
Correlation and path coefficient analysis ON body WEIGHT and morphometric traits of small yellow croaker Pseudosciaena polyactis
1.
Marine Fisheries Research Institute of Zhejiang, Zhoushan 316021, China;
2. Zhejiang Ocean University, Zhoushan 316022, China;
3. Xiangshan Aquaculture Technology Extension Service, Ningbo 315700, China;
4. Key Lab of Mariculture and Enhancement of Zhejiang Province, Zhoushan 316021, China
2. Zhejiang Ocean University, Zhoushan 316022, China;
3. Xiangshan Aquaculture Technology Extension Service, Ningbo 315700, China;
4. Key Lab of Mariculture and Enhancement of Zhejiang Province, Zhoushan 316021, China
Abstract:
To understand the relationship between body weight and morphometric traits of small yellow croaker Pseudosciaena polyactis, body weight (Y, g) and other eight morphometric traits (cm), e.g., total length (X1), body length (X2), head length (X3), trunk length (X4), tail length (X5), tail handle length (X6), tail handle height (X7), and body height (X8) were correlated on 729 individuals of 4.5-month-old P. polyactis. The correlation coefficient, path coefficient, and decision coefficient were calculated in correlation analysis, multiple regression analysis, and path analysis, and the effect of each morphometric trait on body weight was analyzed. The results show that all the correlation coefficients achieved highly significant levels (P<0.01). In addition, we established a regression equation using morphological traits as variables and body weight as dependent variable, in which direct and indirect path coefficients and determinant coefficients were calculated. Four morphometric traits achieved highly significant levels (P<0.01), and they are the main decisive morphometric traits on body weight. Among them body length was the most decisive variable (path coefficient = 0.589). The multiple linear regression model equation was established as Y = -32.377 + 3.064X2 - 1.511X4 + 14.285X7 + 4.438X8. The body weight and morphometric traits were highly correlated (R2=0.927). This research may provide a practical tool for the measurement of P. polyactis in selective breeding.
Key words:
Pseudosciaena polyactis
cultured stock
morphometric traits
correlation analysis
path analysis
小黄鱼(Pseudosciaena polyactis,Bleeker)又名小鲜、黄花鱼等,隶属于硬骨鱼纲Osteichthyes、石首鱼科Sciaendae、黄鱼属Larimichthys,是中国近海渔业的重要经济种类(李建生等,2009),其肉质鲜美、营养价值高,曾与大黄鱼(Larimichthys crocea)、墨鱼(Sepiella maindron)、带鱼(Trichiurus japonicas)并称为我国“四大海产”。小黄鱼为暖温性底层鱼类,具有规律性洄游习性(Jin et al,1996),3月份自越冬场开始生殖洄游,于4—5月在沿岸浅水区产卵(徐献明等,2012)。另外,小黄鱼属于广温、广盐性鱼类(谢立峰等,2006),但其进行产卵时对水温要求比较严格,而盐度条件则可小幅度波动(刘勇等,2007)。
小黄鱼作为我国四大海产之一,具有其特有的经济重要性,自20世纪70年代以来,由于过度捕捞、环境污染、气候变化等原因,小黄鱼产量逐年减少,Chen等(1997)在研究渤海和黄海的渔业资源分布时,发现小黄鱼资源已经从以前接近20%的渔获量锐减到2%。进入20世纪90年代以来,由于禁渔区、禁渔期和伏季休渔措施的实施,小黄鱼产量逐渐恢复上升(Froese et al,2014)。然而,产量增加的部分主要是当龄鱼(林龙山等,2004a),因此,虽然小黄鱼渔获量有所增加,但是其年龄组成序列缩短、性成熟提前、渔获小型化的资源衰退现象非常明显(林龙山等,2004b)。为了避免小黄鱼像大黄鱼一样出现野生资源几乎灭绝的现象,需要采取措施对小黄鱼群体进行有效的管理和保护(Wang et al,2015)。采取小黄鱼的人工繁殖和良种选育工作是恢复小黄鱼的渔业资源,满足人们日益增长的消费需求的重要手段。近年来,国内已经相继开展小黄鱼的人工繁育工作,试图通过增殖放流补充海洋中小黄鱼资源。谢立峰等(2006)于2003年开展了小黄鱼亲鱼培育技术的研究; 徐献明等(2012)于2012年对近海捕获的小黄鱼亲本采取人工授精法获得受精卵,开展小黄鱼的人工繁殖研究,随后又对近海捕获的小黄鱼幼鱼进行人工培育研究(徐献明等,2014),但是由于多方面原因,上述研究最终未能取得小黄鱼的规模化繁育突破。2014年,浙江省海洋水产研究所科研人员对野生小黄鱼进行驯化养殖,亲鱼越冬及强化培育后,于2015年开展小黄鱼亲鱼的人工催产授精、苗种培育等研究,成功获得小黄鱼苗种2.5万尾,实现了小黄鱼的人工繁育。作为“四大海产”中,第三种人工繁育取得成功的鱼类,小黄鱼人工繁殖技术的重大突破,对渔业资源的恢复与利用意义重大,将为下一步增殖放流和人工养殖提供新品种。
我国从20世纪50年代末就已开始海水鱼类人工繁殖研究,到目前为止有很多种鱼类获得成功,并已经培育出具有优良性状新品种(全国水产技术推广总站,2010,2014),如大黄鱼“闽优1号”、“东海1号”,大口黑鲈“优鲈1号”等,这些新品种均取得了较好的经济效益。相比于上述鱼类,小黄鱼的人工繁殖及良种培育研究还处于刚刚起步阶段。在鱼类生长性状的遗传育种过程中,具有直观性和可度量性的体质量是常用的目标性状(楼允东,1999; 张敏莹等,2010)。但是在实际生产中,对于体质量的准确测量往往存在一定难度,而如果是对与体质量存在一定相关性的形态性状进行测量,则较为准确快捷,因此有必要通过相关分析(Ezekiel,1941)方法衡定不同形态性状与体质量的关联程度,然后采用回归分析(Land,1969)和通径分析(Efroymson,1960)方法剖析各个形态性状对体质量的影响方式和作用大小,这一研究方法已经在虾类(刘小林等,2004; 王志铮等,2011; 栗志民等,2011)、蟹类(耿绪云等,2007)、贝类(Ahmed et al,2000; 王明华等,2014; 张嘉丽等,2014)及鱼类(王新安等,2008; 杨贵强等,2011)等水产动物中开展研究。
由于小黄鱼人工繁殖研究刚刚取得突破,许多研究工作尚未开展,小黄鱼形态性状与体质量之间关系的研究还未见报道。因此,本研究对繁殖成功的小黄鱼养殖群体进行体质量和形态性状的测量和统计分析,研究形态性状与体质量的相关性并进行通径分析,衡量各个形态性状与体质量的关联程度,并深入剖析各形态性状对体质量直接作用和间接作用,确定影响小黄鱼体质量的主要形态性状及其决定作用,以期发现与体质量相关性较高的形态性状,建立体质量增长的多元回归方程,为开展小黄鱼的选择育种提供参考资料、奠定理论基础。
1 材料与方法 1.1 实验材料以2015年于舟山市西轩渔业科技岛通过野生小黄鱼繁殖获得的后代群体为研究材料,待鱼苗生长至4.5月龄时,随机取样729尾。用游标卡尺准确测量每一尾实验鱼的8个形态性状,包括全长(X1,cm)、体长(X2,cm)、头长(X3,cm)、躯干长(X4,cm)、尾部长(X5,cm)、尾柄长(X6,cm)、尾柄高(X7,cm)、体高(X8,cm),精确至0.01 cm。用电子天平准确称量体质量(Y,g),精确至0.01 g。各个形态性状测量方法如图 1所示。
 |
| 图 1 小黄鱼形态性状测量部位示意图 Fig. 1 Sketch of mensuration part of morphometric traits for P. polyactis |
运用SPSS 17.0软件对形态性状和体质量进行描述性统计,统计参数包括各个性状的最小值、最大值、平均值、标准差、标准误、变异系数等。采用Pearson法对各性状进行表型相关分析; 在表性相关分析的基础上运用逐步引入-剔除法建立形态性状与体质量的多元线性回归方程,进行形态性状对体质量的通径分析,剖析各形态性状对体质量的直接作用(通径系数)和间接作用(间接通径系数),间接通径系数公式如下:

其中,rxixj为形态性状Xi和Xj之间的Pearson相关系数; Pxj为性状Xj对体质量的通径系数。
利用相关系数和通径系数,根据下列公式计算形态性状对体质量的决定系数:
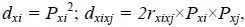
其中,dxi为形态性状Xi对体质量的决定系数; dxixj为某两个形态性状Xi和Xj对体质量的共同决定系数; rxixj为两个形态性状Xi和Xj之间的Pearson相关系数; Pxi为性状Xi对体质量的通径系数; Pxj为性状Xj对体质量的通径系数。
2 结果 2.1 表型数据描述性统计对所有形态性状和体质量数据进行描述性统计,结果列于表 1。
表 1 表型性状的描述性统计
Tab. 1 The descriptive statistics of phenotypic traits
| 性状 | 样本数 | 最小值 | 最大值 | 平均值 | 标准误 | 标准差 | 变异系数(%) |
| 全长(cm) | 729 | 7.55 | 15.99 | 12.19 | 0.04 | 1.18 | 9.68 |
| 体长(cm) | 729 | 6.32 | 13.05 | 10.08 | 0.04 | 1.01 | 10.02 |
| 头长(cm) | 729 | 1.92 | 4.00 | 2.84 | 0.01 | 0.29 | 10.21 |
| 躯干长(cm) | 729 | 1.81 | 6.44 | 3.55 | 0.02 | 0.51 | 14.37 |
| 尾部长(cm) | 729 | 2.28 | 4.85 | 3.69 | 0.02 | 0.42 | 11.38 |
| 尾柄长(cm) | 729 | 1.52 | 3.80 | 2.45 | 0.01 | 0.30 | 12.24 |
| 尾柄高(cm) | 729 | 0.44 | 1.15 | 0.82 | 0.01 | 0.10 | 12.20 |
| 体高(cm) | 729 | 1.89 | 3.86 | 2.85 | 0.01 | 0.31 | 10.88 |
| 体质量(g) | 729 | 4.23 | 36.63 | 17.42 | 0.19 | 5.26 | 30.20 |
从表 1中可以看出,所有性状中,体质量的变异系数最大,达30.20%; 8个形态性状的变异系数则相对较小,并且相互之间差异不是很大,分布在9.68%—14.37%之间,其中最大的为躯干长,最小的为全长。计算各个性状数据的标准误可以发现,所有性状的标准误均较小,表明本次随机采集的样本各参数对总体的估计可靠性较高。所有性状中,除了全长,其余性状测量数据变异系数均在10%以上,说明不同个体之间生长速度方面存在一定差异,这是进行选择育种的重要前提。
2.2 性状之间的相关性分析对小黄鱼的形态性状和体质量进行表型相关性分析,获得了所有性状之间的相关系数,结果列于表 2。从表中可以看出,各性状之间均表现为极显著水平的表型相关性(P<0.01),但是不同性状之间相关系数差别较大,在0.310—0.943范围内。形态性状相互间的相关性分析中,全长和体长的相关性最高,其相关系数为0.943; 而头长与躯干长的相关性最低,其相关系数仅有0.310。形态性状与体质量之间的相关性分析结果显示,8个形态性状中,全长与体质量相关性最高,其相关系数高达0.942,而躯干长与体质量之间的相关性则最低,其相关系数为0.684。
表 2 表型性状之间的相关性分析及显著性检验
Tab. 2 Correlation analyses and significance test on phenotypic traits
| 性状 | 体长 | 头长 | 躯干长 | 尾部长 | 尾柄长 | 尾柄高 | 体高 | 体质量 |
| 全长 | 0.943** | 0.717** | 0.718** | 0.894** | 0.848** | 0.875** | 0.868** | 0.942** |
| 体长 | 0.731** | 0.823** | 0.895** | 0.855** | 0.861** | 0.843** | 0.924** | |
| 头长 | 0.310** | 0.679** | 0.647** | 0.644** | 0.656** | 0.730** | ||
| 躯干长 | 0.555** | 0.612** | 0.667** | 0.623** | 0.684** | |||
| 尾部长 | 0.860** | 0.811** | 0.811** | 0.881** | ||||
| 尾柄长 | 0.744** | 0.767** | 0.826** | |||||
| 尾柄高 | 0.849** | 0.904** | ||||||
| 体高 | 0.898** | |||||||
| **表示变量之间存在极显著相关 | ||||||||
本研究在相关分析的基础上,采用多元回归的分析方法研究了形态性状对体质量的作用,同时进行自变量的共线性诊断,以免自变量之间存在严重共线性问题,影响多元回归模型的可靠性。检验结果显示全长性状存在严重共线性,所以将全长性状剔除后,对其余变量进行多元回归分析。表 3列出了采用逐步引入-剔除自变量的方法进行回归分析过程中各种参数的变化情况。表中数据显示,随着自变量的加入,回归方程R值在逐渐增加,从0.924增加到0.962,估计的标准误也由2.011逐渐降低为1.437,表明模型估计的准确性在不断增加,也就说明模型中自变量的增加对于更加准确地解释体质量具有重要作用。
表 3 模型汇总
Tab. 3 The model summary
| 模型 | 相关系数R | 决定系数R2 | 矫正决定系数 | 标准估计的误差 |
| 第1步 | 0.924a | 0.854 | 0.854 | 2.011 |
| 第2步 | 0.950b | 0.903 | 0.903 | 1.639 |
| 第3步 | 0.959c | 0.919 | 0.919 | 1.497 |
| 第4步 | 0.962d | 0.926 | 0.925 | 1.437 |
| a预测变量:(常量),体长; b预测变量:(常量),体长,体高; c预测变量:(常量),体长,体高,尾柄高; d预测变量:(常量),体长,体高,尾柄高,躯干长 | ||||
采用逐步法构建多元回归方程,获得了方程中每个自变量的偏回归系数(即非标准化回归系数),对各个自变量的偏回归系数进行显著性检验,检验结果列于表 4。
表 4 回归系数结果
Tab. 4 The calculation result of regression coefficient
| 变量 | 非标准化回归系数 (±SE) | 标准化回归系数 | 显著性 Sig. |
| (常量) | -32.377±0.555 | 0.000 | |
| 体长 | 3.064±0.153 | 0.589 | 0.000 |
| 体高 | 4.438±0.363 | 0.262 | 0.000 |
| 尾柄高 | 14.285±1.175 | 0.272 | 0.000 |
| 躯干长 | -1.511±0.191 | -0.145 | 0.000 |
| 因变量: 体质量 | |||
表 4中数据显示,对截距-32.377的检验结果为极显著(P<0.001),对各偏回归系数的显著性的检验结果为体长(X2)、躯干长(X4)、尾柄高(X7)和体高(X8)均达极显著水平(P<0.01),说明截距和这4个形态性状的偏回归系数与0之间均存在显著性差异,可认为所求的多元线性回归方程成立。采用F 检验的方法对构建的多元回归方程进行显著性检验(表 5),检验结果显示,回归方程达到极显著水平(F = 2254.455,P<0.01),说明该回归方程具有统计意义; 被纳入模型中的4个自变量对体质量的决定系数(R2)为0.926(表 3),说明模型中所纳入的4个形态性状对体质量有较大的决定作用。由此得出,以形态性状为自变量,体质量为因变量的多元回归方程可写为: Y = -32.377 + 3.064X2 - 1.511X4 + 14.285X7 + 4.438X8。
表 5 多元回归方程的方差分析
Tab. 5 Analysis of variance of multiple regression equations
| 指数 | 总平方和SS | 自由度df | 均方MS | F值 | Sig. |
| 回归 | 18629.368 | 4 | 4657.342 | 2254.455 | 0.000 |
| 残差 | 1495.668 | 724 | 2.066 | ||
| 总计 | 20125.036 | 728 |
本次研究共测量了8个形态性状,经共线性诊断剔除了共线性严重的全长及对体质量影响不显著的3个形态性状,剩余4个性状对体质量的影响达到极显著水平(P<0.01),所以回归方程中包含了该4个形态性状作为自变量。对回归方程中的偏回归系数进行标准化后可获得各性状对体质量的通径系数(表 4),检验结果表明,4个形态性状对体质量的通径系数(直接作用)均有统计学意义(P<0.05),但是不同形态性状对体质量的直接作用有所差异,最大高达0.589,最小为负值-0.145。直接作用由大到小排序依次为: 体长>尾柄高>体高>躯干长。由此可知,不同形态性状对体质量的影响作用存在很大差别,所以,采用通径分析的方法进行各个性状对体质量的直接和间接作用的深入剖析具有重要意义。
根据通径分析原理将形态性状对体质量的影响效应剖分为直接作用(通径系数)和间接作用(间接通径系数),结果列于表 6。通径系数反映自变量对依变量的直接作用,在所研究的目标性状中,体长对体质量的通径系数最大,即其对体质量的直接作用最大,尾柄高对体质量的直接作用次之,而躯干长对体质量的直接作用为负值,值为-0.145。
表 6 相关系数分解
Tab. 6 Decomposition of correlation coefficient
| 变量 | 与体质量的相关系数 | 通径系数 | 间接通径系数 | VIF | ||||
| 体长 | 躯干长 | 尾柄高 | 体高 | 总计 | ||||
| 体长 | 0.924 | 0.589 | — | -0.119 | 0.234 | 0.221 | 0.336 | 8.378 |
| 躯干长 | 0.684 | -0.145 | 0.485 | — | 0.181 | 0.163 | 0.829 | 4.480 |
| 尾柄高 | 0.904 | 0.272 | 0.507 | -0.097 | — | 0.222 | 0.632 | 4.864 |
| 体高 | 0.898 | 0.262 | 0.497 | -0.187 | 0.231 | — | 0.541 | 3.287 |
分析间接通径系数可以发现,小黄鱼的体长通过其余形态性状作用于体质量的间接通径系数各不相同,比较发现,所有间接作用中,尾柄高通过体长对体质量的间接作用最大(0.507)。结合性状之间的表型相关性分析得出,与体长相关性越高的形态性状,体长通过该性状对体质量起作用的间接通径系数越大,这一结果可以通过间接通径系数的计算公式进行解释。分析其它几个形态性状也可得出同样规律。另外还可发现,躯干长通过其它形态性状对体质量的间接作用均大于其自身的直接作用。由于躯干长对体质量是负影响作用,所以当其它性状通过其作用于体质量时均显示负影响,这一结果同样可以由间接通径系数计算公式进行解释。除了体长性状,其余三个性状通过其它性状对体质量的间接作用的总和均远远大于其自身对体质量的直接作用。其它性状通过体长作用于体质量的间接作用均大于通过其它形态性状的作用。表中各形态性状对体质量的方差膨胀因子(variance inflation factor,VIF)数值均小于经验值(VIF=10),表明各形态性状直接和间接作用的估计和回归模型的构建均无共线性的影响,因而本次分析结果是准确可靠的。
2.5 形态性状对体质量的决定程度各形态性状对体质量的决定系数见表 7。其中,对角线上为各形态性状对体质量的单独决定系数,对角线以上为两个性状对体质量的共同决定系数。比较发现,几个形态性状对体质量的单独决定系数差别较大。比较而言,体长的决定作用最大,其决定系数为0.347; 其次为尾柄高; 而躯干长的决定系数最小,仅为0.021。在两个性状共同决定系数中,体长和尾柄高对体质量的共同决定作用最大,决定系数为0.276。单独决定系数和两性状共同决定系数的总和等于总的决定系数,为0.927。根据总决定系数可计算出剩余因子e=0.270,这个值还比较大,说明对小黄鱼体质量有影响的自变量除了本次研究的几个形态性状之外,还有一些影响较大的因素没有考虑到,因此对小黄鱼体质量影响因素的全面分析还有待于进一步研究。
表 7 养殖小黄鱼形态性状对体重的决定系数
Tab. 7 The determinant coefficient of the morphometric traits on the body weight of P. polyactis
| 性状 | 体长 | 躯干长 | 尾柄高 | 体高 |
| 体长 | 0.347 | -0.141 | 0.276 | 0.260 |
| 躯干长 | 0.021 | -0.053 | -0.047 | |
| 尾柄高 | 0.074 | 0.121 | ||
| 体高 | 0.069 | |||
| 总决定系数 | 0.927 | |||
| 剩余项决定系数 | 0.073 | |||
| 剩余因子 | 0.270 | |||
在鱼类的选择育种中,生长性状的主要衡量指标为体质量,其次即为全长、体长等一些可测量的形态性状。本研究对小黄鱼的形态性状和体质量性状之间的关系进行探讨。首先对形态性状和体质量进行相关分析,确定形态性状与体质量之间的相关关系,结果显示,8个形态性状与体质量间的相关系数最小为0.684,均达到极显著水平(P<0.01),说明在小黄鱼的选择育种中,对形态性状进行选择,可以实现对体质量的间接选择。本研究中,与体质量相关性最高的前4个形态性状依次为: 全长>体长>尾柄高>体高。这一排序结果与刘贤德等(2010)对大黄鱼、Nguyen等(2010)和Trọng等(2013)对吉富罗非鱼形态性状与体质量相关性的研究结果存在一定差异。说明不同鱼类的形态性状与体质量之间的关系存在种群特异性。
3.2 通径分析必要性本次研究中,所有形态性状与体质量均为极显著相关(P<0.01),但是经过共线性检验发现全长存在严重共线性,所以需要将其剔除。对剩余7个形态性状与体质量进行了通径分析,结果表明,对体质量的直接影响仅4个形态性状达到极其显著水平,其余3个性状对体质量影响不显著(P>0.05)。由此可见,通过性状间的相关性分析只能得知形态性状与体质量之间总的关联性,无法区分形态性状对体质量的直接作用和通过其它形态性状对体质量的间接作用,因此,不能全面考察变量间的相互关系。多元回归分析在一定程度上能够消除变量之间的混淆,但由于偏回归系数带有单位,无法直接进行比较,需要对偏回归系数进行标准化处理,即进行通径分析。通径分析克服了相关分析与回归分析的不足,能够真实反映各自变量和依变量的关系(王新安等,2008)。通径分析可以获得各个形态性状的通径系数和间接通径系数,其中,通径系数表示形态性状对体质量直接作用的大小(耿绪云等,2007; 王明华等,2014),而间接通径系数则表示某个形态性状通过其它形态性状对体质量实现间接作用的大小(何小燕等,2009)。因此,采用通径分析的方法进行形态性状对体质量的直接作用和间接作用深入剖析,从中找出影响小黄鱼体质量的主要因素非常必要。
3.3 回归分析和通径分析剔除共线性严重的全长后,以7个形态性状作为自变量,体质量为因变量通过逐步引入-剔除的方法对形态性状和体质量进行多元回归分析,结果显示,体长、躯干长、尾柄高及体高与体质量的标准化回归系数极显著,而不显著的头长、尾部长和尾柄长性状则被剔除,以偏回归系数(非标准化回归系数)显著的4个形态性状为自变量,体质量为因变量构建多元回归方程,准确描述了形态性状与体质量之间的真实关系。回归方程中包含的形态性状对体质量总决定系数高于主要决定因子临界点0.85(王新安等,2011),说明所筛选的形态性状能在很大程度上反映体质量增长量的变异; 已剔除的性状对体质量增长变异影响不大,保留下来的形态性状确定为体质量增长的主要因素。但是剩余因子计算结果显示,除了本次研究涉及的形态性状,还存在着其它对体质量有影响的性状未找到,因此,需要增加测量性状的数目,使得对小黄鱼体型描述更加详细; 还需要考虑多个性状间的比例关系,建立一个更完善的模型,以此提高形态性状与体质量的相关性,这些都需要今后进一步深入的研究。
构建的多元回归方程经检验达到极显著水平(P<0.01),说明回归方程有意义,可见对于小黄鱼而言,形态性状作为自变量对体质量进行预测和估计是恰当的,这一结果在大黄鱼(刘贤德等,2008,2010)、大口黑鲈(何小燕等,2009)、罗非鱼(董在杰等,2010; 肖炜等,2015)等其它鲈形目及半滑舌鳎(刘峰等,2015)、牙鲆(王凯等,2008)、大菱鲆(王新安等,2008)等鲽形目鱼类中均已得到证实。因此,通过形态性状进行小黄鱼体质量方面的选育是可行的。近年来在多种鱼选育中开展了结合形态性状的选育研究。如,刘贤德等(2010)研究了13月龄和20月龄大黄鱼形态性状对体质量的影响,结果显示,这两个生长时期,均是体高对体质量的直接作用最大(0.522,0.394); 王新安等(2013)研究红鳍东方魨发现形态性状中体周长对体质量的直接影响(0.533)最大; 而本次研究中,养殖小黄鱼形态性状中对体质量直接作用最大的是体长(通径系数: 0.589),与上述报道有所不同,并且通径系数大于上述报道。
4 结论本文对729尾养殖小黄鱼的形态性状与体质量之间的关系进行了研究,包括形态性状与体质量的相关性分析、回归分析及通径分析。结果显示,8个形态性状与体质量均表现为极显著相关性,但是不同性状与体质量的相关性差别较大,其中,最高的性状是全长; 回归分析结果显示,全长存在严重的共线性,所以将其剔除后分析得出,有4个形态性状均与体质量存在显著的线性关系,以其为自变量,体质量为因变量构建了多元回归方程: Y = -32.377 + 3.064 X2 - 1.511 X4 + 14.285 X7 + 4.438 X8; 经过通径分析获得各形态性状对体质量的直接作用和间接作用,其中体长对体质量直接作用最大(0.589); 尾柄高通过体长对体质量的间接作用最大(0.507)。找出对体质量影响最大的几个形态性状,有助于实现对体质量的间接选择,获得正确的亲本,这对于小黄鱼的资源保护和恢复,以及开展规模化的人工养殖及优良品种选育均有重要意义。
参考文献
| 王凯, 刘海金, 刘永新, 等. 2008. 牙鲆形态性状对体重的影响效果分析. 上海海洋大学学报, 17(6): 655–660 |
| 王志铮, 吴一挺, 杨磊, 等. 2011. 日本沼虾(Macrobrachium nipponensis)形态性状对体重的影响效应. 海洋与湖沼, 42(4): 612–618 |
| 王明华, 钟立强, 蔡永祥, 等. 2014. 黄颡鱼形态性状对体重的影响效果分析. 浙江海洋学院学报: 自然科学版, 33(1): 41–46 |
| 王新安, 马爱军, 庄志猛, 等. 2013. 红鳍东方鲀(Takifugu rubripes)形态性状对体重的影响效果. 海洋与湖沼, 44(1): 135–140 |
| 王新安, 马爱军, 许可, 等. 2008. 大菱鲆幼鱼表型形态性状与体重之间的关系. 动物学报, 54(3): 540–545 |
| 王新安, 马爱军, 黄智慧, 等. 2011. 大菱鲆(Scophthalmus maximus)大规模家系选育优良家系的生长发育规律. 海洋与湖沼, 42(2): 266–273 |
| 刘勇, 严利平, 程家骅. 2007. 2003年东海北部和黄海南部外海小黄鱼产卵群体的分布特征及其与水温、盐度的关系. 中国水产科学, 14(7): 89–96 |
| 刘峰, 陈松林, 刘肖峰, 等. 2015. 半滑舌鳎3个形态性状与体质量的相关及通径分析. 海洋学报, 37(4): 94–102 |
| 刘小林, 吴长功, 张志怀, 等. 2004. 凡纳对虾形态性状对体重的影响效果分析. 生态学报, 24(4): 857–862 |
| 刘贤德, 蔡明夷, 王志勇, 等. 2008. 闽-粤东族大黄鱼生长性状的相关与通径分析. 中国海洋大学学报, 38(6): 916–920 |
| 刘贤德, 蔡明夷, 王志勇, 等. 2010. 不同生长时期大黄鱼形态性状与体重的相关性分析. 热带海洋学报, 29(5): 159–163 |
| 李建生, 林龙山, 程家骅. 2009. 东海北部秋季小黄鱼分布特征及其与底层温度和盐度的关系. 中国水产科学, 16(3): 348–356 |
| 杨贵强, 徐绍刚, 王跃智, 等. 2011. 硬头鳟幼鱼部分形态性状和体重的关系. 动物学杂志, 46(1): 16–22 |
| 肖炜, 李大宇, 邹芝英, 等. 2015. 湘湖品系尼罗罗非鱼形态性状对体重的影响. 西南农业学报, 28(1): 433–438 |
| 何小燕, 刘小林, 白俊杰, 等. 2009. 大口黑鲈形态性状对体重的影响效果分析. 水产学报, 33(4): 597–603 |
| 张敏莹, 刘凯, 段金荣, 等. 2010. 太湖秀丽白虾形态性状对体重影响的通径分析. 中国农学通报, 26(21): 417–421 |
| 张嘉丽, 王庆恒, 邓岳文, 等. 2014. 斧文蛤(Meretrix lamarchii)形态性状对体质量的影响效果分析. 渔业科学进展, 35(6): 110–113 |
| 林龙山, 程家骅. 2004a. 东海区小黄鱼渔业生物学现状的分析. 中国海洋大学学报, 34(4): 565–570 |
| 林龙山, 程家骅, 任一平, 等. 2004b. 东海区小黄鱼种群生物学特征的分析. 中国水产科学, 11(4): 333–338 |
| 耿绪云, 王雪惠, 孙金生, 等. 2007. 中华绒螯蟹(Eriocheir sinensis)一龄幼蟹外部形态性状对体重的影响效果分析. 海洋与湖沼, 38(1): 49–54 |
| 栗志民, 刘志刚, 王辉, 等. 2011. 企鹅珍珠贝(Pteria penguin)主要经济性状对体重的影响效果分析. 海洋与湖沼, 42(6): 798–803 |
| 徐献明, 张志勇, 吴建平, 等. 2012. 小黄鱼人工繁育试验. 水产养殖, 33(12): 12–13 |
| 徐献明, 张志勇, 曹昆. 2014. 小黄鱼幼鱼人工培育技术. 水产养殖, 35(8): 22–24 |
| 董在杰, 梁政远, 明俊超, 等. 2010. 尼罗罗非鱼体重与体维可量性状之间的关系. 广东海洋大学学报, 30(1): 32–38 |
| 谢立峰, 付贤康. 2006. 小黄鱼亲体培育技术. 中国水产, 50–51 |
| 楼允东. 1999. 鱼类育种学. 北京: 中国农业出版社, 3 |
| Ahmed M, Abbas G. 2000. Growth parameters of the finfish and shellfish juvenile in the tidal waters of Bhanbhore, Korangi Creek and Miani Hor Lagoon. Pakistan Journal of Zoology, 32(1): 21–26 |
| Chen D G, Liu Q, Zeng X Q, et al. 1997. Catch composition and seasonal variation of setnet fisheries in the Yellow and Bohai Seas. Fisheries Research, 32(1): 61–68 |
| Froese R, Pauly D. 2014. FishBase: world wide web electronic publication. http://www.fishbase.org |
| Jin X S, Tang Q S. 1996. Changes in fish species diversity and dominant species composition in the Yellow Sea. Fisheries Research, 26(3-4): 337–352 |
| Land K C. 1969. Principles of path analysis. Sociological Methodology, 1 3–37 |
| Nguyen N H, Ponzoni R W, Abu-Bakar K R, et al. 2010. Correlated response in fillet weight and yield to selection for increased harvest weight in genetically improved farmed tilapia (GIFT strain), Oreochromis niloticus. Aquaculture, 305(1-4): 1–5 |
| Trọng T Q, Han A M, Arendonk J A M V, et al. 2013. Heritability and genotype by environment interaction estimates for harvest weight, growth rate, and shape of Nile tilapia (Oreochromis niloticus) grown in river cage and VAC in Vietnam. Aquaculture, 384-387 119–127 |
| Wang L, Liu S F, Zhuang Z M, et al. 2015. Mixed-stock analysis of small yellow croaker Larimichthys polyactis providing implications for stock conservation and management. Fisheries Research, 161 86–92 |