 2016, Vol. 47
2016, Vol. 47

2011年胶州湾网采浮游植物群落结构及其环境影响因子

http://dx.doi.org/10.11693/hyhz20160300050
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 罗璇, 孙晓霞, 郑珊, 赵永芳. 2016.
- LUO Xuan, SUN Xiao-Xia, ZHENG Shan, ZHAO Yong-Fang. 2016.
- 2011年胶州湾网采浮游植物群落结构及其环境影响因子
- ON PHYTOPLANKTON COMMUNITY STRUCTURE AND CAUSATIVE FACTORS IN THE JIAOZHOU BAY IN 2011
- 海洋与湖沼, 47(5): 915-923
- Oceanologia et Limnologia Sinica, 47(5): 915-923.
- http://dx.doi.org/10.11693/hyhz20160300050
-
文章历史
- 收稿日期:2016-03-10
- 收修改稿日期:2016-05-16
引用本文
罗璇, 孙晓霞, 郑珊, 赵永芳 . 2016. 2011年胶州湾网采浮游植物群落结构及其环境影响因子[J]. 海洋与湖沼, 47(5): 915-923
LUO Xuan, SUN Xiao-Xia, ZHENG Shan, ZHAO Yong-Fang . 2016. ON PHYTOPLANKTON COMMUNITY STRUCTURE AND CAUSATIVE FACTORS IN THE JIAOZHOU BAY IN 2011[J]. Oceanologia et Limnologia Sinica, 47(5): 915-923.
2011年胶州湾网采浮游植物群落结构及其环境影响因子
1.
山东胶州湾海洋生态系统国家野外科学观测研究站 青岛 266071;
2. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室 青岛 266071;
3. 中国科学院大学 北京 100049
2. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室 青岛 266071;
3. 中国科学院大学 北京 100049
摘要:
本文于2011年1-12月对胶州湾海域浮游植物进行了周年调查,获得了同步的温盐和营养盐数据,分析了浮游植物丰度、优势种类、多样性指数和季节变化特征及其与环境影响因子的关系。结果表明:共检出浮游植物108种(包括变种和变型),其中硅藻门34属93种,甲藻门6属14种,金藻门1属1种。浮游植物主要由硅藻和甲藻两大类组成,其中硅藻占绝对优势,全年甲藻/硅藻比均小于0.08。优势种共有35种,夏、冬季优势种交替明显,全年几乎均出现的种类为圆筛藻(Coscinodiscus spp.),旋链角毛藻(Chaetoceros curvisetus)和中肋骨条藻(Skeletonema costatum)。生态类型主要以广布种、广温近岸种、温带近岸种为主,也出现了少数暖水种和外洋种。浮游植物丰度分别在2月、4月和7月份达到峰值,其中4月全年最高,为591×106cell/m3,2月是次高峰,数量为468×106cell/m3,7月为172×106cell/m3,秋季浮游植物丰度相对较低。本研究结果与胶州湾浮游植物典型双峰型变化不同。浮游植物群落多样性指数平均为2.4,秋、冬季节较高,春、夏季节偏低,水平分布上呈现湾外>湾口>湾内的趋势,均匀度平均值为0.26,分布趋势与多样性指数基本相同。由此可见,湾外浮游植物群落结构较湾口和湾内更为稳定,温度、硅酸盐、亚硝酸盐、硝酸盐、铵盐和磷酸盐对浮游植物丰度和群落结构存在明显影响。
关键词:
胶州湾
浮游植物
群落结构
周年变化
环境因子
ON PHYTOPLANKTON COMMUNITY STRUCTURE AND CAUSATIVE FACTORS IN THE JIAOZHOU BAY IN 2011
1.
Jiaozhou Bay Marine Ecosystem Research Station, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China;
2. Laboratory for Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
2. Laboratory for Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
Abstract:
An annual investigation on phytoplankton community was carried out in Jiaozhou Bay, Qingdao, East China, from January to December in 2011. Environmental data included temperature, salinity, and nutrients. The abundance, dominant species, diversity index, and dinoflagellate/diatom ratio were calculated. Total 108 species and varieties of phytoplankton were identified, of which 93 in 34 genera were Bacillariophyta, 14 species in 6 genera were Pyrrophyta, and 1 species in 1 genus was Chrysophyta. The phytoplankton community consisted mainly of diatom and dinoflagellate, and dominated by diatom. The annual average dinoflagellate/diatom ratio was less than 0.08. There were 35 dominant species, of which the following species spread widely in all seasons:Coscinodiscus spp., Chaetoceros curvisetus, and Skeletonema costatum. The ecological types were mainly eurytopic species, neritic temperate species, and a few warm water species and oceanic species in different seasons. The dominant species alternated obviously in winter and summer. The cell number of phytoplankton peaked in February, April, and July, at 591×106, 468×106, and 172×106cell/m3, respectively, and was relatively low in the autumn. This result was different from typical two-peak pattern reported previously. The average diversity index (H') was 2.4, relatively higher in autumn and winter. The value of H' was the highest outside of the bay, followed by that at the mouth, and then inside of the bay, the least. The average evenness (J) of the phytoplankton community was 0.26, and its horizontal distribution was similar to that of H', which means that the structure of phytoplankton community in the outside of Jiaozhou Bay was more stable than that at the mouth and inside the bay. Temperature, SiO32--Si, NO2--N, NO3--N, NH4+-N, and P were the main environmental factors affecting the abundance and community structure of phytoplankton.
Key words:
Jiaozhou Bay
phytoplankton
community structure
annual variation
environmental factors
位于食物链金字塔底端的浮游植物是海洋生态系统中最重要的初级生产者。研究海域的浮游植物数量和种类组成不仅有利于了解该海域生态系统特征, 对于监测海洋生态系统健康状况和海洋生物资源的可持续发展也具有重要意义。浮游植物的生长繁殖除了受到自身生物学特性的影响外, 还受到海流、扰动、温度、盐度、营养盐和其它生物等因素的影响(Lalli et al, 1993)。
胶州湾位于山东半岛南部, 海湾面积397km2, 湾内与外海的水体交换情况良好。由于受到自然变化和人类活动的双重影响, 胶州湾是沿海典型的半封闭模式海湾(齐雨藻等, 2004; 孙松等, 2005), 同时也是我国近海生态系统的缩影。长期并同步研究胶州湾浮游植物变化规律, 将是研究我国近海生态系统结构与功能变化以及变动趋势的基础。针对胶州湾的浮游植物特征, 国内许多学者开展过季节性、地域性的研究(钱树本等, 1983; 刘东艳等, 2002; 齐雨藻等, 2004; 吴玉霖等, 2004, 2005; Liu et al, 2004; 李艳等, 2005; 李广玉等, 2005; 孙晓霞等, 2011; 郑珊等, 2014; 杨世民等, 2014, 2015)。通过比对相关历史数据发现, 近二十年来, 由于海水中营养盐浓度和结构的变化, 胶州湾浮游植物种类和数量都发生了很大改变, 总量和中肋骨条藻(Skeletonema costatum)、角毛藻(Chaetoceros spp.)等小型链状硅藻数量呈现逐渐增加的趋势, 优势种组成也发生明显改变, 波状石鼓藻(Lithodesmium undulates)等暖水性种类的数量持续升高, 甲藻类浮游植物数量升高、分布范围扩大(孙晓霞等, 2011)。由此可见, 胶州湾海域的浮游植物群落以及整个生态系统状况正处于变化之中, 对该海域浮游植物群落应该开展持续的定点同步监测。从20世纪90年代开始, 胶州湾海洋生态系统研究站开始进行每年四个季度月的长期调查, 积累了大量的宝贵历史数据。
本研究工作在胶州湾海洋生态系统研究站的相关研究基础上, 对2011年胶州湾浮游植物开展了连续性周年调查, 结合之前报道过的数据, 对胶州湾浮游植物种类和数量的变化进行分析, 并在比较主要环境因子和相关调查指标相关性的基础上初步探讨了影响浮游植物群落结构的主要环境因素, 为了解胶州湾生态系统的现状, 揭示全球变化影响下近海生态系统的演变规律提供科学依据。
1 材料与方法 1.1 调查站位本研究在胶州湾内外海域共设12个调查站位(图 1)。调查时间为2011年1—12月, 调查频率为每月中旬1次。
 |
| 图 1 胶州湾调查区域站位图 Fig. 1 Sampling stations in the Jiaozhou Bay |
浮游植物取样按照《海洋调查规范》(国家技术监督局, 1992), 采样工具为浅水Ⅲ型浮游生物网(网口直径37cm, 网口面积为0.1m2, 网身长140cm, 网目76μm), 于各站位自水体底层至表层垂直拖网1次。样品采集后, 立即用5%甲醛固定保存, 带回实验室静置沉降24小时后移出上清液, 浓缩至100—150mL。利用0.1mL浮游植物计数框于Olympus BX51显微镜下进行浮游植物种类鉴定和计数, 对浮游植物种类的鉴定和命名参照相关资料(金德祥等, 1965; Tomas, 1997; 孙军等, 2002; 郭玉洁等, 2003; 杨世民等, 2006)进行。
浮游植物优势度计算公式: Y=(ni/N)×fi
浮游植物多样性H'采用香农-威纳指数计算公式(Shannon et al, 1949):
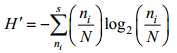
其中ni为第i种的细胞个数, N为所采集样品中所有种类的总个体数, fi为该种在各站中出现的频率。
浮游植物均匀度指数采用Pielou(1969)的计算公式:
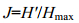
其中H′为前式计算的多样性指数值, Hmax为log2S, 表示多样性指数的最大值, S为样品总种类数。J值范围在0—1之间, J值大时, 体现种间个体数分布较均匀; J值小时, 反映中间个体数分布欠均。由于污染环境的种间个体数分布差别大, 即J是低值。
2 结果与讨论 2.1 胶州湾浮游植物丰度的时空分布格局 2.1.1 水平分布从图 2可以看出, 2011年胶州湾浮游植物丰度(×103cell/m3)的平面分布因月份而异。1月份浮游植物丰度湾内和湾外差异较小, 湾外略高于湾内, 呈现从湾内向湾外, 从北部向南部逐渐增多的趋势。2—6月份胶州湾浮游植物分布均呈现湾内大于湾口和湾外的分布格局, 但在不同月份其最大峰值出现的海域不同, 2—5月份, 浮游植物的高值区主要集中在湾内东部, 而6月份则出现在湾内西北部。7月份, 胶州湾浮游植物的分布格局与之前有明显的差异, 高值区出现在湾口处, 湾内西部区域次之。而8—11月份胶州湾浮游植物的分布又呈现湾内大于湾口和湾外的格局, 高值区均出现在湾内西部区域。12月份湾外浮游植物丰度高于湾内, 湾内高值区主要集中在湾中部。综上所述, 除了冬季1月和12月期间湾外浮游植物丰度高于湾内, 以及夏季7月份湾口处浮游植物丰度最高以外, 其余月份胶州湾浮游植物丰度的水平分布均表现为湾内 > 湾口 > 湾外。这种分布模式和2010年大部分月份的调查结果相似, 但是2010年1月和7月并没有表现出高值区转移的现象(郑珊等, 2014), 其原因还有待进一步研究。
 |
| 图 2 胶州湾浮游植物丰度周年水平分布(×103cell/m3)平面分布的周年变化 Fig. 2 Annual variation in horizontal distribution of phytoplankton cell abundance (×103cell/m3) in the Jiaozhou Bay |
浮游植物丰度年平均为1.24× 108cell/m3。由图 3可见, 2011年全年的浮游植物总丰度数量呈现三峰分布, 浮游植物丰度最高峰值出现在春季的4月份, 达到5.92×108cell/m3, 次高峰值出现在冬季的2月份, 达到4.68×108cell/m3, 第三高峰值出现在夏季的7月份, 为1.72×108cell/m3, 8—12月份浮游植物丰度维持在相对较低的水平, 12月份浮游植物丰度达到全年最低值, 仅为2.73×106cell/m3。钱树本等(1983)的周年调查表明, 1977年2月至1978年1月期间胶州湾浮游植物丰度的最高值出现在8月, 达109cell/m3, 次高峰出现在2月, 达108cell/m3; 刘东艳等(2002)报道1995—1996年胶州湾北部的浮游植物细胞密度最高峰值出现在1996年1月, 达1.4×107cell/m3, 次高峰出现在1995年9月, 达8.07×106cell/m3; 陈碧鹃等(2000)研究发现1998年胶州湾北部的浮游植物细胞密度最高峰值出现在9月, 为1.3×108cell/m3, 次高峰值出现在4月, 为1.5× 107cell/m3; 李艳等(2005)的调查显示, 2003年胶州湾浮游植物的最高峰值出现在9月, 为6.2×106cell/m3, 次高峰值出现在2月, 为5.5× 106cell/m3; 郑珊等(2014)对2010年胶州湾海域相同调查站位的浮游植物丰度进行了研究, 发现浮游植物丰度年平均为337.5×104cell/m3, 年度最高值出现在2月份, 为1.1× 107cell/m3, 次高峰出现在1月份, 为9.1×106cell/m3, 并且7—12月份浮游植物丰度水平相对较低。对比历史资料发现, 2011年胶州湾浮游植物丰度的最高值比1978年的低一个数量级, 比1995—1996年的结果高一个数量级, 和1998年的调查结果相当, 比2003年的高两个数量级, 比2010年高一个数量级。由此可见, 从20世纪70年代开始, 胶州湾浮游植物丰度峰值呈现先下降, 后上升, 再下降, 再升高的波动趋势。孙晓霞等(2011)研究了近30年来胶州湾网采浮游植物数量的长期变化, 也认为其总体呈现先下降后上升的趋势。此外, 以往研究认为胶州湾浮游植物丰度的周年变化存在“双周期”现象, 上述历史调查资料中的浮游植物高峰值出现在1月、2月和9月, 而次高峰值出现在1月、2月和4月, 不同调查年份对浮游植物峰值贡献最大的藻类各不相同。本研究中“双周期”现象不明显, 而是出现了三个峰值, 分别出现在4月、2月和7月; 4月份对峰值贡献最大的是一场中肋骨条藻赤潮, 2月份对次高峰值贡献最大的主要是柔弱根管藻(Rhizosolenia delicatula), 而7月份主要是旋链角毛藻(Chaetoceros curvisetus)。这种现象可能是由于胶州湾的水温、营养盐和摄食作用等因素综合导致某些种类暴发性增殖的结果(孙晓霞等, 2011)。
 |
| 图 3 胶州湾浮游植物总丰度的季节变化 Fig. 3 Seasonal variation in phytoplankton abundance in the Jiaozhou Bay |
从图 4可以看出, 胶州湾浮游植物总丰度和硅藻丰度的季节性变化趋势几乎一致, 说明在胶州湾硅藻占有绝对优势地位。甲藻的丰度相对较低, 呈现双周期动态变化, 分别在冬季的2月和夏季的7月达到峰值, 分别为1.44×106cell/m3和1.35×106cell/m3, 与2010年观察到10月份前甲藻丰度呈现逐渐上升趋势并在10月份达到最大值的现象有很大差异。经鉴定, 发现造成2月份出现丰度峰值的甲藻主要是纺锤角藻(Ceratium fusus), 其次是夜光藻(Noctiluca scintillancs), 而造成7月份峰值的主要是粗刺角藻(Ceratium horridum)。而甲藻/硅藻比全年都维持在较低水平, 平均为0.012, 分别在3月和8月出现两个峰值, 其中3月最高, 为0.07。从2010年甲藻/硅藻比最高值能达到1.147的结果可以看出, 2011年胶州湾浮游植物群落中甲藻/硅藻比虽然相对于2010年有很大的下降, 但先前长期监测结果显示近30年间, 胶州湾海域甲藻/硅藻比总体趋势是逐渐升高的(孙晓霞等, 2011; 孙松等, 2015), 这次下降可能是年际间波动, 其具体变化规律还需要通过长尺度的连续监测揭示。
 |
| 图 4 胶州湾硅藻丰度、甲藻丰度和甲藻/硅藻比的季节变化 Fig. 4 Seasonal variations in diatom and dinoflagellate densities and dinoflagellate/diatom ratio in the Jiaozhou Bay 注: a:胶州湾硅藻和甲藻丰度的季节性变化; b:胶州湾甲藻/硅藻比值的季节性变化 |
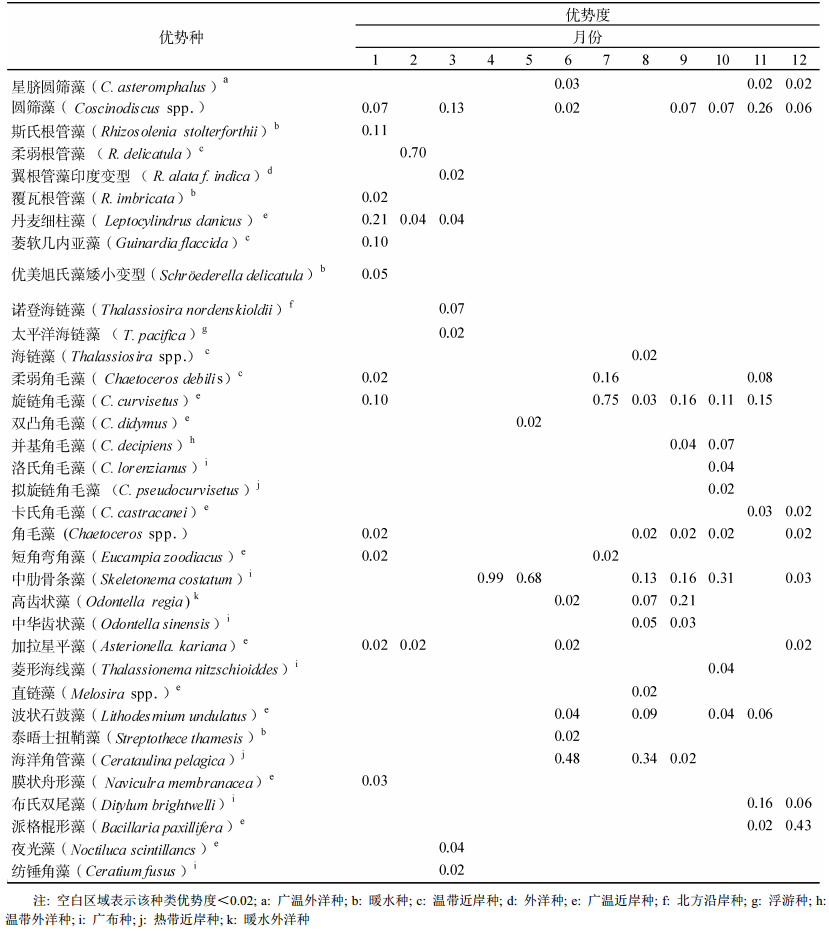
将优势度≥0.02的浮游植物定为优势种, 表 1显示2011年胶州湾浮游植物优势种组成具有明显的季节性差异。
表 1 胶州湾浮游植物优势种及优势度
Tab. 1 Dominant species and the predominancy of phytoplankton in the Jiaozhou Bay
|
冬季(1—3月)浮游植物的多样性最为丰富, 有圆筛藻、斯氏根管藻、柔弱根管藻、翼根管藻印度变型、覆瓦根管藻、丹麦细柱藻、萎软几内亚藻、优美旭氏藻矮小变型、诺登海链藻、太平洋海链藻、柔弱角毛藻、旋链角毛藻、角毛藻、短角弯角藻、加拉星杆藻、膜状舟形藻、纺锤角藻和夜光藻, 其中优势地位明显的是丹麦细柱藻; 秋季(10—12月)浮游植物优势种多样性次之, 有星脐圆筛藻、圆筛藻、柔弱角毛藻、旋链角毛藻、并基角毛藻、洛氏角毛藻、拟旋链角毛藻、卡氏角毛藻、角毛藻、中肋骨条藻、加拉星杆藻、菱形海线藻、波状石鼓藻、布氏双尾藻和派格棍形藻, 其中主要是某些圆筛藻占优势; 夏季(7—9月)优势种有圆筛藻、海链藻、柔弱角毛藻、并基角毛藻、角毛藻、短角弯角藻、中肋骨条藻、高齿状藻、中华齿状藻、直链藻、波状石鼓藻、大洋角管藻, 其中旋链角毛藻占优势地位; 春季(4—6月)优势种多样性最少, 有星脐圆筛藻、圆筛藻、双凸角毛藻、中肋骨条藻、高齿状藻、加拉星杆藻、波状石鼓藻、泰晤士扭鞘藻和大洋角管藻, 其中优势地位明显的是中肋骨条藻。
全年大部分月份均占优势地位的种类为圆筛藻、旋链角毛藻和中肋骨条藻。圆筛藻在各个季节均出现, 在11月份优势度相对较高, 为0.26;旋链角毛藻主要出现在夏秋季节, 在夏季优势度较高, 能达到0.75;中肋骨条藻在春季的优势地位很明显, 在4月和5月份的优势度分别能达到0.99和0.68。在优势种种类数方面, 与2010年调查结果不同的是, 2010年优势种种类数最多的季节是夏季, 其次才是秋季和冬季, 春季优势种种类数最少(郑珊等, 2014); 而本研究中发现2011年冬季的浮游植物优势种种类最多, 其次是秋季和夏季, 春季优势种种数少, 全年呈现优势种种类数先下降后逐渐上升的趋势。
与2004—2008年期间以及2010年胶州湾浮游植物优势种的调查结果作比较(孙晓霞等, 2011; 郑珊等, 2014), 发现2011年胶州湾全年占优势的种类和2004—2008年相似, 主要有中肋骨条藻和角毛藻, 但与2010年的调查结果不同, 后者主要是尖刺拟菱形藻和派格棍形藻。2011年调查期间, 尖刺拟菱形藻虽然在1—5月及10—12月期间均有出现, 但是优势度很低, 均小于0.004, 而派格棍形藻虽然也是在全年频繁出现, 但除在秋季优势度较高外, 别的月份均小于0.01。相对于上述6年的历史调查资料, 翼根管藻印度变型、萎软几内亚藻、优美旭氏藻矮小变型、双凸角毛藻、并基角毛藻、菱形海线藻、泰晤士扭鞘藻成为了2011年出现的新优势种。
2.3 生态类型如表 1所示, 2011年胶州湾浮游植物主要有以下几个生态类型:
广温外洋型:星脐圆筛藻。
广温近岸型:丹麦细柱藻, 旋链角毛藻, 双凸角毛藻, 卡氏角毛藻, 短角弯角藻, 加拉星平藻, 直链藻, 波状石鼓藻, 膜状舟形藻, 派格棍形藻, 夜光藻。
暖水型:斯氏根管藻, 覆瓦根管藻, 优美旭氏藻矮小变型, 泰晤士扭鞘藻。
温带近岸型:柔弱根管藻, 萎软几内亚藻, 海链藻, 柔弱角毛藻。
外洋型:翼根管藻印度变型。
北方沿岸型:诺登海链藻。
浮游型:太平洋海链藻。
温带外洋型:并基角毛藻。
广布型:洛氏角毛藻, 中肋骨条藻, 中华齿状藻, 菱形海线藻, 布氏双尾藻, 纺锤角藻。
热带近岸型:拟旋链角毛藻, 海洋角管藻。
暖水外洋型:高齿状藻。
由此可见, 调查期间胶州湾海域浮游植物优势种主要以广温近岸种、广布种为主, 在个别季节也出现了少数暖水种和外洋种。
2.4 多样性指数(H′)与均匀度(J)2011年胶州湾浮游植物多样性指数有明显的的季节性变化。多样性指数1月份最高, 为4.15;而4月份最低, 仅有0.08。全年多样性指数平均值为2.39。结合图 3和图 4可以发现, 4月份的浮游植物总丰度是全年最高, 但是多样性指数却是全年最低, 这是由于当月中肋骨条藻的优势度占绝对优势, 高达0.99, 其丰度达到587.8×106cell/m3, 形成中肋骨条藻春季赤潮, 导致该月份的种类组成较为单一, 因此多样性指数很低。而3月以及8—12月份虽然浮游植物总丰度较低, 但是多样性指数均大于3。若以C1、C3、C4、B2、A3和A5代表湾内区域, 以D1、D3和D5代表湾口区域, 以其余站位代表湾外区域, 可见这三个海域的浮游植物多样性指数的周年季节变化趋势类似(图 5a所示):从1月至4月下降, 在5—6月稍微回升, 在7月份出现一个峰谷, 8月份再次升高, 直到12月份均维持在较高的水平。并且湾外浮游植物均匀度相对最高, 其次是湾口, 湾内最低。
 |
| 图 5 胶州湾浮游植物多样性指数H′和均匀度J的周年变化 Fig. 5 Annual variation in diversity index H′ and equitability of phytoplankton in the Jiaozhou Bay |
胶州湾海域浮游植物平均均匀度值为0.26。如图 5b所示, 若以C1、C3、C4、B2、A3和A5代表湾内区域, 以D1、D3和D5代表湾口区域, 以其余站位代表湾外区域, 可见这三个海域的浮游植物均匀度J值周年季节变化趋势类似:从1月至4月下降, 在5—6月达到峰值, 7月份出现一个峰谷, 8月份再次升高, 并在之后缓慢下降。并且湾外浮游植物均匀度相对最高, 其次是湾口, 湾内最低。结合多样性指数的结果来看, 胶州湾不同区域多样性指数和均匀度的变化趋势基本一致, 多样性指数高、均匀度高, 浮游植物群落结构就更加稳定。由此可见, 浮游植物群落结构湾外比湾口和湾内更加均匀稳定, 可能是因为湾内海域环境受陆源污染和人工养殖的影响更加严重, 对浮游植物的生长繁殖存在一定的影响。
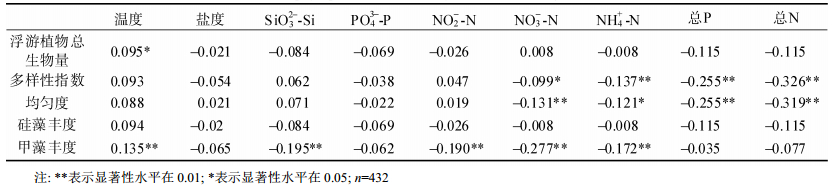
2.5 主要环境因子分析浮游植物群落结构与主要环境因子参数的相关性分析如表 2所示。结果显示:温度和浮游植物总生物量呈现显著正相关, 其中甲藻细胞丰度与温度的正相关关系尤为明显。而盐度和浮游植物群落结构的相关性不明显, 可能由于胶州湾盐度全年较为稳定, 为30.39±0.36。温度与甲藻丰度呈现极显著正相关关系, 硅酸盐浓度和甲藻丰度呈现极显著负相关关系, 表明硅酸盐对甲藻的生长繁殖存在明显的抑制效应。亚硝酸盐浓度的变化与甲藻丰度的相关关系与硅酸盐浓度相同; 硝酸盐与铵盐对甲藻丰度、浮游植物多样性指数以及均匀度的影响趋势一致, 均呈现显著或者极显著的负相关关系。总氮和总磷对浮游植物多样性指数和均匀度的影响趋势一致, 呈现极显著负相关关系。由此可见, 代表富营养化程度的总氮和总磷会影响浮游植物的多样性指数和均匀度, 进而影响浮游植物群落组成。而不同的营养盐结构比例也将对甲藻丰度、多样性指数以及均匀度存在影响。其中亚硝酸盐、硝酸盐、铵盐和硅酸盐的增加将可能从一定程度上抑制甲藻的生长。
表 2 胶州湾浮游植物群落结构与主要环境因子的相关性分析
Tab. 2 Correlation between phytoplankton community structure and major environmental factors
|
(1) 2011年胶州湾海域共检出浮游植物3门41属108种(包括变种和变型), 主要由硅藻和甲藻两大类组成, 硅藻占绝对优势地位。全年甲藻/硅藻比均小于0.08。生态类型主要以近岸广布种和暖温带种为主, 在不同季节也出现了少数暖水种和外洋种, 夏季和冬季优势种交替明显。
(2) 浮游植物丰度分别在2月、4月和8月份达到峰值, 其中4月为全年最高峰, 数量为591× 106cell/m3, 2月为冬季高峰, 数量是468×106cell/m3, 7月为夏季高峰, 数量为172×106cell/m3, 秋季浮游植物丰度相对较低。
(3) 浮游植物群落的多样性指数H′值平均为2.4, 秋、冬季节相对较高, 春、夏季节相对较低; 均匀度J平均为0.26。比较胶州湾不同区域浮游植物群落多样性指数和均匀度的周年变化显示, 胶州湾外浮游植物群落结构相对于湾口和湾内更为稳定。
(4) 温度、氮盐和硅酸盐对浮游植物群落结构存在明显影响。
致谢 本研究中的海洋调查和样品采集工作得到了中国科学院海洋研究所海洋生态系统研究站和浮游动物研究组各位老师、同学及“创新号”考察船全体工作人员的支持, 谨此致谢。
参考文献
| 刘东艳, 孙军, 唐优才, 等, 2002. 胶州湾北部水域浮游植物研究I——种类组成和数量变化. 青岛海洋大学学报, 32 (1) : 67–72 |
| 齐雨藻, 邹景忠, 梁松, 等, 2004. 中国沿海赤潮. 北京: 科学出版社, 1-348 |
| 孙军, 刘东艳, 2002. 中国海区常见浮游植物种名更改初步意见. 海洋与湖沼, 33 (3) : 271–286 |
| 孙松, 孙晓霞, 2015. 海湾生态系统的理论与实践——以胶州湾为例. 北京: 科学出版社, 213-226 |
| 孙松, 张永山, 吴玉霖, 等, 2005. 胶州湾初级生产力周年变化. 海洋与湖沼, 36 (6) : 481–486 |
| 孙晓霞, 孙松, 吴玉霖, 等, 2011. 胶州湾网采浮游植物群落结构的长期变化. 海洋与湖沼, 42 (5) : 639–646 |
| 李艳, 李瑞香, 王宗灵, 等, 2005. 胶州湾浮游植物群落结构及其变化的初步研究. 海洋科学进展, 23 (3) : 328–334 |
| 李广玉, 鲁静, 何拥军, 2005. 胶州湾浮游植物多样性及其与环境因子的关系. 海洋地质动态, 21 (4) : 10–13 |
| 杨世民, 王丽莎, 石晓勇, 2014. 2009年春季胶州湾浮游植物群落结构特征. 海洋与湖沼, 45 (6) : 1234–1240 |
| 杨世民, 王丽莎, 石晓勇, 2015. 2009年夏季胶州湾同步调查浮游植物群落结构特征. 海洋与湖沼, 46 (1) : 102–108 |
| 杨世民, 董树刚, 2006. 中国海域常见浮游硅藻图谱. 青岛: 中国海洋大学出版社, 1-267 |
| 吴玉霖, 孙松, 张永山, 等, 2004. 胶州湾浮游植物数量长期动态变化的研究. 海洋与湖沼, 35 (6) : 518–523 |
| 吴玉霖, 孙松, 张永山, 等, 2005. 环境长期变化对胶州湾浮游植物群落结构的影响. 海洋与湖沼, 36 (6) : 487–498 |
| 陈碧鹃, 陈聚法, 袁有宪, 等, 2000. 胶州湾北部沿岸浮游植物生态特征的研究. 海洋水产研究, 21 (2) : 34–40 |
| 国家技术监督局, 1992. 海洋调查规范. 北京: 中国标准出版社, 17-22 |
| 金德祥, 陈金环, 黄凯歌, 1965. 中国海洋浮游硅藻类. 上海: 上海科学技术出版社, 1-230 |
| 郑珊, 孙晓霞, 赵永芳, 等, 2014. 2010年胶州湾网采浮游植物种类组成与数量的周年变化. 海洋科学, 38 (11) : 1–6 |
| 钱树本, 王筱庆, 陈国蔚, 1983. 胶州湾的浮游藻类. 山东海洋学院学报, 13 (1) : 39–56 |
| 郭玉洁, 钱树本, 2003. 中国海藻志:第五卷:硅藻门:第一册:中心纲. 北京: 科学出版社, 1-493 |
| Lalli C M, Parsons T R, 1993. Biological Oceanography. An Introduction. New York: Pergamon Press, 45-79 |
| Liu D Y, Sun J, Liu Z, et al, 2004. The effects of spring-neap tide on the phytoplankton community development in the Jiaozhou Bay, China. Acta Oceanologica Sinica, 23 (4) : 687–697 |
| Pielou E C, 1969. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, 1-286 |
| Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana Ill: University of Illinois Press, 1-125 |
| Tomas C R, 1997. Identifying Marine Phytoplankton. San Diego: Academic Press, 1-858 |