 2016, Vol. 47
2016, Vol. 47中国海洋湖沼学会主办。
文章信息
- 宋秀凯, 汤宪春, 程玲, 孙伟, 张焕君, 姜会超, 刘元进. 2016.
- SONG Xiu-Kai, TANG Xian-Chun, CHENG Ling, SUN Wei, ZHANG Huan-Jun, JIANG Hui-Chao, LIU Yuan-Jin. 2016.
- 崂山湾网采浮游植物时空分布特征及其与环境因子关系
- SPATIO-TEMPORAL DISTRIBUTION OF NET-COLLECTED PHYTOPLANKTON AND ITS RELATIONSHIP WITH ENVIRONMENTAL FACTORS IN LAOSHAN BAY
- 海洋与湖沼, 47(6): 1205-1213
- Oceanologia et Limnologia Sinica, 47(6): 1205-1213.
- http://dx.doi.org/10.11693/hyhz20160600127
-
文章历史
- 收稿日期:2016-06-20
- 收修改稿日期:2016-07-20




海湾是海岸带的重要组成部分, 是海陆之间相互作用最为强烈的区域之一, 在物质循环和能量传递间发挥着重要作用, 同时也是许多海洋生物重要的索饵场和洄游通道。近年来随着海湾开发的不断深入, 海湾的生态环境受到影响逐年增大, 海水污染日趋严重, 生物多样性日益降低等(王文海等, 2011), 海湾生态环境问题已成为一个全球性的问题, 迫切需要针对具体的海湾生态系统开展深入研究。国内许多学者对渤海湾(康元德, 1991; 俞建銮等, 1993; 王俊等, 1998; 王俊, 2003; 周然等, 2013)、莱州湾(陈碧鹃等, 2001; 郝彦菊等, 2005; 李广楼等, 2006; 宁璇璇等, 2011)和胶州湾(李艳等, 2005; 孙晓霞, 2011; 王妍等, 2013)等进行了大量研究, 但崂山湾生态系统研究较少。崂山湾位于黄海的胶州湾和丁字湾之间, 海湾开阔, 总面积163.13 km2, 是山东省魁蚶等贝类增殖放流的重点海域(张波等, 2015)。全面了解该海湾的浮游植物, 特别是贝类饵料生物的群落结构和季节变化趋势对于贝类资源放流至关重要。
浮游植物是海洋食物链的基础环节, 对栖息生境中的各种环境因子有着较强的依赖性, 其种类组成和数量分布等生态特征在一定程度上反映了海域生态环境的基本特征(Lalli et al, 1993), 同时环境条件的改变也直接或间接地影响到浮游植物的群落结构。由于浮游植物是贝类的重要饵料, 弄清该海域浮游植物的种类组成、丰度平面分布及其丰度的季节变化规律对于放流区域的选择、放流量的多寡密切相关。本文通过2012-2014年连续3年12个航次的浮游植物调查, 分析了崂山湾浮游植物种类和数量年际变化特征、优势种的变迁及其与环境因子关系, 为崂山湾开展魁蚶等贝类资源放流和实施有效的渔业保护提供科学依据。
1 材料与方法 1.1 样品采集与分析2012-2014年每年3月、5月、8月和11月, 在崂山湾海域(120.6°-121°E, 36.0°-36.5°N)进行12个航次监测, 共设14个监测站位(见图 1)。按《海洋调查规范》(GB12763.6-2007)规定, 用浅水Ⅲ型浮游生物网自底层至表层垂直拖网采集浮游植物, 样品用鲁格氏碘液固定, 实验室内进行分类、鉴定、计数和统计。同步调查水温(Water Temperature, WT)、盐度(Salinity)、溶解氧(Dissolved Oxygen, DO)、化学耗氧量(Chemical Oxygen Demand, COD)、pH、无机氮(Inorganic Nitrogen, DIN)、活性磷酸盐(Active Phosphorus, AP)、总氮(Total Nitrogen, TN)、总磷(Total Phosphorus, TP)及叶绿素a (chlorophyll a, chl a)等环境参数, 所有操作均按照《海洋监测规范》(GB17378.4-2007)进行。
 |
| 图 1 崂山湾浮游植物采样站位 Fig. 1 Sampling sites of phytoplankton in Laoshan Bay |
浮游植物丰度以每立方米出现的细胞数表示(cell/m3)。
优势种的优势度: Y=(ni/N′)×fi
式中, ni为第i种的丰度, fi为该种在各站位中出现的频率, N′为总丰度。根据种类优势度公式计算各种生物的优势度, 将Y≥0.02的生物定为优势种(钱迎倩等, 1994)。
物种多样性指数H′采用Shannon-Wiener指数(Shannon et al, 1949): 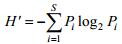
物种丰富度指数dMa采用Margalef指数计算公式(Margalef, 1968): dMa=(S-1)/log2N
均匀度指数J采用Pielou指数计算公式(Pielou, 1969): J=H′/log2S
式中, N为采集样品中所有物种的总个体数, S为样品中的种类总数, Pi为第i种的个体数与样品中的总个体数的比值。
1.2.2 群落结构分析采用生物统计分析软件PRIMER 6.0进行群落结构分析(周红等, 2003), 原始的丰度数据经平方根转化后, 以Bray Curtis相似性系数为基础构建相似性矩阵, 然后使用等级聚类分析CLUSTER将样品按监测时间逐级连接成组, 通过树枝图来表示群落结构年际变化。用SIMPER (Similarity percentage program)分析来计算不同物种对样本组内相似性和组间差异性的平均贡献率。
1.2.3 平面分布及相关性分析浮游植物丰度平面分布图用Surfer8.0软件绘制, 浮游植物细胞丰度及群落结构与环境因子相关关系用SPSS19.0软件分析。
2 结果 2.1 浮游植物种类组成及优势种变迁崂山湾海域共鉴定出浮游植物127种, 隶属于硅藻(Bacillariophyceae)、甲藻(Pyrrophyta)、金藻(Chrysophyta)、蓝藻(Cyanophyta)、绿藻(Chlorophyta) 5大门类25科47属。其中硅藻16科36属105种, 占种类数的82.7%;甲藻6科8属19种, 占种类数的15.0%;金藻、蓝藻、绿藻各1科1属1种。
作为海区的主要优势种, 每个季节优势度前三位的优势种随季节更替明显。3月份主要优势种为冰河拟星杆藻(Asterionopsis glacialis, Y=0.079-0.294)、中肋骨条藻(Skeletonema costatum, Y=0.036-0.517)、加拉星杆藻(Asteroplanus karianus, Y=0.028-0.272)、布氏双尾藻(Ditylum brightwellii)、尖刺拟菱形藻(Pseudo-nitzschia pungens)、夜光藻(Noctiluca scintillans)等; 5月份主要优势种为夜光藻(Y=0.126-0.636)、尖刺拟菱形藻(Y=0.021-0.142)、密联角毛藻(Chaetoceros densus, Y≤0.02-0.060)、派格棍形藻(Bacillaria paxillifera)等; 8月份主要优势种为三角角藻(Ceratium tripos, Y=0.201-0.242)、尖刺拟菱形藻(Y=0.121-0.218)、派格棍形藻(Y≤0.02-0.142)、菱形海线藻(Thalassionema nitzschioide)、梭角藻(Ceratium fusus)、夜光藻等; 11月份主要优势种为虹彩圆筛藻(Coscinodiscus oculus-iridis, Y=0.074-0.107)、派格棍形藻(Y=0.068-0.142)、冰河拟星杆藻(Y=0.021-0.036)、加拉星杆藻、密联角毛藻、旋链角毛藻(Chaetoceros curvisetus)等(表 1)。
| 优势种 | 优势度(Y) | ||||||||||||||
| 3月 | 5月 | 8月 | 11月 | ||||||||||||
| 2013.03 | 2014.03 | 2015.03 | 2012.05 | 2013.05 | 2014.05 | 2012.08 | 2013.08 | 2014.08 | 2012.11 | 2013.11 | 2014.11 | ||||
| 布氏双尾藻Ditylum brightwellii | 0.047 | 0.025 | 0.055 | ||||||||||||
| 虹彩圆筛藻 Coscinodiscus oculus-iridis |
0.040 | 0.091 | 0.074 | 0.107 | 0.105 | ||||||||||
| 加拉星杆藻 Asteroplanus karianus |
0.272 | 0.028 | 0.036 | 0.033 | 0.153 | ||||||||||
| 尖刺拟菱形藻 Pseudo-nitzschia pungens |
0.052 | 0.063 | 0.142 | 0.021 | 0.052 | 0.218 | 0.186 | 0.121 | 0.054 | ||||||
| 菱形海线藻 Thalassionema nitzschioide |
0.023 | 0.089 | 0.032 | ||||||||||||
| 密联角毛藻Chaetoceros densus | 0.029 | 0.060 | 0.026 | 0.125 | 0.021 | 0.045 | |||||||||
| 派格棍形藻Bacillaria paxillifera | 0.023 | 0.123 | 0.068 | 0.142 | 0.074 | 0.068 | |||||||||
| 冰河拟星杆藻 Asterionopsis glacialis |
0.223 | 0.294 | 0.079 | 0.036 | 0.021 | 0.029 | |||||||||
| 旋链角毛藻Chaetoceros curvisetus | 0.050 | ||||||||||||||
| 印度翼根管藻 Rhizosolenia alata f. indica |
0.053 | 0.023 | |||||||||||||
| 中肋骨条藻Skeletonema costatum | 0.036 | 0.457 | 0.517 | 0.023 | 0.044 | 0.037 | 0.022 | ||||||||
| 三角角藻Ceratium tripos | 0.042 | 0.036 | 0.028 | 0.022 | 0.203 | 0.201 | 0.242 | ||||||||
| 梭角藻Ceratium fusus | 0.027 | 0.028 | 0.023 | 0.056 | |||||||||||
| 夜光藻Noctiluca scintillans | 0.042 | 0.052 | 0.185 | 0.636 | 0.126 | 0.047 | 0.036 | 0.025 | |||||||
| 注:优势度≥0.02的物种为该航次的优势种, 表格中粗体数字为该月份海区前两位优势种的优势度 | |||||||||||||||
浮游植物丰度年内分布呈现较为明显的双峰型季节变化曲线, 如图 2a所示。3月份丰度最高, 为第一高峰期, 该季节丰度范围108.7×104-205.9× 104 cell/m3, 平均169.2×104 cell/m3, 2014年3月为所有航次最高值。该季度浮游植物类群以中肋骨条藻、冰河拟星杆藻等硅藻为主, 数量百分比约85.5%-95.5%;甲藻类群以夜光藻为主, 约占总数量的3.4%-12.4%, 监测期间该月份浮游植物类群年际间变化不大, 种类数和各类群丰度相对稳定(图 2b)。8月为次高峰期, 丰度范围34.3×104-82.5× 104 cell/m3, 平均51.8×104 cell/m3。该季度浮游植物类群中三角角藻、梭角藻等甲藻数量较高, 约占总数量的19.5%-50.5%, 特别是2013年8月甲藻比例超过硅藻成为主要类群。11月份丰度范围7.7×104-52.2× 104 cell/m3, 平均26.6×104 cell/m3。该季节浮游植物类群以虹彩圆筛藻、派格棍形藻、冰河拟星杆藻等硅藻为主, 数量百分比约85.5%-95.5%;甲藻数量很低, 数量百分比仅为2.8%-7.7%。5月份丰度最低, 范围7.6×104-35.9×104 cell/m3, 平均17.2×104 cell/m3。该季节夜光藻大量繁殖成为主要优势种之一, 甲藻数量百分比为所有监测航次最高值, 比例范围19.5%-60.5%, 特别是2013年5月, 甲藻为海区主要种群, 有诱发夜光藻赤潮的风险。
 |
| 图 2 浮游植物细胞丰度变化(a)与各类群数量百分比(b) Fig. 2 Variation of phytoplankton cell abundance (a), and the percentage (b) from 2012 to 2014 |
崂山湾浮游植物丰度平面分布极不均匀, 总体上近岸海域浮游植物丰度大于远海。鳌山卫近岸海域各季节浮游植物丰度均较高, 其次为田横岛附近海域, 崂山湾东南部宽阔海域数量相对较低, 见图 3。
 |
| 图 3 崂山浮游植物丰度平面分布 Fig. 3 Horizontal distribution of cell abundance of phytoplankton in Laoshan Bay |
三个年度每年3月, 浮游植物丰度高值区出现在田横岛西侧和鳌山卫东北面的内弯处, 低值区位于远离崂山湾的外侧海域。2014年5月, 高值区依然为田横岛西侧和鳌山卫东北面的内弯处, 2012和2013年度5月份高值区位于鳌山卫西南和王哥庄东面的近岸海域, 低值区均为远离崂山湾的外侧海域。2012年8月浮游植物丰度呈现以田横岛附近往西南方向递减趋势, 2013年8月度丰度呈现以鳌山卫近岸往东南方向递减趋势, 2014 年8月则由湾外向内呈递减趋势。2012年11月和2014年11月变化趋势同3月份, 由田横岛西侧和鳌山卫东北面的内弯处往外递减, 2013年11月则相反, 由湾外向内呈递减趋势。
2.4 浮游植物群落结构年际变化图 4所示为2012-2014年浮游植物群落结构参数(多样性指数、丰富度指数和均匀度指数)变化情况。Shannon-Wiener多样性指数(H′)范围1.829-2.745, 平均2.305, 最高值出现在2013年8月, 最低值出现在2013年3月。Margalef丰富度指数(D)范围0.634-1.507, 平均0.987, 最高值出现在2013年8月, 最低值出现在2012年5月, 其变化规律与Shannon-Wiener指数类似。Pielou均匀度指数(J)范围0.466-0.815, 平均0.675, 最高值出现在2012年5月, 最低值出现在2014年3月。
 |
| 图 4 丰富度指数、多样性指数、均匀度指数年际变化 Fig. 4 Variation in richness, diversity, and evenness of phytoplankton community |
对各个航次的浮游植物群落种类丰度进行平方根转化, 做出Bray-Curtis相似性矩阵, 以此矩阵进行CLUSTER聚类和MDS标序, 结果见图。调查海域浮游植物群落落在42%相似性水平大致可分为4个类群(图 5), 同时使用SIMPER分析了各组分的优势物种。
 |
| 图 5 崂山湾12个监测月份浮游植物群落的CLUSTER聚类和MDS标序 Fig. 5 CUSTER and MDS ordination of phytoplankton community in 12 months in Laoshan Bay |
类群1: 2014年8月和2012年8月的浮游植物群落相似性约42.9%, 这两个月份细胞丰度接近, 甲藻比例相似, 优势种为三角角藻、尖刺拟菱形藻、菱形海线藻等, 贡献率分别为19.2%、15.4%、9.5%, 累计贡献率44.1%。
类群2: 2012年11月、2013年11月、2014年11月的浮游植物群落相似性约42.8%, 这三个月份浮游植物群落细胞丰度较低, 硅藻占据绝对优势, 优势种为虹彩圆筛藻、派格棍形藻、冰河拟星杆藻等, 贡献率分别为13.1%、11.8%和9.9%, 累计贡献率34.8%。
类群3: 2012年5月、2014年5月、2013年5月和8月浮游植物群落相似性约42.4%, 这四个月份浮游植物丰度偏低, 种类数较少, 夜光藻、尖刺拟菱形藻、密联角毛藻等为特征种, 贡献率分别为15.5%、8.4%、6.7%, 累计贡献率为30.6%。
类群4: 2012年3月、2013年3月、2015年3月浮游植物群落相似性约42.1%, 该月份水温较低, 细胞丰度为所监测季节最高的月份, 冰河拟星杆藻、中肋骨条藻、加拉星杆藻等硅藻占绝对优势, 贡献率分别为25.3%、18.8%、11.5%, 累计贡献率为55.6%。
2.6 浮游植物群落与环境因子关系浮游植物群落与水温、盐度、营养盐、叶绿素a等环境因子密切相关, 见表 2。浮游植物丰度很大程度上取决于硅藻丰度, 二者呈极显著正相关(R=0.985, P < 0.01), 并且与其它环境因子相关关系一致, 均与盐度、铵盐、叶绿素a、N/P呈极显著正相关, 相关系数分别为0.845、0.839、0.759、0.779和0.863、0.875、0.758、0.818 (P < 0.01), 与pH和无机氮显著正相关(R=0.604, 0.639和R=0.631, 0.679, P < 0.05), 而与水温呈显著负相关(R=-0.590和R=-0.666, P < 0.05)。甲藻丰度与无机氮和铵盐含量呈显著负相关, 相关系数分别为R=-0.799和R=-0.642 (P < 0.05)。物种多样性指数与化学需氧量显著正相关(R=0.594, P < 0.05), 与无机氮、总磷和活性磷酸盐显著负相关(R=-0.617, -0.628, -0.629, P < 0.05)。
| 理化指标 | 浮游植物丰度 | 硅藻丰度 | 甲藻丰度 | 多样性指数 |
| 浮游植物丰度 | 1 | 0.985** | 0.285 | -0.383 |
| 硅藻丰度 | 0.985** | 1 | 0.116 | -0.406 |
| 甲藻丰度 | 0.285 | 0.116 | 1 | 0.049 |
| 多样性指数 | -0.383 | -0.406 | 0.049 | 1 |
| 水温 | -0.590* | -0.666* | 0.304 | 0.432 |
| 盐度 | 0.845** | 0.863** | 0.068 | -0.367 |
| 酸碱度pH | 0.604* | 0.631* | -0.014 | -0.326 |
| 溶解氧 | -0.572 | -0.511 | -0.449 | 0.305 |
| 化学需氧量COD | 0.197 | 0.190 | 0.064 | 0.594* |
| 总氮 | -0.146 | -0.135 | -0.115 | -0.189 |
| 总磷 | -0.226 | -0.224 | -0.070 | -0.628* |
| 无机氮 | 0.639* | 0.679* | -0.799* | -0.617* |
| 铵盐 | 0.839** | 0.875** | -0.642* | -0.549 |
| 亚硝酸盐 | -0.436 | -0.369 | -0.475 | -0.029 |
| 硝酸盐 | 0.039 | 0.057 | -0.107 | -0.341 |
| 活性磷酸盐 | -0.290 | -0.270 | -0.192 | -0.629* |
| 叶绿素a | 0.759** | 0.758** | 0.147 | -0.519 |
| 氮磷比 | 0.779** | 0.818** | -0.043 | -0.142 |
| **表示相关置信度水平 < 0.01, *表示相关置信度水平 < 0.05, 其余没有明显相关关系 | ||||
崂山湾是我国重要的贝类增养殖放流基地, 浮游植物群落结构季节性变化明显。据中国海湾志记载(中国海湾志编纂委员会, 1993), 崂山湾浮游植物群落结构季节变化明显, 细胞丰度表现为春夏季低, 秋冬季高的特点, 这与本调查结果变化趋势基本一致, 但高峰期出现的时间些许不同。崂山湾浮游植物丰度高峰期出现在3月和8月, 低值期出现在5月和11月, 与地处渤海的莱州湾具有上、下半年各有一次数量高峰的结论一致(陈碧鹃等, 2001), 崂山湾同样一年内具有两个细胞丰度高峰期。其中3月平均丰度169.2×104 cell/m3, 低于海湾志记载的3650×104 cell/m3, 与2013年3月乔芮等(2015)在该海域同期调查的平均丰度314×104 cell/m3一致, 与同处北黄海的胶州湾相似, 3月份为该海域浮游植物丰度最高峰(王妍等, 2013), 此时第一优势种为冰河拟星杆藻, 与中国海湾志记载、乔芮研究结果一致, 该月份优势种没有发生明显改变。5月平均丰度17.2×104 cell/m3, 为监测的丰度最低值, 该季节夜光藻大量繁殖成为主要优势种之一, 此时海区夜光藻平均丰度7.02×104 cell/m3, 与乔芮同期研究结果一致(7.09×104 cell/m3), 此时期甲藻数量百分比为所有监测航次最高值, 比例范围19.5%-60.5%, 这与渤海湾(周然等, 2013)、莱州湾(陈碧鹃等, 2001; 李广楼等, 2006)、胶州湾(孙晓霞等, 2011)等研究发现5月份夜光藻为海区主要优势种结论一致。8月为下半年丰度的高峰期, 此时三角角藻、梭角藻等甲藻成为优势种, 该月份海区出现浒苔绿潮消退后期, 调查时仅发现少量浒苔漂浮在水面上, 大面积聚集情形已消退。由于营养盐被大量消耗, 此时海区无机氮含量仅为0.099±0.065 mg/L, 活性磷酸盐0.006±0.005 mg/L, 由于硅藻对磷酸盐等营养元素需求较甲藻要求高(Egge, 1998), 低磷营养环境限制了硅藻的生长, 甲藻取代硅藻成为优势种, 这与2008年宋秀凯等在浒苔后期发现的低磷营养条件下三角角藻等甲藻大量繁殖成为主要优势种的结果一致(宋秀凯等, 2011)。
海洋环境是海洋生物赖以生存的基础, 海洋生物的活动分布、繁殖和生长都与海洋环境密不可分。崂山湾海域浮游植物细胞丰度由近岸向远海呈逐渐递减趋势, 其中田横岛和鳌山卫近岸为高值区, 远离岸边的区域丰度逐渐降低, 这与海湾近海海域盐度一般较低、营养盐比较丰富有关, 这一平面分布规律与莱州湾、胶州湾等其它海湾浮游植物细胞丰度分布规律一致(朱树屏, 1966; 康元德, 1991; 王俊等, 1998; Gao et al, 2013)。浮游植物群落结构与环境因子密切相关(Zhao et al, 2005; Thillai et al, 2010; 杨晓改等, 2014), 研究海域硅藻丰度与盐度、铵盐、叶绿素a、N/P等环境参数极显著正相关(P < 0.01), 与pH和无机氮显著正相关(P < 0.05), 与水温呈显著负相关。甲藻丰度与硅藻则相反, 与无机氮和铵盐含量呈显著负相关(P < 0.01)。环境因子中的水温、营养盐和盐度等为影响崂山湾海域浮游植物群落演替的主要因子, 群落组成在42%的相似性水平分为春夏秋冬四个类群, 这与王欣等2011年在该海域湾外研究的结果基本一致, 磷酸盐和水温是影响这一区域物种分布格局的重要因素(王欣等, 2014), 群落组成同样在42%的相似性下分为春夏秋冬4个类群。
4 结语(1) 2012-2014年度共采集到浮游植物127种, 隶属5大门类25科47属, 优势种季节更替明显。3月份主要优势种为冰河拟星杆藻、中肋骨条藻和加拉星杆藻等, 5月优势种为夜光藻、尖刺拟菱形藻和密联角毛藻等, 8月优势种为三角角藻、尖刺拟菱形藻和派格棍形藻等, 11月份优势种为虹彩圆筛藻、派格棍形藻和冰河拟星杆藻, 浮游植物群落年际相似性在42%水平聚类分为春夏秋冬四个类群。
(2) 崂山湾浮游植物丰度季节更替明显, 上、下半年均有一个高峰期, 上半年高峰期出现在3月, 平均丰度169.2×104 cell/m3, 下半年高峰期在8月, 平均丰度51.8×104 cell/m3。整体上浮游植物丰度高值区出现在田横岛西侧和鳌山卫东北面的内弯处, 低值区位于远离崂山湾的外侧海域。群落Shannon-Wiener多样性指数平均2.305 (1.829-2.745), Margalef丰富度指数平均0.987 (0.634-1.507), Pielou均匀度指数平均0.675 (0.466-0.815), 群落结构稳定。
(3) 浮游植物丰度很大程度上取决于硅藻丰度, 二者均与盐度、铵盐、叶绿素a、N/P呈极显著正相关(P < 0.01), 与pH和无机氮显著正相关(P < 0.05), 而与水温呈显著负相关(P < 0.05)。甲藻丰度与无机氮和铵盐含量呈显著负相关(P < 0.05)。物种多样性指数与化学需氧量显著正相关(P < 0.05), 与无机氮、总磷和活性磷酸盐显著负相关(P < 0.05)。
| 王妍, 张永, 王玉珏, 等, 2013. 胶州湾浮游植物的时空变化特征及其与环境因子的关系. 安全与环境学报, 13 (1) : 163–170 |
| 王欣, 盛化香, 唐衍力, 等, 2014. 崂山湾人工鱼礁区浮游植物群落结构与环境因子的关系. 渔业科学, 35 (4) : 7–12 |
| 王俊, 2003. 渤海近岸浮游植物种类组成及其数量变动的研究. 海洋水产研究, 24 (4) : 44–50 |
| 王俊, 康元德, 1998. 渤海浮游植物种群动态的研究. 海洋水产研究, 19 (1) : 43–52 |
| 王文海, 吴桑云, 丰爱平, 2011. 试论健康海湾与海湾健康指标. 海岸工程, 30 (3) : 90–97 |
| 中国海湾志编纂委员会, 1993. 中国海湾志-第四分册-山东半岛南部和江苏省海湾. 北京: 海洋出版社, |
| 宁璇璇, 纪灵, 王刚, 等, 2011. 2009年莱州湾近岸海域浮游植物群落的结构特征. 海洋湖沼通报, (3) : 97–104 |
| 朱树屏, 1966.黄河口附近海区浮游植物的季节变异.见:太平洋西部渔业研究委员会第八次全体会议论文集.北京:科学出版社, 1-10 |
| 乔芮, 栾青杉, 王俊, 等, 2015. 2013年春季崂山湾浮游植物群落及其环境控制. 海洋科学, 39 (7) : 13–21 |
| 孙晓霞, 孙松, 吴玉霖, 等, 2011. 胶州湾网采浮游植物群落结构的长期变化. 海洋与湖沼, 42 (5) : 639–646 |
| 李艳, 李瑞香, 王宗灵, 等, 2005. 胶州湾浮游植物群落结构及其变化的初步研究. 海洋科学进展, 23 (3) : 328–334 |
| 李广楼, 陈碧鹃, 崔毅, 等, 2006. 莱州湾浮游植物的生态特征. 中国水产科学, 13 (2) : 292–299 |
| 杨晓改, 薛莹, 昝肖肖, 等, 2014. 海州湾及其邻近海域浮游植物群落结构及其与环境因子的关系. 应用生态学报, 25 (7) : 2123–2131 |
| 宋秀凯, 张秀珍, 刘爱英, 等, 2011. 浒苔过量繁殖后海区环境因子与浮游植物群落演替规律研究. 安全与环境学报, 11 (2) : 150–157 |
| 张波, 袁伟, 王俊, 2015. 崂山湾春季鱼类群落的摄食生态及其主要种类. 中国水产科学, 22 (4) : 820–827 |
| 陈碧鹃, 陈聚法, 崔毅, 等, 2001. 莱州湾东部养殖区浮游植物的生态特征. 海洋水产研究, 22 (3) : 64–70 |
| 周红, 张志南, 2003. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用. 青岛海洋大学学报, 33 (1) : 58–64 |
| 周然, 彭士涛, 覃血波, 等, 2013. 渤海湾浮游植物与环境因子关系的多元分析. 环境科学, 34 (3) : 864–873 |
| 郝彦菊, 王宗灵, 朱明远, 等, 2005. 莱州湾营养盐与浮游植物多样性调查与评价研究. 海洋科学进展, 23 (2) : 197–204 |
| 俞建銮, 李瑞香, 1993. 渤海、黄海浮游植物生态的研究. 黄渤海海洋, (3) : 52–59 |
| 钱迎倩, 马克平, 1994. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 141-165 |
| 康元德, 1991. 渤海浮游植物的数量分布和季节变化. 海洋水产研究, (12) : 31–54 |
| Egge J K, 1998. Are diatoms poor competitors at low phosphate concentrations?. Journal of Marine Systems, 16 (3-4) : 191–198 DOI:10.1016/S0924-7963(97)00113-9 |
| Gao Y, Jiang Z B, Liu J J, et al, 2013. Seasonal variations of Net-phytoplankton community structure in the southern Yellow sea. Journal of Ocean University of China, 12 (4) : 557–567 DOI:10.1007/s11802-013-2258-x |
| Lalli C M, Parsons T R, 1993. Biological Oceanography:An Introduction. New York: Pergamon Press, 45-79 |
| Margalef R, 1968. Perspectives in Ecological Theory. Chicago: University of Chicago Press, |
| Pielou E C, 1969. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, 1-294 |
| Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana L: University of Illinois Press, 1-117 |
| Thillai R K, Rajkumar M, Sun J, et al, 2010. Seasonal variations of phytoplankton diversity in the Coleroon coastal waters, Southeast coast of India. Acta Oceanologica Sinica, 29 (5) : 97–108 DOI:10.1007/s13131-010-0068-1 |
| Zhao L, Wei H, 2005. The influence of physical factors on the variation of phytoplankton and nutrients in the Bohai Sea. Journal of Oceanography, 61 (2) : 335–342 DOI:10.1007/s10872-005-0044-0 |