 2017, Vol. 48
2017, Vol. 48

三门湾网采浮游植物季节变化及影响因素

http://dx.doi.org/10.11693/hyhz20160700153
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 陈悦, 刘晶晶, 高月鑫, 寿鹿, 廖一波, 黄伟, 江志兵. 2017.
- CHEN Yue, LIU Jing-Jing, GAO Yue-Xin, SHOU Lu, LIAO Yi-Bo, HUANG Wei, JIANG Zhi-Bing. 2017.
- 三门湾网采浮游植物季节变化及影响因素
- SEASONAL VARIATION AND THE FACTORS ON NET-PHYTOPLANKTON IN SANMEN BAY
- 海洋与湖沼, 48(1): 101-112
- Oceanologia et Limnologia Sinica, 48(1): 101-112.
- http://dx.doi.org/10.11693/hyhz20160700153
-
文章历史
- 收稿日期:2016-07-21
- 收修改稿日期:2016-09-12
引用本文
陈悦, 刘晶晶, 高月鑫, 寿鹿, 廖一波, 黄伟, 江志兵. 2017. 三门湾网采浮游植物季节变化及影响因素[J]. 海洋与湖沼, 48(1): 101-112
CHEN Yue, LIU Jing-Jing, GAO Yue-Xin, SHOU Lu, LIAO Yi-Bo, HUANG Wei, JIANG Zhi-Bing. 2017. SEASONAL VARIATION AND THE FACTORS ON NET-PHYTOPLANKTON IN SANMEN BAY[J]. Oceanologia et Limnologia Sinica, 48(1): 101-112.
三门湾网采浮游植物季节变化及影响因素
1.
国家海洋局第二海洋研究所 杭州 310012;
2. 国家海洋局海洋生态系统与生物地球化学重点实验室 杭州 310012
2. 国家海洋局海洋生态系统与生物地球化学重点实验室 杭州 310012
摘要:
于2015-2016年春(5月)、夏(8月)、秋(11月)和冬(1月)四个航次对三门湾18个站位进行拖网采集和理化因子测定,分析了三门湾浮游植物群落结构、优势种组成、时空分布和主要影响因素,并结合历史数据,分析了浮游植物群落结构的变化规律,探讨了其主要驱动因子。调查共检出浮游植物8门87属199种(含变种、变型和未定名种),其中硅藻51属149种,甲藻18属27种,绿藻、裸藻、隐藻、定鞭藻、金藻和蓝藻偶有检出。四季共检出优势种8属13种,其中琼氏圆筛藻(Coscinodiscus jonesianus)为全年优势种。浮游植物年均丰度为1632.04×104cell/m3,其中春季最低(53.06×104cell/m3),夏季最高(5548.91×104cell/m3)。聚类和相似性分析结果表明,浮游植物群落季节变化和区域差异明显。典范对应分析表明,影响三门湾海域浮游植物群落的主要因子依次为温度、盐度、DIN和DSi。结合历史资料发现,三门湾浮游植物群落丰度总体呈增加趋势(排除20世纪80年代冬季赤潮影响),主要种类组成仍以硅、甲藻为主,赤潮藻及嗜氮性硅藻种类数及优势度均上升。此外,三门湾浮游植物由大型硅藻向链状硅藻演替。海域富营养化和水温上升可能是主要驱动因子。
关键词:
浮游植物
环境因子
季节变化
典范对应分析
三门湾
SEASONAL VARIATION AND THE FACTORS ON NET-PHYTOPLANKTON IN SANMEN BAY
1.
Second Institute of Oceanography, State Oceanic Administration, Hangzhou 310012, China;
2. Key Laboratory of Marine Ecosystem and Biogeochemistry, State Oceanic Administration, Hangzhou 310012, China
2. Key Laboratory of Marine Ecosystem and Biogeochemistry, State Oceanic Administration, Hangzhou 310012, China
Abstract:
Sanmen Bay (28.96-29.18°E, 121.58-121.85°N), located off Zhejiang, East China, is a semi-enclosed bay in a relatively still water environment. To understand the main influential factors on the bay environment, we conducted seasonal marine surveys from May 2015 to January 2016 and the follow-up analyses on community structure, temporal-spatial variation, composition and variation of dominant phytoplankton species, and compared them against available historical data. Among all the samples collected, we identified 8 phyla, 87 genera, and 199 species, including 149 diatom species, 27 dinoflagellate species, and few other 6 species (chlorophyta, euglenophyta, cryptomonad, haptophyte, chrysophyta and cyanophyta). Among the total, 13 species of 8 genera were dominant in different seasons, of which Coscinodiscus jonesianus was an annual dominant species. The net-phytoplankton abundance showed a typical seasonal pattern; the highest average value appeared in summer (5548.91×104cell/m3) and the lowest in spring (53.06×104cell/m3), in annual average of 1632.04×104cell/m3. Both phytoplankton community parameters and the physico-chemical factors varied significantly (P<0.05) in season. Clusters and similarity analysis revealed significant variations in phytoplankton community in temporal and special realm, especially at bottom section and at the bay mouth. Canonical correspondence analysis (CCA) presented that temperature, nutrients, and salinity were the main factors on the phytoplankton community. In particular, the phytoplankton abundance has been increasing in the past 30 years, during which phytoplankton community succeeded from large diatom genus to chain-forming diatoms. We believe that the in-bay eutrophication and temperature increase were the main factors responsible for the succession.
Key words:
phytoplankton
environmental factor
seasonal variation
canonical correspondence analysis (CCA)
Sanmen Bay
三门湾位于浙东沿岸,其湾内风浪较小、海涂宽阔,是浙江省重要的渔业资源生产基地。近年来,人类活动增加和经济发展对该海域生态环境造成了一定的影响。浮游植物作为海洋最主要的初级生产者,在生态系统物质循环、能量流动过程中起着至关重要的作用。虽然往年也有三门湾浮游植物的调查(刘镇盛等,2003; 朱根海等,2010; 刘晶晶等,2015),但往往局限于单一季节,且缺乏环境因子对其时空变化影响的研究。本文以三门湾网采浮游植物及理化数据为基础,首次整合四季数据,分季节、分区域研究影响浮游植物群落的主要环境因子,并对比历史资料,分析三门湾浮游植物群落变化趋势,为深入了解环境变化对该海域浮游植物的影响提供基础资料,同时也为三门湾生态环境长期监测提供历史数据积累。虽然网采方法会遗漏个体较小的浮游植物物种,但考虑到20 世纪80 年代以来的几次调查数据均由网采获得,为了历史数据的可比性,本文仍沿用网采的方法。另一方面,近岸海域主要优势种仍是圆筛藻(Coscinodiscus)等大型硅藻及角毛藻(Chaetoceros)、骨条藻(Skeletonema)等链状硅藻(浙江沿岸生态环境及海湾环境容量编委会,2015),网采方法在一定程度上也能反映海域浮游植物群落结构变化。
1 研究区域和方法 1.1 研究区域和站位布设三门湾调查共设18 个大面站,站位分布见图 1,根据三门湾的地理位置,将其分为湾底Ⅰ(S01 -S06)、湾中部Ⅱ(S07-S12)和湾口Ⅲ(S13-S18)。于2015 年春季(5 月)、夏季(8 月)、秋季(11 月)和2016年冬季(1 月)各采样一次,其中由于海况原因,夏季未采集S08和S11 站位。
 |
| 图 1 三门湾海域调查站位 Fig. 1 Sampling stations in the Sanmen Bay 注:Ⅰ. 湾底海域; Ⅱ. 湾中部海域; Ⅲ. 湾口海域 |
样品采集按照《海洋调查规范》(国家标准化委员会,2007)采用浅Ⅲ型(网孔直径76μm)浮游生物网自底至表作垂直拖网采集,用Hydro-bios 流量计(HYDRO-BIOS 公司,德国)记录滤水量。样品经中性甲醛(上海凌峰化学试剂有限公司,中国)固定,静置24h 并浓缩后用ZEISS Scope A1 显微镜(Carl Zeiss 公司,德国)进行观察、计数和鉴定。浮游植物物种鉴定参考《中国海域甲藻》和《中国海域硅藻图谱》等书籍(中国科学院中国孢子植物志编辑委员会,1980; 金德祥,1982; 杨世民等,2006,2014)。
理化数据均通过采集表层(0.5m)水样测定得到,其中温度、盐度和水深采用CTD 传感器(HACH Sea bird 公司,美国)测定。其余理化因子测定均按照《海洋监测规范》(国家标准化委员会,2007),其中溶解无机氮(dissolved inorganic nitrogen,DIN)、溶解无机磷(dissolved inorganic phosphorus,DIP)和硅酸盐(dissolved silicate,DSi)使用7230G 分光光度计(上海精科仪器有限公司,中国)采用分光光度法,悬浮物浓度采用重量法。
1.3 数据处理用PEIMER 6.0 软件计算浮游植物种类数S、Margalef 丰富度指数d、Pielou 均匀度指数J(Pielou,1969)和Shannon-Winner 多样性指数H′(Magurran,1988),同时对浮游植物物种丰度进行lg(x+1)转换后,建立Bay-Curtis 相似性聚类分析。浮游植物群落比较采用ANOSIM 分析。用SPSS 20.0 软件对三门湾不同海域环境因子作非参数检验(K-W 检验),比较各组数据间差异。用Canoco4.5 软件选各季优势度大于0.01的17 种浮游植物和环境因子进行CCA 排序。站位和丰度分布图采用Surfer 11 绘制,柱状图采用SigmaPlot 10.0 软件绘制。
优势度(Y)计算公式为:
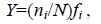
式中,ni 为第i 种的个体数,N 为总个体数,fi 为第i 种在各样品中出现的频率,若某物种的Y≥0.02 时,则定为优势种。
富营养化指数计算公式为:
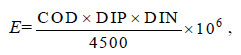
式中,COD、DIN和DIP 单位均为mg/L,E≥1 时,即可认为水体呈富营养化,且E 与水体富营养化程度呈正比(邹景忠等,1983)。
2 结果与分析 2.1 种类组成调查共鉴定出浮游植物8 门87 属199 种(含变种、变型和未定名种),其中硅藻种类数最多,共51 属149 种(占74.9%); 甲藻次之,共18 属27 种(占13.6%);绿藻6 属7 种(占3.5%); 裸藻5 属6 种(占3.0%); 隐藻3 属4 种(占2.0%); 蓝藻2 属3 种(占1.5%); 金藻1 属2 种(占1.0%); 定鞭藻1 属1 种(占0.5%)。浮游植物种类季节分布呈冬季(128 种)>秋季(116 种)>夏季(104 种)>春季(98 种),四季种类组成以硅藻和甲藻为主(表 1)。
表 1 三门湾四季浮游植物种类组成
Tab. 1 The composition of phytoplankton species in Sanmen Bay in different seasons
| 门类 | 春 | 夏 | 秋 | ||
| 硅藻门 | Bacillariophyta | 30 属 69 种 | 35 属 74 种 | 41 属 95 种 | |
| 甲藻门 | Pyrrophyta | 11 属 15 种 | 13 属 18 种 | 8 属 15 种 | |
| 绿藻门 | Chlorophyta | 4 属 4 种 | 3 属 3 种 | 1 属 1 种 | |
| 裸藻门 | Euglenophyta | 4 属 5 种 | 3 属 4 种 | 1 属 1 种 | |
| 蓝藻门 | Cyanophyta | 2 属 2 种 | - | 1 属 1 种 | |
| 隐藻门 | Cryptophyta | 2 属 2 种 | 2 属 3 种 | 1 属 1 种 | |
| 金藻门 | Chrysophyta | - | 1 属 1 种 | 1 属 2 种 | |
| 定鞭藻门 | Prymnesiophyta | 1 属 1 种 | 1 属 1 种 | - | |
| 总计 | Total | 54 属 98 种 | 58 属 104 种 | 54 属 116 种 | |
| 注: “-”表示未检出 | |||||
调查海域浮游植物年均丰度为1632.04 ×104cell/m3,且季节差异明显。夏季丰度最高(平均丰度为5548.90×104cell/m3),秋、冬季次之(分别为780.77×104cell/m3和580.64×104cell/m3),春季最低(仅为53.06×104cell/m3)。
丰度空间分布上,夏季湾口最高,次高值区位于湾底; 秋、冬两季浮游植物丰度高值区逐渐向湾内移动,秋季高值区为湾口近湾中部,冬季为湾底和湾中部,湾口丰度最低; 春季浮游植物细胞丰度分布与冬季相似,高值区位于湾中部靠近湾底(图 2)。
 |
| 图 2 三门湾四季浮游植物丰度分布(×104cell/m3) Fig. 2 Distribution of phytoplankton abundance in Sanmen Bay in different seasons(×104cell/m3) 注: a. 春季; b. 夏季; c. 秋季; d. 冬季 |
本次调查共有优势种8 属13 种。其中琼氏圆筛藻(Coscinodiscus jonesianus)为全年优势种; 蛇目圆筛藻(C. argus)、虹彩圆筛藻(C. oculus-iridis)和伏氏海毛藻(Thalassiothrix frauenfeldii)为春、秋、冬三季优势种; 骨条藻(Skeletonema spp.)为夏、秋、冬三季优势种。夏季,链状硅藻占绝对优势,如骨条藻(Y=0.81)和拟旋链角毛藻(Chaetoceros pseudocurvisetus)(Y=0.07)等(表 2)。
表 2 三门湾浮游植物优势种平均丰度(×104cell/m3)和优势度(Y)
Tab. 2 Dominance and abundance of dominant phytoplankton species in Sanmen Bay
| 优势种 | 春 | 夏 | 秋 | 冬 | ||||
| 丰度 | Y | 丰度 | Y | 丰度 | Y | 丰度 | Y | |
| 中华盒形藻(Biddulphia sinensis) | 6.91 | 0.12 | - | - | - | - | - | - |
| 并基角毛藻(Chaetoceros decipiens) | - | - | - | - | 36.75 | 0.05 | - | - |
| 劳氏角毛藻(C. lorenzianus) | - | - | - | - | 35.61 | 0.04 | - | - |
| 拟旋链角毛藻(C. pseudocurvisetus) | - | - | 376.08 | 0.07 | - | - | - | - |
| 蛇目圆筛藻(Coscinodiscus argus) | 3.85 | 0.07 | - | - | 48.16 | 0.06 | 46.59 | 0.08 |
| 星脐圆筛藻(C. asteromphalus) | 2.12 | 0.04 | - | - | - | - | - | - |
| 琼氏圆筛藻(C. jonesianus) | 22.56 | 0.43 | 87.1 | 0.02 | 95.98 | 0.12 | 76.02 | 0.13 |
| 虹彩圆筛藻(C. oculus-iridis) | 4.59 | 0.09 | - | - | 31.3 | 0.04 | 19.68 | 0.03 |
| 楔形半盘藻(Hemidiscus cuneiformis) | - | - | - | - | 22.14 | 0.03 | 25.92 | 0.04 |
| 弯菱形藻(Nitzschia sigma) | 3.66 | 0.07 | - | - | - | - | - | - |
| 尖刺伪菱形藻(Pseudo-nitzschia pungens) | - | - | - | - | 125.04 | 0.16 | 28.29 | 0.05 |
| 骨条藻(Skeletonema spp.) | - | - | 4513.03 | 0.81 | 215.39 | 0.26 | 245.33 | 0.38 |
| 伏氏海毛藻(Thalassiothrix frauenfeldii) | 1.41 | 0.03 | - | - | 35.77 | 0.05 | 15.9 | 0.03 |
| 注: “-”表示非优势种 | ||||||||
浮游植物d、H′和J 均存在显著的季节差异(P<0.05),其中d和H′差异极显著(P<0.01)。夏季d、H′和J(分别为3.60、0.48和1.69)明显低于春(分别为4.21、0.61和1.98)、秋(4.63、0.66和2.44)、冬季(4.54、0.66和2.38)。
春季H′和J 均呈现自湾中部向湾底及湾口降低的趋势,而d 则自湾底向湾口逐渐增高; 夏季d、H′和J 湾底最高,湾中部和湾口相对较低,冬季分布趋势与夏季相反; 秋季H′和J 分布趋势与夏季相反,d 呈现自湾中部向湾底及湾口逐渐降低的趋势(图 3)。
 |
| 图 3 三门湾四季网采浮游植物多样性指数(H′)、丰富度指数(d)和均匀度指数(J)分区比较 Fig. 3 Seasonal variations of H′,d and J in different sections of Sanmen Bay 注: a. 多样性指数H′; b. 丰富度指数d; c. 均匀度指数J。不同季节间H′和J 比较采用K-W 检验,不同小写字母表示全区季节间d、H′和J 存在显著差异(P<0.05) |
调查期间透明度、温度、盐度、悬浮物浓度、DSi、DIP和DIN 均存在显著的季节性变化(P<0.05)。冬季透明度、温度和盐度明显低于其余三季,春、夏季温度和盐度较高,秋季透明度最高; 悬浮物浓度变化趋势与透明度相反; 春、夏两季营养盐(DSi、DIP和DIN)低于秋、冬季。
各季环境因子区域分布上,春、秋两季透明度呈湾底向湾口逐渐降低的趋势,夏、冬两季湾中部最低,湾底和湾口较高; 悬浮物浓度分布与透明度相反;春、夏季温度自湾底向湾口降低,秋、冬季则相反; 四季盐度均呈湾底向湾口升高的趋势; 春、夏季DSi 湾中部最低,湾底及湾口较高,秋、冬季自湾底向湾口降低; 夏、秋、冬三季DIN 湾底最低,向湾口升高,春季分布趋势相反; 春季DIP 高值区出现在湾中部,湾底和湾口较低,秋季分布与春季相反,夏、冬季均自湾底向湾口降低(图 4)。
 |
| 图 4 三门湾四季环境因子分区比较 Fig. 4 Seasonal variation of environmental factors in different sections of Sanmen Bay 注: a: 透明度; b: 温度(℃); c: 悬浮物浓度(mg/L); d: 盐度; e: 溶解硅酸盐(mg/L); f: 溶解无机氮(mg/L); g: 溶解无机磷(mg/L); 同行不同小写字母表示不同区域差异显著(P<0.05) |
根据聚类分析结果(图 5),可将浮游植物分为春、夏、秋、冬季群落,单因素ANOSIM 也表明,浮游植物群落季节差异显著(R = 0.99,P = 0.001)。表 3 相似性分析结果表明各季湾底(S01-S06)及湾口(S13-S18)均存在显著差异,其中春、夏、冬季湾底和湾中部(S07-S12)也存在显著差异(P<0.05)。
 |
| 图 5 三门湾各站位浮游植物样品聚类分析 Fig. 5 Clusters analysis on different phytoplankton samples in Sanmen Bay 注: “1,5,8,11”代表采集的相应月份 |
表 3 三门湾不同区域浮游植物单因素相似性分析
Tab. 3 A one-way analysis of similarity for phytoplankton community in different sections in Sanmen Bay
| 区域 | 春 | 夏 | 秋 | 冬 | ||||
| R | P | R | P | R | P | R | P | |
| 湾底 vs.湾中部 | 0.15 | 0.082 | 0.27 | 0.052 | 0.05 | 0.604 | 0.06 | 0.288 |
| 湾底 vs.湾口 | 0.66 | 0.002 | 0.62 | 0.002 | 0.3 | 0.015 | 0.49 | 0.002 |
| 湾中部 vs.湾口 | 0.38 | 0.015 | 0.46 | 0.024 | 0.17 | 0.139 | 0.26 | 0.004 |
| 注: R 代表相似性系数, P 代表显著性 | ||||||||
根据浮游植物的出现比例和丰度,选取各季优势度Y>0.01 的17 种用于CCA 排序(表 4)。蒙特卡罗检验(Monte Carlo test)检验表明,第一轴和全部轴均呈极显著差异(P = 0.002),故CCA 排序结果可信。
表 4 CCA 排序中浮游植物种类代码
Tab. 4 The codes of phytoplankton species for CCA ordination
| 编号 | 种类 |
| sp.1 | 中华盒形藻(Bidduiphia sinensis) |
| sp.2 | 并基角毛藻(Chaetoceros decipiens) |
| sp.3 | 劳氏角毛藻(C. lorenzianus) |
| sp.4 | 拟旋链角毛藻(C. pseudocurvisetus) |
| sp.5 | 蛇目圆筛藻(Coscinodiscus argus) |
| sp.6 | 星脐圆筛藻(C. asteromphalus) |
| sp.7 | 琼氏圆筛藻(C. jonesianus) |
| sp.8 | 虹彩圆筛藻(C. oculus-iridis) |
| sp.9 | 苏里圆筛藻(C. thorii) |
| sp.10 | 布氏双尾藻(Ditylum brightwellii) |
| sp.11 | 楔形半盘藻(Hemidiscus cuneiformis) |
| sp.12 | 弯菱形藻(Nitzschia sigma) |
| sp.13 | 尖刺伪菱形藻(Pseudo-nitzschia pungens) |
| sp.14 | 骨条藻(Skeletonema spp. ) |
| sp.15 | 菱形海线藻(Thalassionema nitzschioides) |
| sp.16 | 圆海链藻(Thalassiosira rotula) |
| sp.17 | 伏氏海毛藻(Thalassiothrix frauenfeldii) |
CCA 分析图(图 6)中的7 个环境因子可解释浮游植物总变量的55.2%。第1 轴和第2 轴的特征值分别为0.205和0.119,并各自解释了31.5%和18.3%的物种变量。第1 轴和第2 轴的物种-环境相关系数分别为0.876和0.804,表明这7 个环境因子与18 种浮游植物种类相关性较好。温度、DSi、DIN和盐度是影响浮游植物群落的主要环境因子。结合CCA 排序轴与各环节因子的相关性大小(表 5),第1 轴主要受温度(0.7363)和DIN(-0.6519)等因子的影响,第2 轴主要受盐度(0.6648)和DSi(0.5973)等因子的影响。劳氏角毛藻(C. lorenzianus)(sp.3)和拟旋链角毛藻(sp.4)与温度正相关,尖刺伪菱形藻(Pseudo-nitzschia pungens)(sp.13)、伏氏海毛藻(sp.17)和骨条藻(sp.14)等赤潮藻种与营养盐(DIP、DIN和DSi)密切正相关。
 |
| 图 6 主要浮游植物种类与环境因子之间的CCA 排序图 Fig. 6 CCA ordination of main phytoplankton species with environmental variables |
表 5 CCA 分析排序轴与环境因子的相关性系数
Tab. 5 Axes-environmental correlation coefficient of CCA
| 环境因子 | 轴 1 | 轴 2 |
| 透明度 | 0.412 | -0.0854 |
| 温度 | 0.7363 | 0.3061 |
| 盐度 | 0.1243 | 0.6648 |
| 悬浮物 | -0.3757 | -0.1924 |
| 硅酸盐 | -0.378 | -0.5973 |
| 溶解无机磷 | -0.4109 | -0.4484 |
| 溶解无机氮 | -0.6519 | -0.3493 |
近30 年来,三门湾海域浮游植物年均丰度呈先下降后升高的规律,其中20 世纪80 年代冬季浮游植物丰度明显高于同期其余季节,比对发现仅该季出现赤潮藻种中肋骨条藻(S. costatum),因此该季可能发生该藻种赤潮。若排除该次调查数据,则浮游植物年均丰度总体呈上升趋势(图 7a)。20 世纪80 年代浮游植物年均丰度为157.47×104cell/m3(不包括冬季),本次调查年均丰度达1740.84×104cell/m3,增加11 倍。此外,不同时期三门湾海域浮游植物丰度亦呈现季节变化,20 世纪80 年代冬季出现峰值,在2006-2007 年和本次调查中则夏季最高(图 7b)。此外,比较浮游植物不同年份同季丰度得到,其变化趋势与年均丰度变化趋势总体一致,但2006-2007 年春季和20 世纪80 年代冬季数据高于本次调查。
 |
| 图 7 不同年份三门湾浮游植物丰度及其季节变化 Fig. 7 Seasonal changes in net-phytoplankton abundance in different years in Sanmen Bay 注: a. 不同年份三门湾浮游植物丰度变化; b. 不同年份三门湾浮游植物丰度的季节变化; “*”表示有赤潮发生; 图b 中由于查阅到1982年及1987 年两次秋季采样数据,因此20 世纪80 年代秋季浮游植物丰度有两组柱状数据,对应年份已作标注 |
由表 6 可以看出,三门湾海域浮游植物主要种类以硅藻为主,甲藻次之。赤潮藻种种类数和优势度均显著上升,本次调查优势种中赤潮种种类数达5 属6种,包括中华盒形藻(Biddulphia sinensis)、伏氏海毛藻、拟旋链角毛藻、劳氏角毛藻、骨条藻和尖刺伪菱形藻,为历年最高。骨条藻、旋链角毛藻(C. curvisetus)和尖刺伪菱形藻等链状嗜氮性硅藻的丰度和优势度也呈上升趋势,其中尖刺伪菱形藻自21 世纪以来,在各季优势种中出现频率增加。此外,该海域浮游植物自20 世纪80 年代以来主要种类由布氏双尾藻(Ditylum brightwellii)和豪猪棘冠藻(Corethron hystrix)等大型硅藻向劳氏角毛藻、并基角毛藻(C. decipiens)和尖刺伪菱形藻等链状硅藻演替(本次调查主要种类组成中链状硅藻种类数为历年最高,同时除春季外,夏、秋、冬三季主要种类中均出现链状硅藻)。另外,如楔形半盘藻(Hemidiscus cuneiformis)和拟旋链角毛藻等部分暖水种兴起,且本次冬季调查优势种首次出现了楔形半盘藻。
表 6 不同年份三门湾海域浮游植物主要种类
Tab. 6 The main species of net-phytoplankton in different years in Sanmen Bay
| 年份 | 季节 | 主要种类 | 参考文献 |
| 1981 | 冬 | 布氏双尾藻、中肋骨条藻*、菱形藻、角藻、小环藻(Cyclotella spp.) | (中国海湾志编纂委员会,1992) |
| 1982 | 春 | 圆筛藻、布氏双尾藻、菱形藻、豪猪棘冠藻 | |
| 1982 | 夏 | 圆筛藻、布氏双尾藻、菱形藻、根管藻(Rhizosolenia spp.)#、角毛藻(Chaetoceros spp.) | |
| 1982 | 秋 | 圆筛藻、菱形藻、角藻、小环藻 | |
| 1987 | 秋 | 圆筛藻(Coscinodiscus spp.)、劳氏角毛藻*,#、中肋骨条藻*、布氏双尾藻、尖刺伪菱形藻*、甲藻 | (浙江省海岸带海洋生物资源调查报告海洋生物资源专业组,1988) |
| 2002—2003 | 春 | 具齿原甲藻(Prorocentrum dentatum)*、中肋骨条藻* | (宁修仁等,2005) |
| 2002—2003 | 夏 | 中肋骨条藻*、拟旋链角毛藻#、尖刺伪菱形藻* | |
| 2002—2003 | 秋 | 中肋骨条藻*、尖刺伪菱形藻* | |
| 2002—2003 | 冬 | 中肋骨条藻*、布氏双尾藻 | |
| 2006 | 秋 | 劳氏角毛藻*,#、星脐圆筛藻(Coscinodiscus asteromphalus)、琼氏圆筛藻 | (浙江沿岸生态环境及海湾环境容量编委会,2015) |
| 2006 | 冬 | 星脐圆筛藻、中心圆筛藻(Coscinodiscus centralis)、辐射圆筛藻(Coscinodiscus radiatus)、琼氏圆筛藻、中肋骨条藻* | |
| 2007 | 春 | 蛇目圆筛藻、星脐圆筛藻、中心圆筛藻、琼氏圆筛藻、辐射圆筛藻、洛伦菱形藻(Nitzschia lorenziana)、美丽菱形藻(Nitzschia puicherrima)、长海毛藻(Thalassiothrix longissima) | |
| 2007 | 夏 | 拟旋链海毛藻*,#、琼氏圆筛藻、辐射圆筛藻、中肋骨条藻*、纺锤角藻(Ceratium fusus)*、 大角角藻(Ceratium macroceros)*,# | |
| 2015 | 春 | 中华盒形藻*、蛇目圆筛藻、星脐圆筛藻、琼氏圆筛藻、虹彩圆筛藻、弯菱形藻(Nitzschia sigma)、伏氏海毛藻 | 本调查 |
| 2015 | 夏 | 拟旋链角毛藻*,#、琼氏圆筛藻、骨条藻* | |
| 2015 | 秋 | 并基角毛藻、劳氏角毛藻*,#、蛇目圆筛藻、琼氏圆筛藻、虹彩圆筛藻、楔形半盘藻#、尖刺伪菱形藻*、骨条藻*、伏氏海毛藻 | |
| 2016 | 冬 | 蛇目圆筛藻、琼氏圆筛藻、虹彩圆筛藻、楔形半盘藻#、尖刺伪菱形藻*、骨条藻*、伏氏海毛藻 | |
| 注: “*”表示赤潮藻种; “#”表示暖水种; 2002-2003 年为水样数据 | |||
三门湾海域长期处于富营养化状态(E>1),且日趋严重,至本次调查达6.57,属于中度富营养化(3<E≤9)。近30 年来,营养盐含量和氮磷比(N/P)总体也呈上升趋势,其中与20 世纪80 年代相比,本次调查的DIN年均含量增加近3 倍,N/P 增加2 倍(表 7、图 8a-8c)。对不同年份数据进行季节比对,得到20 世纪80 年代和2002-2003 年两次调查冬季富营养化指数和营养盐含量较同期春、夏、秋季高,其中20 世纪80 年代冬季DIP 含量和富营养化指数明显高于其余三季(图 8d)。
表 7 不同年份三门湾浮游植物氮磷比(N/P)、营养盐浓度和富营养化指数(E)
Tab. 7 N/P,nutrient concentration,and eutrophication index in different years in Sanmen Bay
| 年份 | 营养盐/(mg/L) | E | N/P | 参考文献 | |
| DIN | DIP | ||||
| 20 世纪 80 年代 | 0.26 | 0.032 | 4.59 | 17.8 | (中国海湾志编纂委员会,1992) |
| 2002—2003 | 0.49 | 0.026 | 2.35 | 42.35 | (宁修仁,2005) |
| 2006—2007 | 0.62 | 0.033 | 4.18 | 41.9 | (浙江沿岸生态环境及海湾环境容量编委会等,2015) |
| 2015—2016 | 0.63 | 0.036 | 6.57 | 39.18 | 本调查 |
| 注: DIN 表示溶解无机氮, DIP 表示溶解我机磷; 富营养化指数均采用营养指数法计算 | |||||
 |
| 图 8 不同年份三门湾营养盐含量、氮磷比和富营养化指数的季节变化 Fig. 8 Seasonal changes of N/P,nutrient concentration and eutrophication index in different years in Sanmen Bay 注: a: 溶解无机氮(mg/L); b: 溶解无机磷(mg/L); c: 氮磷比; d: 富营养化指数 |
三门湾属半封闭港湾,湾口开阔,毗连东海。该海域主要受到陆源入海径流、江浙沿岸流和台湾暖流的共同影响(刘镇盛等,2006)。因此,湾内海域中的浮游植物既有径流入海带来的淡水种和半咸水种,如单角盘星藻(Pediastrum simplex)、小席藻(Phormidium tenue)、颗粒直链藻极狭变种(Melosira granulata var.angustissima)和小球藻(Chlorella spp.)等,又有江浙沿岸流带来的近岸低盐种和河口土著种,如琼氏圆筛藻、布氏双尾藻、肘状针杆藻(Synedra ulna)和微小原甲藻(Prorocentrum minimum)等,该类群也是春、秋、冬季主要优势种。此外,还有台湾暖流带来的部分暖水种和外海高盐种,如大洋角管藻(Cerataulina pelagica)、拟旋链角毛藻和劳氏角毛藻等,夏季该类群优势度显著上升。
优势种组成上,春季优势种以琼氏圆筛藻为主,组成上均为大型硅藻(表 2),这可能是由于春季营养盐含量较低(图 4e-4g),而小型链状硅藻由于其繁殖速率快,在高营养盐环境下才更具竞争力,因此大型硅藻较链状硅藻更适应于春季水体环境(Wang et al,2009)。夏季水体均温28.8℃,平均盐度26.0,为旋链角毛藻等角毛藻属藻种最适生长环境(温度25-30℃,盐度25-31),同时角毛藻对光照强度和时间敏感性较强,较长的光照时间和强度有利于角毛藻的大量繁殖(Göksan et al,2003; 王朝晖等,2006)。此外,高温也有利于促进骨条藻的生长(张晓东,2012)。因此夏季骨条藻(Y=0.81)和拟旋链角毛藻(Y=0.07)等链状硅藻替代大型硅藻成为三门湾海域主要优势种。CCA分析中骨条藻、劳氏角毛藻和拟旋链角毛藻与温度也呈正相关(图 6)。其中,骨条藻的大量生长导致夏季优势种数量、H′、d和J 均低于其他三季(表 2,图 3)。秋、冬季浮游植物群落组成相似度较高(图 5)。优势种组成上,两季共有赤潮藻种5 种(琼氏圆筛藻、伏氏海毛藻、骨条藻、尖刺伪菱形藻和劳氏角毛藻),为全年最高,其中劳氏角毛藻是秋季优势种(表 2),这可能由于秋、冬季海域受高营养盐的江浙沿岸流控制(中国海湾志编纂委员会,1992),与此前大量研究表明的赤潮藻种生长与营养盐含量上升密切相关的结论一致(韩秀荣等,2003; 叶属峰等,2003; 陈月红等,2015)。CCA 分析中尖刺伪菱形藻、伏氏海毛藻和骨条藻等赤潮藻种与营养盐的正相关也证明了该结论(图 6)。此外,秋季温度相对较高,角毛藻属如并基角毛藻等仍能大量生长,因此该种仍为优势种。
3.2 浮游植物的区域比较ANOSIM和聚类分析(表 3、图 5)表明各季湾底及湾口浮游植物群落均存在显著差异,这与水温、盐度和透明度等环境因子的变化有着密切关系。
由图 4a 可知,春季湾底透明度高,真光层较厚,适应浮游植物生长,因此春季湾底丰度最高(图 2)。夏季台湾暖流入侵加剧(中国海湾志编纂委员会,1992),因此夏季湾口海域聚集台湾暖流携带的大量外海暖水种,丰度上升。同时,高透明度、寡营养盐的外海水入侵也使得湾口处透明度较高而悬浮物浓度和营养盐含量较低(图 4a、图 4c、图 4e-4g)。此外,该季湾底水体稳定,透明度和水温较高,加之丰水期陆源冲淡水的营养盐补充,使得湾底不仅适应于土著种的生长,也为部分暖水种提供了良好的生长条件,形成丰度峰值区(图 2),同时群落参数(H′、d和J)也高于其余两区域(图 3)。秋季台湾暖流势力减退(中国海湾志编纂委员会,1992),湾口由于内湾水和海水交换引起的水体扰动导致悬浮物浓度上升而透明度下降(图 4a、图 4c),大部分藻类生长受到限制(Song et al,2008),因此秋季湾口海域丰度较夏季明显降低。而湾中部及湾口近内湾站位(S13-S15)营养盐含量和透明度相对较高,水体较温暖(图 4a-4b;图 4e-4f),部分暖水种仍能继续生长,形成丰度高值区。冬季受高营养盐沿岸流控制,悬浮物浓度上升,透明度下降,使得适应浑浊水体的琼氏圆筛藻、蛇目圆筛藻和对光照适应性较强的骨条藻等种类大量生长(张晓东,2012),而其他藻类生长受到不同程度限制,优势度下降。冬季湾底海域水体相对稳定,成为丰度峰值区,湾口水动力较强,加之温度和透明度较低,不适于浮游植物的生长,丰度最低(图 2)。
3.3 浮游植物对环境变化的响应三门湾海域浮游植物丰度有明显的年际和季节变化,总体呈增加趋势。20 世纪80 年代冬季发生赤潮,导致该次采样冬季丰度明显高于同期其余三季,且为历年同季最高。在2006-2007 年及本次调查中,夏季浮游植物丰度最高,这可能是由于夏季属于丰水期,丰富的降水使得该海域得到高营养盐地表径流大量补充,加之温度相对较高,更利于浮游植物的生长(图 7),这与吴玉霖等(2004)对胶州湾浮游植物调查结果一致。由表 7 可知,近30 年三门湾海域营养盐含量和富营养化指数均呈上升趋势,为浮游植物的生长提供了充足的物质基础,促使浮游植物大量生长,使得本次调查浮游植物年均丰度较20 世纪80 年代增加11 倍。此外,由于2006-2007 年春季N、P 营养盐含量高于不同时期同季调查,这可能也导致该次调查浮游植物丰度同季最高(图 7、图 8)。另外,不同年度四季调查中秋、冬季上升显著,这可能是由于全球气候变化使得海水温度升高,有利于浮游植物生长(孙晓霞等,2011)。其中由表 6可知,1987年秋季赤潮藻种种类数较1982 及2006 年同季调查明显增多(劳氏角毛藻、中肋骨条藻和尖刺伪菱形藻),赤潮种的大量生长可能导致该次调查浮游植物丰度为历年调查最高(略高于本次调查)。
由表 6 可知,近30 年三门湾海域硅藻组成上,链状硅藻替代部分大型硅藻成为优势种,这可能由于该海域营养盐含量升高,使得繁殖速率较快的链状硅藻大量生长(Zhou et al,2008; Wang et al,2009)。加之三门湾海域N/P 逐年升高,链状硅藻由于个体较小,其细胞组成对P 需求较少,相对于大型硅藻更适应于高N/P 环境(沈志良,2002; Glibert et al,2011)。而链状硅藻的优势度上升会替代角藻和原甲藻等大型甲藻成为优势种,这可能是本次调查优势种中未出现甲藻的原因之一,同时调查时期未处于甲藻优势期或者采样站位未位于甲藻优势区也有可能导致甲藻在本次调查未成为海域优势种,具体原因还有待进一步论证。
此外,近30 年三门湾海域暖水种兴起,该趋势可能是浮游植物对气候变暖、海水温度升高的响应。李家星等(2012)对东海表层水温(sea surface temperature,SST)年际变化的分析结果也表明近52 年来东海水温呈升高趋势。本次调查中劳氏角毛藻、楔形半盘藻和拟旋链角毛藻等暖水种的大量生长也从一定程度上反应了气候变暖和SST 升高对浮游植物群落结构的影响,其中冬季调查优势种首次出现暖水种。同时嗜氮链状硅藻和暖水种的兴起也可能是本调查浮游植物年均丰度上升的原因之一(刘东艳,2004)。另外,三门湾海域由于湾内水体交换较缓慢和周边城镇工业生活污水入海等原因,长期处于富营养化状态(E>1)(邹景忠等,1983),加之受到沿岸经济开发和人口增加等人为影响,富营养化程度日趋严重(表 7),导致海域中赤潮藻种种类数和优势度明显上升(吴在兴等,2014)。由此可见,海域富营养化和水温上升可能是浮游植物丰度上升和优势种组成改变的主要驱动因子。
4 结语2015-2016 年四季调查,共检出浮游植物8 门87 属199 种,硅藻为主(51 属149 种),甲藻次之(18属27 种)。优势种8 属13 种,其中琼氏圆筛藻为全年优势种,夏季骨条藻和角毛藻等链状硅藻占绝对优势。全年该海域浮游植物年平均丰度为1632.04×104cell/m3 且季节差异明显。夏季丰度最高,达5548.90×104cell/m3; 春季最低,仅53.06×104cell/m3。浮游植物群落组成、多样性指数(H′、J和d)和理化因子(透明度、温度、盐度、悬浮物浓度、DSi、DIP和DIN)等均存在显著的季节变化和区域差异(尤其是湾底和湾口)。近30 年来,三门湾富营养化程度和浮游植物丰度总体呈上升趋势。主要种类组成上,硅藻仍为该海域绝对优势种,其中部分大型硅藻逐步被链状硅藻所替代,且赤潮藻种和嗜氮性链状硅藻种类数和占比逐年上升。
致谢 感谢刘小涯、于培松、张海峰、蔡小霞、胡佶等协助采样并提供理化数据。
参考文献
| 王朝晖, 齐玉藻, 陈菊芳, 等, 2006. 大亚湾角毛藻细胞数量波动及其与环境因子关系的多元分析. 生态学报, 26 (4) : 1096–1102 |
| 中国科学院中国孢子植物志编辑委员会, 1980. 中国海藻志第五卷(硅藻门). 北京: 科技出版社, |
| 中国海湾志编纂委员会, 1992. 三门湾. 见:中国海湾志编纂委员会主编. 中国海湾志第五分册(上海市和浙江省北部海湾). 北京:海洋出版社, 234-308 |
| 叶属峰, 黄秀清, 2003. 东海赤潮高发区营养盐时空分布特征及其与赤潮的关系. 海洋环境科学, 22 (2) : 2416–2425 |
| 宁修仁, 2005. 三门湾海域环境质量现状及其年际变化. 见:宁修仁主编. 乐清湾、三门湾养殖生态和养殖容量研究与评价. 北京:海洋出版社, 120-160 |
| 朱根海, 陈立红, 钱键, 2010. 三门核电站附近海域浮游植物季节变化研究. 环境科学与技术, 33 (12F) : 36–45 |
| 刘东艳, 2004. 胶州湾浮游植物与沉积物中硅藻群落结构演替的研究. 青岛:中国海洋大学博士学位论文, 1-127 |
| 刘晶晶, 江志兵, 陈悦, 等, 2015. 三门湾春季网采浮游植物群落特征. 海洋学研究, 33 (1) : 74–80 |
| 刘镇盛, 王春生, 张志南, 2006. 三门湾浮游动物的季节变动及微型浮游动物摄食影响. 生态学报, 26 (12) : 3931–3941 |
| 刘镇盛, 蔡昱明, 刘子琳, 等, 2003. 三门湾秋季浮游植物现存量和初级生产力. 东海海洋, 21 (2) : 30–36 |
| 孙晓霞, 孙松, 吴玉霖, 等, 2011. 胶州湾网采浮游植物群落结构的长期变化. 海洋与湖沼, 42 (5) : 639–646 |
| 李家星, 魏皓, 罗晓凡, 2012. 东海陆架表层水温年际变化时空特征分析. 海洋与湖沼, 43 (3) : 624–634 |
| 杨世民, 李瑞香, 董树刚, 2014. 中国海域甲藻Ⅰ(原甲藻目、鳍藻目). 北京: 海洋出版社, 155 |
| 杨世民, 董树刚, 2006. 中国海域常见浮游硅藻图谱. 青岛: 中国海洋大学出版社, 258 |
| 吴玉霖, 孙松, 张永山, 等, 2004. 胶州湾浮游植物数量长期动态变化的研究. 海洋与湖沼, 35 (6) : 518–523 |
| 吴在兴, 俞志明, 宋秀贤, 等, 2014. 基于水质状态和生态响应的综合富营养化评价模型-以山东半岛典型海域富营养化评价为例. 海洋与湖沼, 45 (1) : 20–31 |
| 邹景忠, 董丽萍, 秦保平, 1983. 渤海湾富营养化和赤潮问题的初步探讨. 海洋环境科学, 2 (3) : 41–54 |
| 沈志良, 2002. 胶州湾营养盐结构的长期变化及其对生态环境的影响. 海洋与湖沼, 33 (3) : 322–331 |
| 张晓东, 2012. 厦门港骨条藻属Skeletonema (Bacillariophyta)物种多样性及周年变化的研究. 青岛:中国科学院研究生院(海洋研究所)硕士学位论文, 22-45 |
| 陈月红, 石晓勇, 韩秀荣, 等, 2015. 闽浙近岸海域赤潮暴发前后海水营养盐特征分析. 海洋通报, 34 (5) : 568–576 |
| 国家标准化委员会, 2008. GB/T 12763.6-2007海洋调查规范第6部分:海洋生物调查. 北京: 中国标准出版社, 30-33 |
| 国家标准化委员会, 2008. GB 17378.4-2007海洋监测规范第4部分:海水分析. 北京: 中国标准出版社, 57-121 |
| 金德祥, 1982. 中国海洋底栖硅藻类. 北京: 海洋出版社, 237 |
| 浙江沿岸生态环境及海湾环境容量编委会, 2015. 三门湾海洋环境质量变化趋势综合评价. 见:寿鹿, 曾江宁主编. |
| 浙江沿岸生态环境及海湾环境容量. 北京:海洋出版社, 136-228 |
| 韩秀荣, 王修林, 孙霞, 等, 2003. 东海近海海域营养盐分布特征及其与赤潮发生关系的初步研究. 应用生态学报, 14 (7) : 1097–1101 |
| Glibert P M, Burkholder J M, 2011. Harmful algal blooms and eutrophication:"strategies" for nutrient uptake and growth outside the Redfield comfort zone. Chinese Journal of Oceanology and Limnology, 29 (4) : 724–738 DOI:10.1007/s00343-011-0502-z |
| Göksan T, Durmaz Y, Gökpınar Ş, 2003. Effects of light path lengths and initial culture density on the cultivation of Chaetoceros muelleri (Lemmermann, 1898). Aquaculture, 217 (1-4) : 431–436 DOI:10.1016/S0044-8486(01)00854-7 |
| Magurran A E, 1988. Ecological Diversity and Its Measurement. New Jersey, USA: Princeton University Press, 179 |
| Pielou E C, 1969. An introduction to mathematical ecology. BioScience, 78 (1) : 7–12 |
| Song S Q, Sun J, Luan Q S, et al, 2008. Size-fractionated phytoplankton biomass in autumn of Changjiang (Yangtze)River Estuary and its adjacent waters after the three Gorges Dam construction. Chinese Journal of Oceanology and Limnology, 26 (3) : 268–275 DOI:10.1007/s00343-008-0268-0 |
| Wang Z H, Zhao J G, Zhang Y J, et al, 2009. Phytoplankton community structure and environmental parameters in aquaculture areas of Daya Bay, South China Sea. Journal of Environmental Sciences, 21 (9) : 1268–1275 DOI:10.1016/S1001-0742(08)62414-6 |
| Zhou M J, Shen Z L, Yu R C, 2008. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Continental Shelf Research, 28 (12) : 1483–1489 DOI:10.1016/j.csr.2007.02.009 |