 2017, Vol. 48
2017, Vol. 48中国海洋湖沼学会主办。
文章信息
- 李晓静, 周政权, 陈琳琳, 李宝泉, 刘甜甜, 艾冰花, 杨陆飞, 刘博, 王诗帅. 2017.
- LI Xiao-Jing, ZHOU Zheng-Quan, CHEN Lin-Lin, LI Bao-Quan, LIU Tian-Tian, AI Bing-Hua, YANG Lu-Fei, LIU Bo, WANG Shi-Shuai. 2017.
- 渤海湾曹妃甸围填海工程对大型底栖动物群落的影响
- EFFECT OF COASTAL RECLAMATION ON BENTHIC MACROFAUNA IN COASTAL AREA OF CAOFEIDIAN, BOHAI BAY
- 海洋与湖沼, 48(3): 617-627
- Oceanologia et Limnologia Sinica, 48(3): 617-627.
- http://dx.doi.org/10.11693/hyhz20161200292
-
文章历史
- 收稿日期:2016-12-31
- 收修改稿日期:2017-02-14




2. 中国科学院大学 北京 100049;
3. 浙江海洋大学 舟山 316022;
4. 烟台大学 烟台 264003;
5. 莱州市渔政渔港监督管理站 莱州 261400
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Zhejiang Ocean University, Zhoushan 316022, China;
4. Yantai University, Yantai 264003, China;
5. Fishery and Fishing Harbor Administration Station of Laizhou, Laizhou 261400, China
随着沿海经济发展和人口增长, 土地短缺的问题日益突出, 围填海作为解决土地短缺, 拓展发展空间的有效手段, 已被沿海国家广泛采纳。然而, 围填海工程在带来巨大的经济利益的同时, 也对海洋环境和海洋生物等造成了严重的破坏。20世纪中后期, 人们已经意识到围填海的巨大破坏力, 荷兰、日本、美国等发达国家开始进行围填海区域湿地等的恢复研究, 注重经济进步与海岸带可持续利用综合发展(Wang et al, 2014; 李宝泉等, 2016)。
中国大陆海岸线长度约为18000km, 包括海岛在内的总海岸线长度则达到30000km, 中国的围填海工程开始于20世纪中期, 规模持续增大, 围填海几乎遍及海岸线各处。据估计, 到2020年, 中国对围填海造陆面积的需求将超过5880km, 接近过去50年围填海总面积的一半, 其影响不言而喻(Wang et al, 2014)。
曹妃甸海域位于河北省唐山市唐海县南部, 渤海湾东北部, 东经118°28´40″—118°34´0″、北纬38°54´0″—39°05´0″。曹妃甸沙岛北侧与大陆岸线之间发育大片潮间浅滩, 滩地地形破碎复杂, 生境形式多样, 生物资源丰富(索安宁等, 2012)。渤海湾主要的两个围填海区域为曹妃甸和天津滨海新区, 其中曹妃甸围填海工程开始于2004年预计到2020年围填海面积达310km(尹延鸿, 2007); 天津新区始于1994年, 预计围填海面积为2270km(肖庆聪等, 2012)。
大型底栖生物是海洋生态系统中种类最多、生态关系最复杂的生物类群, 在海洋生态系统的物质循环和能量流动中发挥着重要的作用。大型底栖动物活动范围有限, 有些营固着生活, 对逆境的逃避能力较差, 受环境影响更为深刻(李新正, 2011)。索安宁等(2012)对曹妃甸围填海工程的环境回顾性评价表明, 在5类海洋生物群落中底栖动物受影响程度最大, 其中群落密度影响最大, 其次为底栖动物种数、生物量和生物多样性指数(索安宁等, 2012)。Lu等(2002)于1998—2000年对新加坡Sungei Punggol河(榜鹅河)围填海区域生物群落进行了研究, 发现围填海区域的大型底栖动物群落物种数和丰度均明显减少, 而附近区域的大型底栖动物的物种数和丰度均有所上升(Lu et al, 2002)。索安宁等(2015)研究了2004—2010年围填海对曹妃甸海域鱼类群落的影响, 对曹妃甸围填海前中后期的鱼类群落物种丰富度、密度、生物量、生物多样性指数进行了分析, 结果表明围填海导致鱼类的密度和生物量均有所减少, 附近海域的鱼卵和幼鱼密度和生物量也有所下降, 这主要是由于围填海产生的悬浮物导致的(Suo et al, 2015)。
对大型底栖动物长周期的调查和分析, 对于研究环境条件长期变化引起的生物响应具有重要意义(Holland, 1985)。渤海湾围填海工程的影响研究多集中于天津滨海新区沿岸, 对于曹妃甸附近海域的研究极少, 而对曹妃甸海域围填海对大型底栖动物群落的研究则更少。本文利用2013年和2014年的调查数据进行区域和年际比较分析, 明确了曹妃甸围填海工程对大型底栖动物群落的影响范围和程度, 为渤海湾发展规划、环境保护与可持续发展提供基础资料。
1 材料与方法 1.1 站位布设和样品采集分别于2013年9—10月和2014年4—5月在渤海湾布设站位进行调查(图 1), 站位的布设原则主要考虑距离围填海区域的远近及受影响程度的差异, 图中以CFD标记站位为曹妃甸围填海工程区影响较大海域, 其他以BH标记站位为距离围填海区域较远区域, 进行对比。由于受天气海况等影响, 两个航次成功采集的样品有所差异, 2013年成功采集23个站位, 2014年成功采集20个站位。采用0.1m2的采泥器采集底泥, 经0.5mm孔径的网筛分选后用95%的酒精现场固定生物样品, 带回实验室进行种类鉴定、个体计数和称重。样品的处理、保存、称重均按照《海洋调查规范第6部分:海洋生物调查》(GB/T 12763.6-2007) 进行。水深、水温、盐度、溶解氧(DO)和pH等环境因子利用YSI环境监测系统(600QS-M-O, 美国)现场同步测定。营养盐用Niskon采水器采集表层海水, 带回实验室利用营养盐自动流体分析仪(AutoAnalyzer 3, Bran Luebbe, Germany)测定, 测定方法按照《海洋监测规范第4部分:海水分析》(GB 17378.4-2007) 进行。
 |
| 图 1 2013年和2014年渤海湾采样站位 Fig. 1 Sampling stations of macrobenthos off Caofeidian in Bohai Bay in 2013 and 2014 注: ●: 2013年和2014年相同调查站位; △:仅2013年调查站位, +:仅2014年调查站位 |
采用优势度指数计算优势种组成, 多样性指数计算物种多样性特征。计算公式分别如下:
优势度指数(陈亚瞿等, 1995):
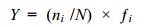 (1)
(1)Shannon-Wiener多样性指数(Shannon et al, 1949):
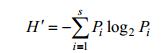 (2)
(2)Margalef物种丰富度指数(Margalef, 1968):
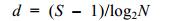 (3)
(3)Pielou物种均匀度指数(Pielou, 1975):
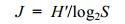 (4)
(4)式中, N为所有站位采集的大型底栖动物总个体数; ni为第i种底栖动物的总个体数; fi为物种i在各站位出现的频率; S为采泥样品中的种类总数; Pi为第i种的个体数与样品中总个体数的比值(ni/N)。当物种优势度Y > 0.02时, 该种即为优势种(陈亚瞿等, 1995)。
1.3 群落结构分析采用PRIMER 6.0进行多元统计分析:首先对物种的丰度数据进行平方根转换以平衡优势种和稀有种在群落中的影响, 并计算站位间的Bray-Curtis相似性系数, 构建相似性矩阵, 然后采用CLUSTER等级聚类方法和非度量多维标度(non-metric multi-dimensional scaling, MDS)排序方法分析大型底栖动物的群落分布格局。通过ANOSIM分析不同聚类组间的差异性, 并通过SIMPER分析(similarity percentage program)不同物种对各群落组内相似性和组间差异性的平均贡献率(Clarke et al, 1994; 周红等, 2003)。
1.4 群落与环境因子相关性分析为减少群落中的机会种对分析结果的干扰, 对两次调查站位的环境因子与大型底栖动物群落的物种数、总丰度、总生物量和群落多样性指数进行对数(ln(x+1))处理后, 利用分析统计软件SPSS进行环境因子与群落生物指数的相关性分析。并利用生物统计软件PRIMER对所有站位的丰度和生物量指数与环境因子进行BVSTEP分析, 获取解释群落结构的最佳环境因子组合(Clarke et al, 1994)。
1.5 群落健康状况评价采用丰度/生物量比较曲线(abundance and biomass curves)分析大型底栖动物群落受扰动状况, 若生物量曲线始终位于丰度曲线之上, 则群落未受干扰; 若生物量和丰度曲线接近重合, 或者出现部分交叉, 则群落受到中等程度的扰动; 若生物量曲线始终位于丰度曲线之下, 则群落受到严重干扰(田胜艳等, 2006)。
采用AMBI 5.0(http://www.azti.es)计算AMBI和M-AMBI指数, 采用2014年物种列表, 根据各种底栖动物对环境的敏感度不同, 将其分为不同的生态组。采用结果中的AMBI最小值多样性指数(H’)的最高值和丰富度指数S的最高值, 并对上述H’值和S值各乘以115%, 计算M-AMBI指数, 设定M-AMBI的阈值如下: “优” > 0.77; “良好”=0.53-0.77; “中等”=0.39-0.53; “较差”=0.20-0.39; “差” < 0.20, 非底栖无脊椎动物(鱼)除外。
2 结果 2.1 渤海湾曹妃甸与附近海域大型底栖动物群落现状 2.1.1 渤海湾曹妃甸与附近海域物种组成2013年渤海湾调查海域共发现大型底栖动物128种, 其中多毛类64种为主要优势类群, 占总物种数的50%, 其他依次为软体动物28种(21.88%), 甲壳动物21种(16.41%), 棘皮动物8种(6.25%), 其他动物7种(5.47%)。
2014年共发现大型底栖动物143种, 其中多毛类59种, 占总物种数的41.26%, 其次为甲壳动物43种, 占30.07%, 软体动物27种(18.88%), 棘皮动物8种(5.59%), 其他动物6种(4.20%)
曹妃甸海域(CFD站位)和附近海域(BH站位)各站位的物种数在不同区域(F=0.013, P=0.909 > 0.05, 单因素方差分析)和不同年间(F=1.508, P=0.226 > 0.05, 单因素方差分析)差异均不显著。曹妃甸海域(CFD站位)两个航次的总物种数均高于附近海域(BH站位), 具体表现为, 2013年调查航次曹妃甸海域的多毛类物种数明显高于附近海域, 而2014年调查航次多毛类和甲壳类物种数均明显高于附近海域, 其他主要类群在区域间和航次间均无明显差异(图 2)。
 |
| 图 2 2013年和2014年曹妃甸和附近海域大型底栖动物物种数和物种组成的比较 Fig. 2 Comparison in species number and composition of macrobenthos between Caofeidian and its adjacent areas in 2013 and 2014 注: CFD为曹妃甸海域, BH为曹妃甸附近海域, 括号中数字为调查年份 |
2013年和2014年曹妃甸和附近海域大型底栖动物群落优势种均为凸壳肌蛤(Musculus senhousia)和日本倍棘蛇尾(Amphioplus japonicus)。曹妃甸区域两个调查航次中, 凸壳肌蛤的优势度2014为0.5, 2013年下降为0.264, 优势度均明显; 日本倍棘蛇尾2013年和2014年的优势度分别为0.036和0.038, 变化较小。附近海域优势度组成两个航次期间则发生变化, 2013年凸壳肌蛤(0.36) 优势度高于日本倍棘蛇尾(0.109); 2014年凸壳肌蛤(0.0260) 优势度明显下降低于日本倍棘蛇尾(0.252)。
2.1.3 曹妃甸以及附近海域大型底栖动物的丰度和生物量2013年总平均丰度为1130.44ind/m2, 贡献率最高的为软体动物699.13ind/m2, 占比61.85%, 其次为多毛类181.74ind/m2(16.08%), 棘皮动物165.22ind/m2(14.62%), 甲壳动物75.65ind/m2(6.69%), 其他动物8.70ind/m2(0.77%)。
2013年总平均生物量为50.44g/m2, 其中棘皮动物生物量最高为23.99g/m2, 占总平均生物量的47.56%, 其次为软体动物18.67g/m2(37.02%), 多毛类5.22g/m2(10.34%), 其他动物1.86g/m2(3.69%), 甲壳动物0.70g/m2(1.39%)。
2014年总平均丰度为698.25ind/m2, 软体动物丰度最高为38.08ind/m2, 占总平均丰度的54.43%, 其次为多毛类123.17ind/m2(17.64%), 棘皮动物119.92ind/m2(17.17%), 甲壳动物68.58ind/m2(9.82%), 其他动物6.5ind/m2(0.93%)。
2014年总平均生物量为38.62g/m2, 其中棘皮动物最多为16.35g/m2, 占总平均生物量的42.33%, 其次为软体动物8.24g/m2(21.35%), 其他动物7.25g/m2(18.77%), 多毛类4.60g/m2(11.91%), 甲壳动物2.18g/m2(5.64%)。
大型底栖动物丰度和生物量不同区域间(F=0.045, P=0.833 > 0.05; F=0, P=0.983 > 0.05, 单因素方差分析)及不同年份间(F=1.521, P=0.224 > 0.05; F=0.480, P=0.492 > 0.05, 单因素方差分析)差异均不显著。大型底栖动物丰度和生物量的空间分布不均匀, 站位间有较大差距(图 3), 主要与个体较小但数量较多的凸壳肌蛤和个体较大但数量较少的哈氏刻勒海胆(Temnopleurus hardwickii)、棘刺锚参(Protankyra bidentata)的不均匀分布有关。
 |
| 图 3 2013年和2014年调查区域大型底栖动物丰度和生物量在各站位的分布 Fig. 3 Spatial distributions of microbenthic abundance and biomass in sampling stations in 2013 and 2014 |
2013年曹妃甸海域的物种丰富度指数d、物种均匀度指数J’和香农威纳多样性指数H’平均值分别为2.22±1.10, 0.60±0.29和2.43±1.35。3个多样性指数最高值均出现在CFD29站位, 最低值均出现在CFD42站位。附近海域的物种丰富度指数d、物种均匀度指数J’和香农威纳多样性指数H’平均值分别为2.16±0.78, 0.61±0.27, 2.32±0.95。丰富度指数的最高值位于BH08站位, 均匀度指数的最高值位于BH11站, 香农威纳多样性指数最高值位于BH23站位, 3个指数的最低值均位于BH15站位。
2014年曹妃甸海域物种丰富度指数d、物种均匀度指数J’和香农威纳多样性指数H’平均值分别为3.08±1.48, 0.79±0.23和3.12±1.12。丰富度指数d指数、物种多样性指数H’最高值均位于CFD33站位, 物种均匀度指数J’最高值均位于CFD31;丰富度指数d指数最低值位于CFD39, 物种均匀度指数J’和香农维纳多样性指数H’最低值均位于CFD38。附近海域的丰富度指数d平均值为2.83±0.96, 最高值位于BH10站位, 最低值位于BH18;物种均匀度指数J’和香农威纳多样性指数H’平均值分别为0.74±0.21和2.84±0.75, 最高值位于BH25, 最低值位于BH24。
航次间比较, 曹妃甸海域生物多样性指数均高于附近海域; 区域间比较, 2014年均高于2013年(图 4)。物种丰富度指数和均匀度指数在不同年间有显著差异(F=5.350, P=0.026 < 0.05; F=4.651, P=0.037 < 0.05, 单因素方差分析), 区域间无显著差异(F=0.331, P=0.568 > 0.05; F=0.102, P=0.751 > 0.05, 单因素方差分析)香农威纳多样性指数区域和年份间均无差异(F=0.454, P=0.504 > 0.05; F=3.693, P=0.062 > 0.05, 单因素方差分析)。
 |
| 图 4 2013年和2014年大型底栖动物生物多样性指数 Fig. 4 Species biodiversity indices of microbenthic communities in 2013 and 2014 注: CFD为曹妃甸海域, BH为曹妃甸附近海域, 数字为调查年份 |
大型底栖动物群落结构聚类分析结果见图 5, 且MDS分析与CLUSTER聚类分析结果一致。ANOSIM分析表明, 两次调查的不同聚类组间均存在显著差异(2013年R=0.858, P < 0.01; 2014年R=0.921, P < 0.01, Global Test)。
 |
| 图 5 2013年和2014年大型底栖动物群落CLUSTER和MDS分析 Fig. 5 CLUSTER and MDS analyses on macrobenthos in 2013 and 2014 |
以20%的相似性, 2013年调查区域的大型底栖动物群落可分为8组, 其中CFD37、BH12、CFD38、BH23单独成组, 其他3组分别为:
组Ⅰ:由13个站位组成, 组内平均相似性为35.61%, 主要贡献物种为凸壳肌蛤和日本倍棘蛇尾, 贡献率分别为60.72%和10.64%。
组Ⅱ:由BH21、BH22、CFD29组成, 组内平均相似性为32.87%, 主要贡献物种为日本倍棘蛇尾(32.84%)、深沟毛虫(Sigambra bassi)(13.89%)和双栉虫(Ampharete acutifrons)(13.89%)。
组Ⅲ:包括CFD31和CFD32两个站位, 组内平均相似性为35.67%, 主要贡献物种为侧带裁判螺(Inquisitor latifasciata)(39.97%)、凸壳肌蛤(15.68%)、豆形胡桃蛤(Nucula faba)(11.09%)、丝异蚓虫(Heteromastus filiformis)(11.09%)、长须沙蚕(Nereis longior)(11.09%)、笔帽虫(Pectinaridae)(11.09%)。
以20%的相似性, 2014年调查区域的大型底栖动物群落可分为7组, 其中CFD37、CFD42单独聚为一组, 其他5组分别为:
组Ⅰ:由9个站位组成, 组内平均相似性为26.56%, 主要贡献物种为日本倍棘蛇尾和拟特须虫(Paralacydonia paradoxa), 贡献率分别为34.47%和13.40%。
组Ⅱ:由BH12、BH25、CFD43组成, 组内平均
相似性为27.10%, 主要贡献物种为豆形胡桃蛤(20.54%)、纽虫(Nemertinea)(20.54%)和凹鳍孔鰕虎鱼(Ctenotrypauchen chinensis)(20.54%)。
组Ⅲ:由BH18和CFD39组成, 组内平均相似性为36.02%, 主要贡献物种为塞切尔尼钩虾(Eriopisella sechellensis)(50%)和日本倍棘蛇尾(50%)。
组Ⅳ:由BH24和CFD38组成, 组内平均相似性为29.97%, 主要贡献物种为凸壳肌蛤。
组Ⅴ:由CFD34、CFD35、CFD36三个站位组成, 组内平均相似性为24.45%, 主要贡献物种为多鳃齿吻沙蚕(Nephtys polybranchia) (18.92%)、独指虫(Aricidea fragilis) (17.83%)、寡节甘吻沙蚕(Glycinde gurjanovae) (13.38%)、塞切尔尼钩虾(10.02%)。
2.1.6 大型底栖动物群落与环境因子的关系2013年生物群落指数与环境因子相关性(Pearson Correlation)分析表明, 物种数与总碳、总有机碳和总氮含量、硝酸盐、总溶解态磷呈显著负相关(R= –0.557, P=0.006 < 0.01; R= –0.582, P=0.004 < 0.01; R= –0.721, P=0 < 0.01; R= –0.457, P=0.028 < 0.05; R= –0.444, P=0.034 < 0.05);总丰度与铵盐呈极显著负相关(R= –0.570, P=0.004 < 0.01);物种丰富度指数与总碳、总有机碳和总氮均呈极显著负相关(R= –0.567, P=0.005 < 0.01; R= –0.600, P=0.002 < 0.01; R= –0.706, P < 0.01);均匀度指数与铵盐呈显著正相关(R=0.476, P=0.022 < 0.05), 与硅酸盐、总碳、总有机碳、总氮呈显著负相关(R= –0.482, P=0.020 < 0.05; R= –0.429, P= 0.041 < 0.05; R= –0.428, P=0.042 < 0.05; R= –0.509, P= 0.013 < 0.05);香农威纳多样性指数与硅酸盐、总碳、总有机碳和总氮呈显著负相关(R= –0.466, P=0.025 < 0.05; R= –0.507, P=0.014 < 0.05; R= –0.502, P=0.015 < 0.05; R= –0.623, P=0.001 < 0.01)。BVSTEP分析表明, 铵盐是影响该航次大型底栖动物丰度和生物量分布的关键环境因子, 与丰度的Spearman相关系数为0.310, 与生物量的Spearman相关系数为0.240。
2014年生物群落与环境因子相关性(Pearson Correlation)分析表明, 总生物量与盐度、溶解氧呈显著负相关(R= –0.476, P=0.034 < 0.05; R= –0.488, P= 0.029 < 0.05), 与亚硝酸盐呈显著正相关(R=0.477, P= 0.033 < 0.05), 与硝酸盐呈极显著正相关(R=0.568, P= 0.009 < 0.01);均匀度指数与深度呈显著正相关(R= 0.533, P=0.016 < 0.05);香农威纳多样性指数与深度呈极显著正相关(R=0.598, P=0.005 < 0.01), 与温度呈显著负相关(R= –0.518, P=0.019 < 0.05)。BVSTEP分析表明, 温度、溶解氧和硝酸盐是影响该航次大型底栖动物丰度分布的关键环境因子, 与丰度的Spearman相关系数为0.454, 溶解氧、酸碱度和硝酸盐是影响该航次大型底栖动物生物量分布的关键环境因子, 与生物量的Spearman相关系数为0.368。
2.1.7 大型底栖动物群落受扰动状况2013年和2014年的ABC曲线分析表明, 渤海湾大型底栖动物群落均已受到中等程度的干扰, 2014年大型底栖动物群落生物量优势有所上升(图 6), 说明生物群落受扰动状况较2013年有所改善。
 |
| 图 6 2013年和2014年调查区域ABC曲线 Fig. 6 The ABC curves at all stations in study areas in 2013 and 2014 |
2013年和2014年调查站位M-AMBI指数空间分布如图 7, 曹妃甸围填海区域大型底栖动物群落健康状况明显较差, 2014年大型底栖动物群落状况明显优于2013年, 与ABC曲线分析的受扰动结果一致。
 |
| 图 7 2013年(左图)和2014年(右图)渤海湾M-AMBI指数分布 Fig. 7 Distributions of M-AMBI indices in Bohai Bay in 2013 (left) and 2014 (right) 注: “优” > 0.77; “良好”=0.53-0.77; “中等”=0.39-0.53; “较差”=0.20-0.39; “差” < 0.20 |
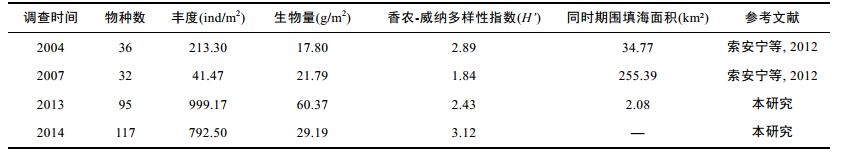
马万栋等(2015)统计了环渤海围填海不同区域不同时期的围填海面积, 其中也涵盖本调查CFD站位的唐山市和秦皇岛市同时期围填海面积(表 1)。2005—2010年曹妃甸海域围填海面积迅速增加, 之后又有所减少(马万栋等, 2015)。索安宁等(2012)对曹妃甸围填海工程环境影响进行了回顾性评价, 2007年大型底栖动物的物种数、丰度和香农威纳多样性指数均明显较低, 这可能与该时期曹妃甸围填海面积迅速增加有关(索安宁等, 2012)。本次调查发现2013—2014年该区域的物种数和生物多样性均有所上升, 这与近年来围填海面积有所下降有关, 生物多样性有所恢复。相关性分析表明围填海面积与大型底栖动物的物种数、丰度、生物量和香农威纳多样性指数均呈负相关关系, 但均未达显著水平(R= –0.644, P=0.555 > 0.05; R= –0.725, P=0.483 > 0.05; R= –0.529, P=0.645 > 0.05; R= –0.841, P=0.364 > 0.05, Pearson相关)。
|
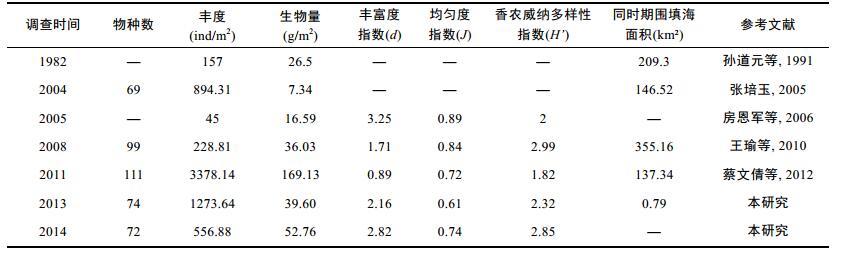
天津沿海区域大型底栖动物群落研究较多, 本文取孙道元等(1991)、张培玉(2005)、房恩军等(2006)、王瑜等(2010)、蔡文倩等(2012)以及本研究2013和2014年的调查数据, 结合马万栋等(2015)同时期围填海面积进行相关性分析(表 2)。围填海面积与大型底栖动物物种数、物种均匀度指数和香农威纳多样性指数成正相关(R=0.467, P=0.533 > 0.05; R=0.702, P=0.298 > 0.05; R=0.647, P=0.353 > 0.05, Pearson相关), 与丰度、生物量、物种丰富度指数呈负相关(R= –0.413, P=0.587 > 0.05; R= –0.101, P=0.899 > 0.05; R= –0.173, P=0.827 > 0.05, Pearson相关), 但均未达显著水平。生物多样性的年际变化显示大型底栖动物多样性在2010年左右有所下降, 近年来有所回升, 与2008年围填海面积急剧增加以及近年来围填海面积有所下降有关。大型底栖动物丰度和生物量受某些机会种如凸壳肌蛤或者重量较大个体的采集影响较大, 2011年大型底栖动物物种数、丰度和生物量明显高于其他年份, 主要因为该年份采集到大量的凸壳肌蛤和长偏顶蛤(蔡文倩等, 2012), 也与2010年围填海面积明显减少有关。
|
曹妃甸2003— 2011年围海造地面积约274.7km, 其中2007—2008年围填海面积增幅最大, 之后几年有所下降(吴越等, 2013)。曹妃甸海域大型底栖动物群落年际比较发现, 2007年大型底栖动物的物种数、丰度和多样性指数有所下降, 由此可以推断, 围填海工程对该区域的大型底栖动物群落造成了一定的影响。
张壮壮等(2015)利用1984、2005和2007—2011年的数据分析, 发现天津沿岸围填海工程引起的岸线长度、占用滩涂面积和近岸浅海面积, 与大型底栖动物总物种数、生物多样性指数呈负相关, 与总生物量、总丰度呈正相关关系。同时改变了群落中不同类群的组成, 即减少甲壳动物和软体动物比例, 增加多毛类种类数量所占比例。本文分析结果发现围填海面积与大型底栖动物丰度和生物量呈负相关关系, 与物种数和香农威纳多样性指数成正相关, 但相关性均未达显著水平, 这可能与我们选取的调查数据季节不同, 且本文的围填海面积为相同时间段的总面积, 与生物数据并非一一对应。张壮壮等(2015)文中也提及研究受调查时间、站点位置和数目、采泥器种类、网筛孔径等影响, 调查统计结果会有所差异。同时, 丰度与生物量受个别机会种的大量采集与重量较大个体的偶然采集影响较大, 无法准确反映大型底栖动物群落扰动状况。
渤海湾2005—2010年围填海面积较大, 大型底栖动物群落波动较大。近年来随着围填海面积的减少, 大型底栖动物群落多样性有所恢复。曹妃甸与附近海域的大型底栖动物群落差异不显著, 与渤海湾曹妃甸围填海工程浩大, 且围填海区域较广, 对渤海湾影响范围较大有关。同时也与渤海湾受人类扰动因素众多, 且围填海工程近年来有所减少, 沿岸群落有所恢复有关。
3.1.2 优势种和优势类群优势种由于其高度的生态适应性, 对群落结构的分析具有重要的作用。蔡文倩等(2012)发现渤海湾生态环境受到严重干扰, 已发生改变, 不再适应对环境敏感的物种生存, 为干扰耐受种提供更多生态, 导致其扩张占据主导地位。自2011年来干扰耐受种凸壳肌蛤一直占据优势地位。本调查发现2013年和2014年凸壳肌蛤优势地位依然显著, 但干扰不敏感种日本倍棘蛇尾的出现, 预示着生态环境的逐渐改善, 逐渐适应其他物种的生存。
2013年和2014年两个航次的大型底栖动物群落优势类群为多毛类, 曹妃甸2014年甲壳动物明显增多, 所占比例仅次于多毛类, 但是两个区域两次调查中多毛类和甲壳动物的生物量均较低, 高丰度和低生物量可以看出调查区域物种小型化依然明显。
3.1.3 群落健康状况2013年和2014年大型底栖动物群落已受到中等程度的干扰, 曹妃甸围填海工程区大型底栖动物群落健康状况明显较差。Lu等(2002)发现Sungei Punggol河靠近围填海区的大型底栖动物群落物种数和丰度明显下降。Li等(2010)发现天津港工业区围填海工程导致大型动物多样性下降甚至灭绝。索安宁等(2012)对曹妃甸围填海工程的回顾性评价中提出, 围填海对底栖动物的影响最大。Suo等(2015)研究发现曹妃甸围填海工程对鱼类群落造成明显的损害, 鱼类丰度、生物量明显下降。由此可以看出, 围填海对附近海域的生物群落造成了明显的扰动, 影响不容忽视。2014年的大型底栖动物群落状况优于2013年, 这与2014年为春季调查航次, 处于大型底栖动物的繁殖季节, 而2013年为秋季调查航次有关。
3.2 围填海对大型底栖动物影响的原因围填海工程占用近岸浅滩, 滩涂湿地为鱼虾类的重要产卵场和索饵场, 浅滩填埋造陆后, 生物全部死亡, 造成生物群落不可逆的损害。围填海工程导致近岸潮流水动力减弱, 淤泥质粉土沉降堆积, 底质类型改变, 水深逐渐变浅, 改变了大型底栖动物生境状况。
底质类型和水深是影响大型底栖动物分布的重要环境因子(李宝泉等, 2005; 李新正等, 2006), 渤海湾优势种主要为凸壳肌蛤, 与该区域的粘土质粉砂为主有关。对2013年和2014年的生物群落进行聚类分组发现, 群落间相似性较低, 这由于渤海湾沿岸受人为干扰严重和环境复杂, 形成了不同的小生境, 导致各个站位的大型底栖动物群落相似性不高(蔡文倩等, 2013)。
大规模的围填海工程改变了原来浅海区的海洋动力边界条件, 势必引起周围海域的潮流、冲淤过程和泥沙迁移的改变, 使近岸海水与外海交换能力下降, 进而导致海水富营养化。
渤海1960—1996年的环境变化和生态系统反应研究发现, 渤海的盐度、温度、溶解无机氮、氮磷比呈上升趋势, 溶解氧、磷、硅和硅氮比呈下降趋势。渤海由于人类的干扰和气候的变化, 造成淡水补给的减少, 进而导致磷、硅等营养盐的限制, 造成渤海生物系统衰退和生物多样性减少(Ning et al, 2010)。而本次调查发现, 大型底栖动物物种数、丰富度指数和、均匀度指数和物种多样性指数与总碳、总有机碳和总氮呈显著或者极显著负相关, 物种数与总溶解态磷呈显著负相关, 物种均匀度指数、多样性指数与硅酸盐呈显著负相关。由此可以看出渤海的富营养化的状态和趋势对大型底栖动物群落已产生不同程度的影响。
围填海疏浚和吹填过程中, 产生高浓度的悬浮物, 造成水体的透明度下降, 影响浮游植物的光合作用, 进而影响以浮游生物为食的大型底栖动物的正常生长(Newcombe et al, 1991)。另外, 许多大型底栖动物幼体营浮游生活, 幼体对悬浮物的抵抗力较差, 悬浮物堵塞幼体的鳃部致其死亡, 直接影响对成体的补充。实验发现荔枝螺幼体对悬浮物的耐受性很差, 悬浮物浓度达到2mg/L时, 将对荔枝螺造成急性毒性效应, 且随着悬浮物浓度的上升, 致死率明显上升(本研究), 杜东等(2012)对曹妃甸近岸海域2009年悬浮泥沙含量分布发现近岸5m以浅地区的悬沙浓度多大于10mg/L, 势必对大型底栖动物幼体造成一定的影响。
4 结论(1) 渤海湾大型底栖动物群落的物种组成仍以多毛类占据主要优势, 优势种主要以干扰耐受种凸壳肌蛤为主, 调查区域物种小型化特征明显。曹妃甸围填海区域与附近海域的物种组成、丰度和生物量以及多样性指数差异并不显著, 与渤海湾沿岸围填海分布较广, 影响较大, 以及近年来生物群落有所恢复有关。
(2) 渤海湾大型底栖动物群落的物种数、丰度和生物量以及多样性指数与围填海面积相关性不显著, 但围填海面积较大的时期, 大型底栖动物群落明显波动较大, 曹妃甸2005—2010年围填海面积明显增多, 而2007年大型底栖动物物种数、丰度和生物多样性指数明显较低, 说明围填海对大型底栖动物群落具有强烈的扰动影响。
(3) 2013年和2014年的渤海湾大型底栖动物群落已受到中等程度的扰动, 水体呈富营养化的状态和趋势, 并对大型底栖动物群落产生了不同程度的影响。
(4) 大型底栖动物丰度、生物量受机会种和个体较大个体的偶然采集影响较大, 对群落受扰动状况反映能力较差, M-AMBI指数对渤海大型底栖动物群落健康状况反映情况较佳, 曹妃甸围填海区大型底栖动物群落生态健康状况明显较差。
| 马万栋, 吴传庆, 殷守敬, 等, 2015. 环渤海围填海遥感监测及对策建议. 环境与可持续发展,(3): 63–65 |
| 王瑜, 刘录三, 刘存歧, 等, 2010. 渤海湾近岸海域春季大型底栖动物群落特征. 环境科学研究, 23(4): 430–436 |
| 尹延鸿, 2007. 对河北唐山曹妃甸浅滩大面积填海的思考. 海洋地质动态, 23(3): 1–10 |
| 田胜艳, 于子山, 刘晓收, 等, 2006. 丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究. 海洋通报, 25(1): 92–96 |
| 孙道元, 刘银城, 1991. 渤海底栖动物种类组成和数量分布. 黄渤海海洋, 9(1): 42–50 |
| 杜东, 刘宏伟, 秦雅飞, 等, 2012. 河北省曹妃甸近岸海域悬浮泥沙含量分布特征研究. 地质调查与研究, 35(2): 189–194 |
| 李宝泉, 李晓静, 周政权, 等, 2016. 围填海及其对底栖生物群落的生态效应. 广西科学, 23(4): 293–298 |
| 李宝泉, 李新正, 于海燕, 等, 2005. 胶州湾底栖软体动物与环境因子的关系. 海洋与湖沼, 36(3): 193–198 |
| 李新正, 2011. 我国海洋大型底栖生物多样性研究及展望:以黄海为例. 生物多样性, 19(6): 676–684 |
| 李新正, 李宝泉, 王洪法, 等, 2006. 胶州湾潮间带大型底栖动物的群落生态. 动物学报, 52(3): 612–618 |
| 肖庆聪, 魏源送, 王亚炜, 等, 2012. 天津滨海新区湿地退化驱动因素分析. 环境科学学报, 32(2): 480–488 |
| 吴越, 杨文波, 王琳, 等, 2013. 曹妃甸填海造地时空分布遥感监测及其影响初步研究. 海洋湖沼通报,(1): 153–158 |
| 张壮壮, 王琳, 杨文波, 等, 2015. 渤海湾近海栖息地变化对大型底栖动物群落结构的影响. 中国水产科学, 22(1): 106–112 |
| 张培玉, 2005. 渤海湾近岸海域底栖动物生态学与环境质量评价研究. 青岛: 中国海洋大学博士学位论文, 86-92 |
| 陈亚瞿, 徐兆礼, 王云龙, 等, 1995. 长江口河口锋区浮游动物生态研究Ⅰ:生物量及优势种的平面分布. 中国水产科学, 2(1): 49–58 |
| 周红, 张志南, 2003. 大型多元统计软件primer的方法原理及其在底栖群落生态学中的应用. 青岛海洋大学学报, 33(1): 58–64 |
| 房恩军, 李军, 马维林, 等, 2006. 渤海湾近岸海域大型底栖动物(Macrofauna)初步研究. 现代渔业信息, 21(10): 11–15 DOI:10.3969/j.issn.1004-8340.2006.10.004 |
| 索安宁, 张明慧, 于永海, 等, 2012. 曹妃甸围填海工程的环境影响回顾性评价. 中国环境监测, 28(2): 105–111 |
| 蔡文倩, 孟伟, 刘录三, 等, 2012. 渤海湾大型底栖动物群落优势种长期变化研究. 环境科学学报, 33(8): 2332–2340 |
| 蔡文倩, 孟伟, 刘录三, 等, 2013. 春季渤海湾大型底栖动物群落结构特征研究. 环境科学学报, 33(5): 1458–1466 |
| Clarke K R, Warwick R M, 1994. Change in Marine Communities:An Approach to Statistical Analysis and Interpretation. Plymouth:Plymouth Marine Laboratory |
| Holland A F, 1985. Long-term variation of macrobenthos in a mesohaline region of Chesapeake bay. Estuaries, 8(2): 93–113 DOI:10.2307/1351860 |
| Li K Y, Liu X B, Zhao X G, et al, 2010. Effects of reclamation projects on marine ecological environment in Tianjin Harbor Industrial Zone. Procedia Environmental Sciences, 2: 792–799 DOI:10.1016/j.proenv.2010.10.090 |
| Lu L, Goh B P L, Chou L M, 2002. Effects of coastal reclamation on riverine macrobenthic infauna (Sungei Punggol) in Singapore. Journal of Aquatic Ecosystem Stress and Recovery, 9(2): 127–135 DOI:10.1023/A:1014483804331 |
| Margalef R, 1968. Perspectives in Ecological Theory. New York, USA: University of Chicago Press, |
| Newcombe C P, Macdonald D D, 1991. Effects of suspended sediments on aquatic ecosystems. North American Journal of Fisheries Management, 11(1): 72–82 DOI:10.1577/1548-8675(1991)011<0072:EOSSOA>2.3.CO;2 |
| Ning X R, Lin C L, Su J L, et al, 2010. Long-term environmental changes and the responses of the ecosystems in the Bohai Sea during 1960-1996. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 57(11-12): 1079–1091 DOI:10.1016/j.dsr2.2010.02.010 |
| Pielou E C, 1975. Ecological Diversity. New York, USA: Wiley-Interscience, |
| Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana, USA: University of Illinois Press, |
| Suo A N, Cao K, Zhao J H, et al, 2015. Study on impacts of sea reclamation on fish community in adjacent waters:a case in Caofeidian, North China. Journal of Coastal Research, 73: 183–187 DOI:10.2112/SI73-032.1 |
| Wang W, Liu H, Li Y Q, et al, 2014. Development and management of land reclamation in China. Ocean & Coastal Management, 102: 415–425 |