 2017, Vol. 48
2017, Vol. 48

胶州湾表层沉积物中甲藻孢囊的分布

http://dx.doi.org/10.11693/hyhz20161200283
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 李影, 汤亚楠, 沈萍萍, 顾海峰, 谭烨辉. 2017.
- LI Ying, TANG Ya-Nan, SHEN Ping-Ping, GU Hai-Feng, TAN Ye-Hui. 2017.
- 胶州湾表层沉积物中甲藻孢囊的分布
- DISTRIBUTION OF DINOFLAGELLATE RESTING CYSTS IN SURFACE SEDIMENT OF JIAOZHOU BAY, CHINA
- 海洋与湖沼, 48(4): 760-766
- Oceanologia et Limnologia Sinica, 48(4): 760-766.
- http://dx.doi.org/10.11693/hyhz20161200283
-
文章历史
- 收稿日期:2016-12-20
- 收修改稿日期:2017-03-03
引用本文
李影, 汤亚楠, 沈萍萍, 顾海峰, 谭烨辉. 2017. 胶州湾表层沉积物中甲藻孢囊的分布[J]. 海洋与湖沼, 48(4): 760-766
LI Ying, TANG Ya-Nan, SHEN Ping-Ping, GU Hai-Feng, TAN Ye-Hui. 2017. DISTRIBUTION OF DINOFLAGELLATE RESTING CYSTS IN SURFACE SEDIMENT OF JIAOZHOU BAY, CHINA[J]. Oceanologia et Limnologia Sinica, 48(4): 760-766.
胶州湾表层沉积物中甲藻孢囊的分布
1.
中国科学院南海海洋研究所 热带海洋生物资源与生态重点实验室 广州 510301;
2. 中国科学院大学 北京 100049;
3. 国家海洋局第三海洋研究所 厦门 361005
2. 中国科学院大学 北京 100049;
3. 国家海洋局第三海洋研究所 厦门 361005
摘要:为了解胶州湾表层沉积物中甲藻孢囊的分布特征,于2015年7月采集胶州湾12个站位的表层沉积物进行甲藻孢囊分析,共鉴定甲藻孢囊37种(分属5个类群)和2种未定种,包括自养型20种和异养型17种,优势种为膝沟藻。其Shannon-Wiener多样性指数[H'(log2)]介于2.84-3.91,平均值为3.42;均匀度指数介于0.75-0.98,平均值为0.88,总体值较高。胶州湾海域各站位的甲藻孢囊丰度介于96-969cyst/g DW(干重),平均丰度为401cyst/g DW,呈现出临近湾口处丰度较高,而湾内较低的空间分布差异特征。尤其值得注意的是,有两种产虾夷扇贝毒素(Yessotoxin,YTX)甲藻[具刺膝沟藻(Gonyaulax spinifera)和多边舌甲藻(Lingulodinium polyedrum)]的孢囊和3种产麻痹性贝类毒素(PSP)甲藻[塔玛/链状亚历山大藻(Alexandrium catenella/tamarense)、微小/相似亚历山大藻(A.minutum/affine)和链状裸甲藻(Gymnodinium catenatum)]的孢囊在胶州湾海域大多数站位均有分布,尽管其数量不高,但其广泛的分布范围仍需引起我们的重视。
关键词:胶州湾 甲藻孢囊 生物多样性 分布
DISTRIBUTION OF DINOFLAGELLATE RESTING CYSTS IN SURFACE SEDIMENT OF JIAOZHOU BAY, CHINA
1.
Key Laboratory of Tropical Marine Bio-resources and Ecology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China;
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Third Institute of Oceanography, State Oceanic Administration, Xiamen 361005, China
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Third Institute of Oceanography, State Oceanic Administration, Xiamen 361005, China
Abstract:
Distribution of dinoflagellate resting cysts in surface sediment of Jiaozhou Bay was investigated in July, 2015. In total, about 37 species representing 5 groups and 2 uncertain taxa were identified from 12 sediment samples, including 20 autotrophic and 17 heterotrophic species. The dominant taxa were Gonyaulax spp. The cyst species diversity (Shannon-Wiener index) ranged from 2.84 to 3.91 at each station, in mean of 3.42 across the bay. In addition, Pielou's evenness index ranged from 0.75 to 0.98, and the average was 0.88, indicating a relatively stable phytoplankton community. However, cyst abundance showed a relative spatial variation from 96 to 969cyst/g DW, in average of 401cyst/g DW across the whole study area. It was generally high around the mouth of the bay and low in inner bay. Besides the dominant species Gonyaulax sp., the resting cysts of several toxic groups were also recognized, including G. spinifera and Lingulodinium polyedrum, Alexandrium catenella/tamarense, A. minutum/affine and Gymnodinium catenatum. Although in relative low abundance, these toxic species could be observed at most stations, indicating a wide distribution in Jiaozhou Bay.
Key words:
Jiaozhou Bay
dinoflagellate cyst
diversity
distribution
甲藻是海洋浮游生物的第二大类群, 其种类和数量仅次于硅藻, 在海洋生态系统中占有非常重要的地位。甲藻孢囊是甲藻度过不良环境的一种休眠方式, 研究表明至今大约200多种甲藻能够形成孢囊(王朝晖, 2007a; Matsuoka et al, 2013)。一般的孢囊是指经有性生殖所形成的休眠孢囊, 对孢囊的研究多围绕此类, 但最新研究结果表明, 在某些甲藻中也存在无性生殖所产生的休眠孢囊(王朝晖, 2007b)。休眠孢囊沉降在海底沉积物中以抵抗不良的外部环境, 一旦环境条件适宜则会再度萌发成营养细胞进入上覆水体, 然后以无性繁殖方式进行增殖, 成为引发赤潮的种源(肖咏之等, 2001; Fertouna-Bellakhal et al, 2015)。由于沉积物中甲藻孢囊是某一时期海域所形成的孢囊集群, 而且大多数甲藻孢囊具有孢粉素的细胞壁, 可长期保存在沉积物中, 因此, 沉积物甲藻孢囊能在一定程度上反映出水体中浮游甲藻的历史信息, 成为追溯各海域浮游植物历史记录和水体环境变化过程有效的信号载体(孙爱梅等, 2006)。孢囊的形成对于物种生存、繁殖、分布与传播等具有重要的生物学意义, 同时也是赤潮发生的“种源”和赤潮消亡的重要原因, 孢囊研究因此成为海洋生态学、海洋地质学和环境学的国际研究热点(Matsuoka et al, 2013; Fertouna-Bellakhal et al, 2015)。目前很多学者对我国沿海重要海湾的沉积物甲藻孢囊进行了较为广泛的调查研究, 取得了一系列的研究成果(肖咏之等, 2001; Wang et al, 2004;黄海燕等, 2009;顾海峰等, 2011;石雅君等, 2011;王艳等, 2012)。但迄今为止, 有关胶州湾海域沉积物甲藻孢囊的研究鲜有报道, 因此, 本文对胶州湾海域表层沉积物中甲藻孢囊的种类及数量分布进行了初步探索, 以期为补充此海域甲藻孢囊的分布状况与多样性认识, 亦为甲藻有毒有害藻华的防御、监测提供一些基础数据与参考资料。
1 材料与方法 1.1 采样区及采样方法胶州湾位于山东半岛南部, 黄海东部, 为半封闭型海湾, 海湾面积约397km2, 其中约64%的海域水深不超过5m, 湾内与外海水体交换情况良好, 半交换周期为5天(杨世民等, 2014)。其东部为港口区, 西部与西北部主要为海水养殖区, 外与黄海海水交换, 湾底部有大沽河、海泊河、李村河、娄山河等河流流入(卢敏等, 2001)。近几十年来, 随着青岛工农业的迅速发展, 大量的营养物质排放入海, 导致水中有机质增多, 赤潮现象频繁发生。1990年在胶州湾的中部海域发生由红色中缢虫大量增殖引起的赤潮, 1997年7月胶州湾东部海域发生由短角弯角藻(Eucampia zoodiacus)过度增殖引起的赤潮, 1998年发生中肋骨条藻(Skeletonema costatum)赤潮, 1999年6—8月, 分别发生中肋骨条藻、短角弯角藻、高贵齿状藻(盒型藻)(Odontella regis)赤潮, 给当地的海洋渔业养殖环境造成了极大的影响(卢敏等, 2001;韩笑天等, 2004)。
2015年7月11—12日, 在胶州湾设12个站位(S3—S14)(图 1), 使用抓斗式采泥器采集表层沉积物。样品采集后置于冰盒中带回实验室, 并于4℃冰箱中保存。
 |
| 图 1 胶州湾表层沉积物采样站位图 Fig. 1 Sediment sampling stations in the Jiaozhou Bay |
取适量沉积物样品分成两份, 其中一份(约10g)放在60℃烘箱中烘干至恒重, 用来测定样品含水率。另一份(约5g)进行网筛过滤, 即超声波震荡之后, 让其通过125μm和15μm的不锈钢网筛, 收集两个网筛之间的部分并转移至表面皿中, 旋转表面皿使液体涡旋(王朝晖等, 2003), 用吸管吸取上层浮液至15mL离心管中, 自然沉降12h, 弃上清液, 用过滤海水定容至10mL。
进行甲藻孢囊分析鉴定时, 吸取1mL已经处理好的样品至1mL浮游生物计数框(Sedgewick-Rafter 1mm2)中, 然后将其置于徕卡显微镜(DM2000)200倍显微镜下对孢囊依次进行种类的鉴定、计数和拍照。在鉴定过程中, 遇到无法鉴定的甲藻孢囊种类时, 以其所在的属来表示。孢囊密度以每克干重沉积物中的孢囊数表示(cyst/g DW)(王朝晖, 2007b)。
1.3 统计分析方法物种多样性指数采用香农-威纳指数(H′, Shannon-Wiener index)(Shannon et al, 1949), 其计算公式为
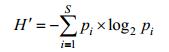
物种均匀度指数(J)(Pielou, 1969), 其计算公式:
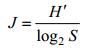
优势种类的优势度指数(Y > 0.02) 判断, 计算公式为:

式中, S为样品中总种类数, pi为第i种的个体数与样品中的总个数(N)的比值, ni为第i种的总个体数, N为所有物种的总个体数, fi为第i种在各站位出现的频率。
2 结果与讨论 2.1 甲藻孢囊的种类组成胶州湾12个表层沉积物中总共鉴定到37种甲藻孢囊和2种未定种(表 1)。其中37种甲藻孢囊分属5个类群:膝沟藻类(Gonyaulacoid group, 16种), Tuberculodinioid类(Tuberculodinioid group, 1种), 钙质类(calciodinellid group, 1种), 裸甲藻类(Gymnodinioid group, 3种)和原多甲藻类(Protoperidinioid group, 16种)。
表 1 胶州湾表层沉积物中甲藻孢囊分布情况 Tab. 1 Distribution of dinoflagellate cysts in the sediment of Jiaozhou Bay
| 孢囊种类 | 站位 | ||||||||||||||
| 中文名 | 藻类学学名 | 孢粉学名称 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| 自养型 | |||||||||||||||
| 膝沟藻类 | |||||||||||||||
| 塔马亚历山大藻/链状亚历山大藻 | Alexandrium catenella / tamarense | + | + | + | + | ||||||||||
| 微小亚历山大藻/相似亚历山大藻 | Alexandrium minutum /affine | + | |||||||||||||
| 具指膝沟藻 | Gonyaulax digitale | Spiniferites bentori | + | + | + | + | + | + | + | + | + | ||||
| 长膝沟藻 | Gonyaulax elongata | + | + | + | + | + | + | + | + | + | + | + | |||
| 斯氏膝沟藻 | Gonyaulax scrippsae | Spiniferites bulloideus | + | + | + | + | + | + | + | + | |||||
| 膜状膝沟藻 | Gonyaulax membranacea | Spiniferites membranaceus | + | + | |||||||||||
| 具刺膝沟藻 | Gonyaulax spinifera | Spiniferites ramosus | + | + | |||||||||||
| 具刺膝沟藻 | Gonyaulax spinifera | Spiniferites hyperacanthus | + | + | + | + | + | + | + | + | + | + | + | ||
| 膝沟藻 | Gonyaulax sp. | Spiniferites sp. | + | + | + | + | + | + | + | + | |||||
| 具刺膝沟藻 | Gonyaulax spinifera | Spiniferites mirabilis | + | + | + | + | + | + | + | + | + | ||||
| 膝沟藻 | Gonyaulax sp. | Spiniferites bentori var. truncate | + | + | + | ||||||||||
| 多边舌甲藻 | Lingulodinium polyedra | Lingulodinium machaerophorum | + | + | + | + | + | + | + | + | + | + | + | ||
| 网状原角管藻 | Protoceratium reticulatum | Operculodinium centrocarpum | + | + | + | + | + | + | + | + | + | + | |||
| 膝沟藻 | Gonyaulax sp. | Spiniferites delicatus | + | + | + | ||||||||||
| 膝沟藻 | Gonyaulax sp. | Spiniferites sp. cf. delicatus | + | ||||||||||||
| 膝沟藻属 | Gonyaulax sp. | Spiniferites sp. | + | + | + | + | + | + | + | + | + | + | |||
| Tuberculodinioid类 | |||||||||||||||
| 窄形扁甲藻 | Pyrophacus steinii | Tuberculodinium vancampoae | + | + | + | + | + | ||||||||
| 钙质类 | |||||||||||||||
| 锥状斯氏藻 | Scrippsiella trochoidea | + | + | + | + | + | |||||||||
| 裸甲藻类 | |||||||||||||||
| 哈曼褐多沟藻 | Pheopolykrikos hartmannii | + | |||||||||||||
| 链状裸甲藻 | Gymnodinium catenatum | + | + | + | |||||||||||
| 异养型 | |||||||||||||||
| 无纹多沟藻 | Polykrikos schwartzii | + | + | + | + | ||||||||||
| 原多甲藻类 | |||||||||||||||
| 原多甲藻 | Protoperidinium sp. | Brigantedinium sp. | + | + | + | + | + | ||||||||
| 褐色原多甲藻 | Protoperidinium avellana | Brigantedinium cariacoense | + | + | |||||||||||
| 宽刺原多甲藻 | Protoperidinium latissimum | + | + | ||||||||||||
| 小齿原多甲藻 | Protoperidinium denticulatum | Brigantedinium sp. | + | + | + | + | |||||||||
| 里昂原多甲藻 | Protoperidinium leonis | Lejeunecysta concreta | + | ||||||||||||
| 赛裸原多甲藻 | Protoperidinium subinerme | Selenopemphix nephroides | + | ||||||||||||
| 里昂原多甲藻 | Protoperidinium leonis | Quinquecuspis conretum | + | ||||||||||||
| 锥形原多甲藻 | Protoperidinium conicum | Selenopemphix quanta | + | + | + | + | + | ||||||||
| 原多甲藻 | Protoperidinium sp. | Trinovantedinium capitatum | + | + | + | + | + | + | |||||||
| 长形原多甲藻 | Protoperidinium oblongum | Votadinium calvum | + | + | + | + | + | + | + | + | |||||
| 原多甲藻 | Protoperidinium sp. | Brigantedinium grande | + | ||||||||||||
| 锥腹原多甲藻 | Protoperidinium conicoides | Brigantedinium simplex | + | ||||||||||||
| 分支原多甲藻 | Protoperidinium divaricatum | Xandaradinium xanthum | + | + | + | ||||||||||
| 微小原多甲藻 | Protoperidinium minutum | + | |||||||||||||
| 原多甲藻 | Protoperidinium sp. | + | + | + | + | + | + | + | |||||||
| 未定孢囊 | Unknown cysts | + | + | + | + | + | + | + | + | + | + | + | + | ||
| 甲藻孢囊A型 | + | + | + | ||||||||||||
| 甲藻孢囊B型 | + | + | |||||||||||||
根据Matsuoka(2001)和王朝晖(2007a)文献, 将37种甲藻孢囊分为20种自养型甲藻孢囊和17种异养型甲藻孢囊。其中除S4、S5、S6之外的其他站位中, 膝沟藻属孢囊总丰度明显多于原多甲藻属孢囊; S5站位中异养型甲藻孢囊较丰富; S4和S6站位中, 自养型和异养型孢囊总丰度无明显的区分(表 1)。根据优势度指数, 所有站位甲藻孢囊的优势种均为膝沟藻孢囊(Gonyaulax sp.)。另外, 多边舌甲藻(Lingulodinium polyedra)在除S4之外的其他站位中均有分布。
2.2 甲藻孢囊的丰度分布胶州湾表层沉积物中不同站位之间甲藻孢囊的总丰度存在明显差异, 介于96和969cyst/g DW之间, 平均值为401cyst/g DW(图 2)。其中S13站的孢囊丰度最高, 为969cyst/g DW; 而S5站的孢囊丰度最低, 仅为96cyst/g DW。
 |
| 图 2 胶州湾各站位孢囊丰度分布图 Fig. 2 Total abundance of the cysts at each station in Jiaozhou Bay |
自养型孢囊丰度显著高于异养型丰度。自养丰度(A)介于36和883cyst/g DW之间, 最高值出现在S13站, 为883cyst/g DW, 最低值是S5站为36cyst/g DW, 平均值为312cyst/g DW; 异养丰度(H)介于36和188cyst/g DW之间, 最高值出现在S8站, 为188cyst/g DW, 最低值是S3站, 仅为6cyst/g DW, 平均值为90cyst/g DW。H:A均值仅为0.59, 最大值1.67, 最低值仅为0.1(表 2), 孢囊总丰度呈现临近湾口的站位丰度较高、而湾内较低的现象, 自养孢囊总丰度变化趋势与孢囊总丰度的分布特点相一致, 异养孢囊总丰度无明显变化趋势。孢囊的分布是海流和底质等海洋环境因素共同调控的结果, 其中底部沉积物颗粒的大小对孢囊的分布有显著的影响, 在沉积物粒径小的地方分布多, 粒径大的地方分布较少(石雅君等, 2011)。胶州湾口处为一海底深槽, 水深流急, 冲刷强烈, 沉积物粒度较粗, 而湾口内沉积物粒度总体较细, 湾口外深槽表层为薄层粘土层沉积(肖菲等, 2006), 粒径较小, 有利于孢囊沉积, 加上水深条件及水流的冲刷沉降作用, 使得靠近湾口的位置孢囊丰度较高。
表 2 胶州湾海域甲藻孢囊的多样性指数、均匀度指数和H︰A Tab. 2 Shannon-Wiener index, Pielou evenness index and H︰A of dinoflagellate cysts in Jiaozhou Bay
| 值 | 站位 | |||||||||||
| S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 | S13 | S14 | |
| 多样性指数 | 3.29 | 3.09 | 2.84 | 3.72 | 3.91 | 3.69 | 3.68 | 3.2 | 3.61 | 3.32 | 3.35 | 3.39 |
| 均匀度指数 | 0.95 | 0.89 | 0.89 | 0.98 | 0.86 | 0.85 | 0.88 | 0.89 | 0.88 | 0.83 | 0.75 | 0.89 |
| H:A | 0.55 | 1 | 1.67 | 1 | 0.18 | 0.72 | 0.59 | 0.71 | 0.34 | 0.13 | 0.1 | 0.11 |
| 注: H:异养丰度; A:自养丰度 | ||||||||||||
空间分布上, 靠近河流入海口的站位, 如S8站位, 其孢囊总丰度及异养孢囊丰度相对较高, 其原因可能是沿岸为人口密集区, 陆源输入带来大量营养盐, 初级生产力较高, 而在湾外S13、S14两个站位出现较高的丰度, 其原因可能是该处水深较大, 表层沉积物受水流影响较小, 加之远离海岸, 人为干扰相对较小, 有利于孢囊沉积。其次, 孢囊的沉积还与底质性质、沉积物扰动及底栖动物的摄食有关(王朝晖等, 2014), 因此其数量的空间分布特征还需要进一步结合环境状况进行相关分析。
2.3 甲藻孢囊多样性的空间分布特征丰富度是指物种的多少, 即群落中生物的种类数。各站位的孢囊丰富度为9—23, 其中S5站孢囊丰富度最低为9, S7站孢囊丰富度最高为23。香农-威纳指数通常用于反映该区域群落结构的复杂程度, 越复杂的群落越趋于稳定, 其缓冲作用也越强, 当环境发生改变时, 对环境的反馈功能也越强(刘东艳等, 2003)。胶州湾甲藻孢囊香农-威纳指数介于2.84— 3.91(表 2), 平均值为3.42, 呈现从北向南先增加后减小的趋势。均匀度指数代表群落内部物种分布的均匀程度, 是群落是否成熟和稳定的特征之一(杨世民等, 2014)。胶州湾表层沉积物中甲藻孢囊的均匀度指数为0.75—0.98, 平均值为0.88, 最大值出现在S6站, 为0.98, 最小值出现在S13站, 为0.75, 总体值较高, 表明甲藻群落相对稳定。
2.4 甲藻孢囊与赤潮的关系许多有毒有害甲藻都能形成孢囊, 孢囊被认为是赤潮发生的“种源”和赤潮消亡的重要原因(王朝晖, 2007a)。本次调查结果显示, 胶州湾某些站位发现两种产YTX(Yessotoxin)毒素的甲藻[具刺膝沟藻(Gonyaulax spinifera)和多边舌甲藻(Lingulodinium polyedrum)]的孢囊, 以及3种产麻痹性贝类毒素(PSP)的甲藻[微小/相似亚历山大藻、塔玛/链状亚历山大藻(Alexandrium minutum/affine, A.catenella/tamarense)和链状裸甲藻(Gymnodinium catenatum)]的孢囊。其中具刺膝沟藻孢囊在除S5站之外的其他站位中均有分布, 多边舌甲藻孢囊除S4站的其他站位中均有分布, 亚历山大藻的孢囊主要集中分布在S5, S7, S8和S13站, 而链状裸甲藻的孢囊主要分布在S3, S6及S12站。
作为中国海区常见的甲藻孢囊种类之一, 亚历山大藻孢囊在世界各大海域也广泛分布。亚历山大藻孢囊不仅是赤潮循环发生的种源, 而且其孢囊也是产生麻痹性贝毒素的重要来源(Fertouna-Bellakhal et al, 2015)。在本次采样调查中, 胶州湾海域亚历山大藻孢囊的丰度为4—10cyst/g DW, 平均值为8cyst/g DW, 与其他有PSP问题海域亚历山大孢囊丰度相比(表 3), 近期暴发赤潮的可能性较小, 但由于PSP毒素是危害海水养殖业的重要问题, 在我国沿海许多海域的贝类中检测出PSP毒素, 其中不少地区贝类样品中毒素含量超出国际安全食用标准(韩笑天等, 2004), 因此除了水体中藻类活体细胞的监测, 还应加强对亚历山大藻孢囊的监测与预警。
表 3 各海域亚历山大藻孢囊丰度比较 Tab. 3 Comparison of Alexandrium tamarense / catenella cyst abundance in different sea areas
| 海域 | 甲藻孢囊种类 | 丰度(cyst/g DW) | 文献 |
| 胶州湾 | 塔马亚历山大藻/链状亚历山大藻 | 8 | 本研究 |
| 大鹏湾 | 塔马亚历山大藻/链状亚历山大藻 | 396 | Wang et al, 2004 |
| 大亚湾 | 塔马亚历山大藻/链状亚历山大藻 | 130 | Wang et al, 2004 |
| 长江口 | 塔马亚历山大藻/链状亚历山大藻 | 46 | Wang et al, 2004 |
| 英国东北沿海 | 塔马亚历山大藻/链状亚历山大藻 | 2—404 | 韩笑天等, 2004 |
| 日本濑户内海广岛湾 | 塔马亚历山大藻/链状亚历山大藻 | 50—9525 | 韩笑天等, 2004 |
| 韩国Stait湾 | 塔马亚历山大藻/链状亚历山大藻 | 16—23 | 韩笑天等, 2004 |
本文首次报道了胶州湾表层沉积物中甲藻孢囊种类与数量的分布状况, 共分离鉴定出甲藻孢囊37种和2种未定种, 其中自养型孢囊多于异养型, 膝沟藻孢囊在调查站位中分布广泛。空间分布上, 各个站位间甲藻孢囊丰度存在明显差异, 介于96—969cyst/g DW之间, 呈现出湾口区域比湾内高的分布特征, 今后还需要进一步结合环境因素进行相关分析。另外需要特别注意的是, 本次调查中发现几种产麻痹性贝类毒素(PSP)的甲藻(微小/相似亚历山大藻、塔马/链状亚历山大藻、链状裸甲藻等)孢囊在胶州湾各个站位广泛分布, 尽管其丰度不高, 暴发赤潮的可能性较小, 但仍需要特别关注。
致谢 国家重点基础研究发展规划项目组团队成员对我们样品的采集与处理给予了大力支持与帮助, 在此一并表示感谢。
参考文献
| 王艳, 黄琳, 顾海峰, 等, 2012. 中国渤海海域甲藻孢囊的种类多样性和生态地理分布. 植物学报, 47(2): 125–132 |
| 王朝晖, 2007a. 中国沿海甲藻孢囊与赤潮研究. 北京: 海洋出版社, |
| 王朝晖, 2007b. 中国典型近海海域甲藻孢囊分布及其与富营养化和赤潮生消关系研究. 广州: 暨南大学博士学位论文 |
| 王朝晖, MatsuokaK, 齐雨藻, 等, 2003. 有毒亚历山大藻(Alexandrium spp.)和链状裸甲藻(Gymnodinium catenatum)孢囊在中国沿海的分布.. 海洋与湖沼, 34(4): 422–430 |
| 王朝晖, 康伟, 2014. 柘林湾表层沉积物中甲藻孢囊的分布与浮游植物休眠体萌发研究. 环境科学学报, 34(8): 2043–2050 |
| 石雅君, 刘东艳, 邵红兵, 等, 2011. 北黄海表层沉积物中甲藻孢囊的分布特征. 海洋通报, 30(3): 320–327 |
| 卢敏, 张龙军, 李超, 等, 2001. 1999年7月胶州湾东部赤潮生消过程生态环境要素分析. 黄渤海海洋, 19(4): 43–50 |
| 刘东艳, 孙军, 陈洪涛, 等, 2003. 2001年夏季胶州湾浮游植物群落结构的特征. 青岛海洋大学学报, 33(3): 366–374 |
| 孙爱梅, 李超, 蓝东兆, 等, 2006. 罗源湾口柱状沉积物中的甲藻孢囊. 台湾海峡, 25(1): 10–18 |
| 杨世民, 王丽莎, 石晓勇, 2014. 2009年春季胶州湾浮游植物群落结构特征. 海洋与湖沼, 45(6): 1234–1240 |
| 肖菲, 尹延鸿, 2006. 青岛海岸带及邻近海域地形和沉积物类型研究. 海洋地质动态, 22(10): 1–5 DOI:10.3969/j.issn.1009-2722.2006.10.001 |
| 肖咏之, 齐雨藻, 王朝晖, 等, 2001. 大亚湾海域锥状斯氏藻赤潮及其与孢囊的关系. 海洋科学, 25(9): 50–54 |
| 顾海峰, 刘婷婷, 蓝东兆, 2011. 中国沿海甲藻包囊研究进展. 生物多样性, 19(6): 779–786 |
| 黄海燕, 陆斗定, 2009. 甲藻孢囊研究进展. 海洋学研究, 27(3): 85–92 |
| 韩笑天, 邹景忠, 张永山, 2004. 胶州湾赤潮生物种类及其生态分布特征. 海洋科学, 28(2): 49–54 |
| Fertouna-Bellakhal M, Dhib A, Fathalli A, et al, 2015. Alexandrium pacificum Litaker sp. nov (Group Ⅳ):Resting cyst distribution and toxin profile of vegetative cells in Bizerte Lagoon (Tunisia, Southern Mediterranean Sea). Harmful Algae, 48: 69–82 |
| Matsuoka K, 2001. Further evidence for a marine dinoflagellate cyst as an indicator of eutrophication in Yokohama Port, Tokyo Bay, Japan:Comments on a discussion by B. Dale. Science of the Total Environment, 264(3): 221–233 DOI:10.1016/S0048-9697(00)00718-X |
| Matsuoka K, Head M J, 2013. Clarifying cyst-motile stage relationships in dinoflagellates. In:Lewis J M, Marret F, Bradley L eds. Biological and Geological Perspectives of Dinoflagellates. London:The Micropalaeontological Society, Special Publications Geological Society, 325-348 |
| Pielou E C, 1969. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, |
| Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana IL: University of Illinois Press, |
| Wang Z H, Matsuoka K, Qi Y Z, et al, 2004. Dinoflagellate cysts in recent sediments from Chinese coastal waters. Marine Ecology, 25(4): 289–311 DOI:10.1111/mae.2004.25.issue-4 |