 2017, Vol. 48
2017, Vol. 48中国海洋湖沼学会主办。
文章信息
- 金丹璐, 张清科, 王友发, 朱咏梅, 张玉明, 王建平, 竺俊全. 2017.
- JIN Dan-Lu, ZHANG Qing-Ke, WANG You-Fa, ZHU Yong-Mei, ZHANG Yu-Ming, WANG Jian-Ping, ZHU Jun-Quan. 2017.
- 鲤科经济鱼类马口鱼(Opsariichthys bidens)胚胎发育及仔稚鱼形态与生长观察研究
- OBSERVATION OF EMBRYONIC, LARVA AND JUVENILE DEVELOPMENT OF OPSARIICHTHYS BIDENS
- 海洋与湖沼, 48(4): 838-847
- Oceanologia et Limnologia Sinica, 48(4): 838-847.
- http://dx.doi.org/10.11693/hyhz20170200034
-
文章历史
- 收稿日期:2017-02-20
- 收修改稿日期:2017-03-16




2. 浙江省新昌县水产技术推广中心 新昌 311300;
3. 宁波市海洋与渔业研究院 宁波 315012
2. Fisheries Technical Extension Center of Xinchang County, Xinchang 311300, China;
3. Ningbo Academy of Oceanology and Fisheries, Ningbo 315012, China
鱼类胚胎及仔稚鱼发育是早期生活史研究的重要方面。对经济鱼类胚胎及仔稚鱼发育特点及规律的了解, 有助于指导育苗生产。目前有关鲤科经济鱼类胚胎及仔稚鱼发育的研究较多, 已见鲃亚科的宽口光唇鱼(Acrossocheilus monticola)(严太明等, 1999)、中华倒刺鲃(Spinibarbus sinensis)(黄洪贵, 2009a)、倒刺鲃(Spinibarbus denticulatus)(易祖盛等, 2004; 黄洪贵, 2009b)、大鳞鲃(Barbus capito)(Houman et al, 2002; 徐伟等, 2011)及光唇鱼(Acrossocheilus fasciatus)(姜建湖等, 2012), 鲌亚科的团头鲂(Megalobarama amblycephala)(李军等, 1993)、厚颌鲂(Megalobrama pellegrini)(王剑伟等, 2005)及翘嘴红鲌(Erythroculter ilishaeformis)(顾志敏等, 2008), 鮈亚科的稀有鮈鲫(Gobiocypris rarus)(常剑波等, 1995), 裂腹鱼亚科的齐口裂腹鱼(Schizothorax prenanti)(吴青等, 2004)、细鳞裂腹鱼(Schizothorax chongi)(陈礼强等, 2008)及四川裂腹鱼[Schizothorax (Racoma) kozlovi] (He et al, 2014)等种类的研究报道。
马口鱼(Opsariichthys bidens)属鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鮈亚科(Danioninae), 多生活于溪流及水库等清澈水体中, 尤其喜在水流较急的浅滩、底质为砂石的小溪和江河支流中集群生活, 以小鱼、小虾、水生昆虫等为食(金柏等, 1985); 马口鱼肉质与品味具佳, 深受消费者青睐。马口鱼成鱼体长一般10—20cm, 体重50—150g, 体银灰略带红色, 雄鱼具蓝色横纹、色彩艳丽, 具一定观赏价值。马口鱼繁殖力强、生长快、病害少, 人工养殖条件下喂食配合饲料生长良好, 是一种适合在库叉、溪流及河道中放养, 也适合山区养殖场利用溪流水、泉水、水库水进行设施养殖及非饮用型水库中进行网箱养殖, 以及游钓业开发的特色经济鱼类。近年来, 马口鱼自然资源因过度利用及栖息地水环境污染等原因而衰退, 因此人工育苗及增养殖开发引起重视。目前, 有关马口鱼的研究已见生殖力(李强等, 2010)、人工繁殖和养殖(姜景田, 2010)、消化道结构(母丹利等, 2016)与食性(姜志强等, 1995)、年龄与生长(李强等, 2007)、遗传(Johansson, 2006; Perdices et al, 2005)、分类(陈宜瑜, 1982)等方面, 而马口鱼早期生活史的研究欠缺, 胚胎及仔稚鱼发育的研究未见报道。本研究在人工培育条件下观察了马口鱼胚胎发育过程、仔稚鱼形态变化与生长特性, 旨在了解马口鱼胚胎及仔稚鱼发育特点及规律, 积累马口鱼早期生活史资料, 为马口鱼人工繁殖、鱼苗培育及养殖提供理论参考。
1 材料与方法 1.1 材料实验用鱼为浙江省宁波市鄞州盛龙特种水产养殖场培育的人工繁殖用马口鱼亲鱼, 雌鱼体重35— 50g, 雄鱼体重75—100g, 共200余尾, 雌雄比1 : 1, 暂养于面积为60m2的水泥池中, 供实验用。
1.2 方法 1.2.1 人工授精、孵化及仔稚鱼培育2014年7月, 挑选经人工催产后发情的亲鱼, 用人工干法授精获得受精卵24000余粒; 将受精卵散布于孵化框底(大小60cm×50cm×5cm, 底部为60目筛网, 共5个), 置于孵化池(大小2m×2m×0.6m的水泥池)水体表层, 在水温26—28℃, 微流水、微充气、遮光条件下孵育; 仔鱼孵出后保持水温26—28℃, 在流水、充气、避直射光条件下培育; 仔鱼开口后投喂活体枝角类、桡足类。受精卵孵化、仔稚鱼培育用水为洁净的地下水, 水质符合淡水养殖标准。
1.2.2 胚胎发育观察及有效积温的推算于孵化框中定期随机取样40—50粒/次, 于Leica DM1000显微镜下观察胚胎发育过程, 用Panasonic DMC-ZS15数码相机对胚胎发育过程进行显微拍照。发育期的划分以观察胚胎数的50%以上显示某发育期的特征为准。胚胎发育过程的有效积温推算:
K = N (T−C)
其中, K为有效积温, N为发育所需时间, T为生境温度值, C为生物学零度, 以此计算出马口鱼胚胎发育生物学零度和受精卵发育至仔鱼孵出的有效积温(℃·d)。
生物学零度与发育所需有效积温的计算公式如下:
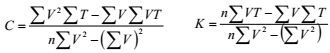
其中, V=1/N; n为数据组数(董根等, 2013)。
1.2.3 仔稚鱼发育观察及生长测量定期随机取样28—32尾/次, 在Leica DM1000型显微镜与Novel NTB-2B型解剖镜下观察不同发育阶段仔稚鱼形态特性, 并用Panasonic DMC-ZS15数码相机拍照; 利用目微尺测量各发育阶段仔稚鱼的全长、肛前距、头长、眼径、体高和卵黄囊尺寸。
1.2.4 数据处理仔稚鱼测量数据用Microsoft Office Excel 2010和SPSS 13.0进行处理, 结果以平均值±标准差(X±SD)表示。
2 结果 2.1 胚胎发育马口鱼的成熟卵呈圆形、黄色, 比重大于水, 属沉性卵, 卵径1.00—1.20mm。受精后遇水迅速吸水膨胀, 直径达1.4—1.6mm, 沉性, 但不具黏性(图 1: 1)。在水温26—28℃下, 受精后37h孵化出仔鱼。根据马口鱼胚胎发育过程的形态特征变化, 按胚盘形成期、卵裂期、囊胚期、原肠胚期、神经胚期、胚孔封闭期、器官发生期、心跳期和孵出期等阶段进行描述(表 1)。
| 发育阶段 | 发育时间 | 积温(℃·h) | 图版序号 | |
| 受精卵 | 0h | 0.00 | Ⅰ-1 | |
| 胚盘形成期 | 30min | 13.39 | Ⅰ-2 | |
| 卵裂期 | ||||
| 2细胞期 | 1h | 26.79 | Ⅰ-3 | |
| 4细胞期 | 1h 30min | 40.18 | Ⅰ-4 | |
| 8细胞期 | 1h 50min | 49.11 | Ⅰ-5 | |
| 16细胞期 | 2h | 53.58 | Ⅰ-6 | |
| 32细胞期 | 2h 20min | 62.51 | Ⅰ-7 | |
| 64细胞期 | 2h 40min | 71.43 | Ⅰ-8 | |
| 多细胞期 | 3h 55min | 104.92 | Ⅰ-9 | |
| 囊胚期 | ||||
| 囊胚早期 | 4h 50min | 127.54 | Ⅰ-10 | |
| 囊胚中期 | 6h 20min | 167.12 | Ⅰ-11 | |
| 囊胚晚期 | 7h 15min | 191.31 | Ⅰ-12 | |
| 原肠胚期 | ||||
| 原肠早期 | 8h 35min | 223.06 | Ⅰ-13 | |
| 原肠中期 | 10h | 259.88 | Ⅰ-14 | |
| 原肠晚期 | 11h 50min | 303.19 | Ⅰ-15 | |
| 神经胚期 | 13h 30min | 351.84 | Ⅰ-16 | |
| 胚孔封闭期 | 15h 5min | 393.62 | Ⅰ-17 | |
| 器官发生期 | ||||
| 肌节出现期 | 16h 20min | 426.61 | Ⅰ-18 | |
| 眼基期 | 18h 5min | 487.12 | Ⅰ-19 | |
| 眼囊期 | 18h 25min | 496.18 | Ⅰ-20 | |
| 尾芽期 | 19h 50min | 540.60 | Ⅰ-21 | |
| 尾鳍褶出现期 | 21h 30min | 592.80 | Ⅰ-22 | |
| 肌肉效应期 | 24h 5min | 662.34 | Ⅰ-23 | |
| 眼晶体出现期 | 27h | 733.01 | Ⅰ-24 | |
| 心脏原基期 | 29h 20min | 770.33 | Ⅰ-25 | |
| 耳石出现期 | 31h 50min | 832.13 | Ⅰ-26 | |
| 心跳期 | 33h 40min | 855.47 | Ⅰ-27 | |
| 孵出期 | 37h | 968.95 | Ⅰ-28 |
 |
| 图 1 马口鱼胚胎发育 Fig. 1 Embryonic development of O. bidens 注: 1.受精卵; 2.胚盘形成期; 3. 2细胞期; 4. 4细胞期; 5. 8细胞期; 6. 16细胞期, 顶面观; 7. 32细胞期, 顶面观; 8. 64细胞期, 顶面观; 9.多细胞期; 10.囊胚早期; 11.囊胚中期; 12.囊胚晚期; 13.原肠早期; 14.原肠中期; 15.原肠晚期; 16.神经胚期; 17.胚孔封闭期; 18.肌节出现期, 示肌节(→); 19.眼基期, 示眼原基(→); 20.眼囊期, 示眼囊(→); 21.尾芽期, 示尾芽(→); 22.尾鳍褶出现期, 示尾鳍褶(→); 23.肌肉效应期; 24.眼晶体出现期, 示眼晶体(→); 25.心脏原基期, 示心脏原基(→); 26.耳石出现期, 示耳石(→); 27.心跳期, 示心脏(→); 28.孵出期 |
受精后30min, 卵原生质朝动物极端流动, 逐渐隆起形成胚盘, 约20min后, 隆起的高度约为受精卵直径的1/4(图 1: 2)。
2.1.2 卵裂期受精后1h, 胚盘经裂为2个大小相同的分裂球(图 1: 3); 30min后, 再次经裂为四个等大的分裂球(图 1: 4), 前两次卵裂的经裂面互相垂直; 20min后, 第三次经裂, 8个分裂球排列成两行, 每行4个分裂球(图 1: 5); 10min后, 第四次经裂, 16个分裂球呈四行排列, 每行4个分裂球(图 1: 6); 20min后, 第五次分裂, 形成32个分裂球(图 1: 7)。受精后2h 40min, 分裂形成64个分裂球(图 1: 8)。受精后3h 55min, 卵裂球明显变小, 界限不清, 形成多层排列的细胞团, 为多细胞期(图 1: 9)。
2.1.3 囊胚期随着卵裂的继续进行, 分裂球变得更小。受精后4h 50min, 卵黄囊上方由分裂球构成的囊胚层隆起似高帽状, 高度为卵黄囊高度的1/4—1/3, 为囊胚早期(图 1: 10); 1h 30min后, 囊胚层的高度明显降低, 为囊胚中期(图 1: 11); 受精后7h 15min, 囊胚层的高度降低至接近卵黄囊外周高度, 为囊胚晚期(图 1: 12)。
2.1.4 原肠胚期囊胚层细胞继续沿卵黄囊外周向植物极端下包。受精后8h 35min, 囊胚层细胞下包至卵黄高度约1/3处, 胚环形成, 为原肠早期(图 1: 13)。受精后10h, 囊胚层细胞下包至卵黄高度约2/3处, 胚环端显著增厚, 并隆起形成胚盾, 为原肠中期(图 1: 14)。受精后11h 50min, 囊胚层细胞下包至卵黄高度约4/5处, 胚孔形成, 为原肠晚期(图 1: 15)。
2.1.5 神经胚期受精后13h 30min, 囊胚层细胞下包至几乎完全包裹卵黄, 仅露出细小的卵黄拴, 胚胎的神经板雏形出现(图 1: 16)。
2.1.6 胚孔封闭期受精后15h 5min, 胚孔封闭, 卵黄囊背面形成弧形的胚体(图 1: 17)。
2.1.7 器官发生期受精后16h 20min, 胚体包绕卵黄囊外周的4/5, 中部4—6对肌节形成, 每个肌节如“(”状, 为肌节出现期(图 1: 18); 受精后18h 5min, 胚体前端近头部的左右两侧椭圆形的眼原基形成, 此时肌节有8—10对, 为眼基期(图 1: 19); 15min后, 眼基中部下凹形成眼囊, 为眼囊期(图 1: 20); 受精后19h 50min, 胚体尾部出现一个芽状突起, 称为尾芽, 为尾芽期(图 1: 21); 受精后21h 30min, 随着尾芽的生长, 其边缘外凸形成尾鳍褶, 此时尾部与头部之间靠近尾部的卵黄囊向内凹陷, 此为尾鳍褶出现期(图 1: 22), 肌节数明显增多, 形状变为“ < ”形; 受精后24h 5min, 胚体肌肉出现微弱收缩, 随着收缩频率的提高, 胚体开始左右扭动, 受精后25h 30min, 胚体尾鳍已经发生, 随着胚体肌肉运动, 卵黄囊逐渐变小, 为肌肉效应期(图 1: 23); 受精后27h, 眼囊逐渐变圆, 囊内具圆形晶体, 为眼晶体出现期(图 1: 24)。受精后29h 20min, 胚体头部与卵黄囊连接处出现心脏原基, 为心脏原基期(图 1: 25), 卵黄囊进一步变小; 受精后31h 50min, 耳囊结构明显, 内部有2粒耳石, 为耳石出现期(图 1: 26)。
2.1.8 心跳期受精后33h 40min, 胚体出现心跳, 心跳频率为约60次/min, 血液在卵黄囊与胚体之间流动, 为心跳期(图 1: 27), 胚体尾部扭动频繁。
2.1.9 孵出期受精后37h, 胚体扭动加剧, 开始出膜, 大部分胚体尾部先出膜(图 1: 28)。
2.2 仔稚鱼发育 2.2.1 早期仔鱼初孵仔鱼, 全长3.74±0.04mm, 头部及尾部与卵黄囊分离, 躯干部贴于卵黄囊上; 卵黄囊前半部呈椭圆形, 后半部呈棒状; 心脏由心房、心室和静脉窦组成, 内部血液流动明显; 肛门原基形成, 尾椎骨斜向上翘(图 2: 1, 1-a, 1-b)。仔鱼活动能力弱, 一般静卧不动, 偶见其短暂运动。孵后12h的仔鱼, 全长5.41±0.25mm, 卵黄囊变小, 眼后下方可见5对鳃弧, 其内血液流动明显, 胸鳍原基明显可见(图 2: 2, 2-a, 2-b)。孵后24h的仔鱼, 全长5.62±0.06mm, 卵黄囊继续变小, 耳囊中耳石上方可见弧形半规管, 眼内黑色素开始积累, 心脏中红色血液流动明显(图 2: 3, 3-a, 3-b)。孵后32h的仔鱼, 全长5.64±0.22mm, 眼中黑色素增多, 鳃弧更加明显, 血液红色加深(图 2: 4, 4-a, 4-b)。孵后40h的仔鱼, 全长6.03±0.14mm, 卵黄囊明显变小呈棒状, 眼内布满黑色素, 躯体部与卵黄囊相接处出现一列枝状黑色素(图 2: 5, 5-a, 5-b)。孵后48h的仔鱼, 全长6.20±0.19mm, 头部出现黄色点状斑, 口裂明显, 胸鳍鳍条形成; 枝状黑色素增多, 肠道明显(图 2: 6, 6-a, 6-b)。孵后60h的仔鱼, 全长6.25±0.11mm, 肠道更加明显; 肛门至尾鳍间可见一列枝状黑色素, 可见肾脏原基(图 2: 7, 7-a, 7-b)。孵后72h的仔鱼, 全长6.40±0.22mm, 口裂和鳃盖已能开合。孵后84h的仔鱼, 全长6.45±0.08mm, 头部及躯体部黑色素分布增多, 鳔一室清晰可见(图 2: 8, 8-a, 8-b), 少数仔鱼开始平游。孵后96h的仔鱼, 全长6.49±0.19mm, 卵黄囊很小, 头部与躯干部背面呈浅黄绿色, 肛门与外界相通(图 2: 9, 9-a, 9-b), 多数仔鱼能平游。
 |
| 图 2 马口鱼仔稚鱼发育(Ⅰ) Fig. 2 Larval and juvenile development of O. bidens (Ⅰ) 注: 1.初孵仔鱼, 1-a.示肛门原基, 1-b.示尾椎骨; 2.孵后12h仔鱼, 2-a.示鳃弧, 2-b.示胸鳍原基; 3.孵后24h仔鱼, 3-a.示半规管, 3-b.示心脏内的血液; 4.孵后32h仔鱼, 4-a.示眼, 4-b.示心脏内的血液; 5.孵后40h仔鱼, 5-a.示眼, 5-b.示枝状黑色素; 6.孵后48h仔鱼, 6-a.示鳃裂, 6-b.示肠道; 7.孵后60h仔鱼, 7-a.示肠道, 7-b.示肾脏原基; 8.孵后84h仔鱼, 8-a.示鳔1室, 8-b.示躯体部黑色素; 9.孵后96h仔鱼, 9-a.示鱼体背部皮肤, 9-b.示肛门 |
孵后108h的仔鱼, 全长达6.52±0.18mm, 卵黄囊几乎消失, 肠道内可见食物, 营外源性营养(图 3: 10, 10-a, 10-b)。孵后5d的仔鱼, 全长6.69±0.21mm, 尾鳍鳍条8—10条, 黑色素分布继续增多, 鳔一室, 肠道蠕动频繁(图 3: 11, 11-a, 11-b)。孵后6d仔鱼, 全长6.88±0.19mm, 鳔仍一室, 肠道内充满食物, 尾鳍鳍条增至18条(图 3: 12, 12-a, 12-b)。孵后10d仔鱼, 全长8.27±0.28mm, 吻端、头背部及体侧黑色素增多, 尾鳍黄色点状色素分布明显(图 3: 13, 13-a, 13-b)。孵后14d仔鱼, 全长9.94±0.91mm, 鳔两室, 前室小于后室, 可见背鳍(图 3: 14, 14-a, 14-b)。孵后16d仔鱼, 各鳍发育已近完成(图 3: 15, 15-a, 15-b)。
 |
| 图 3 马口鱼仔稚鱼发育(Ⅱ) Fig. 3 Larval and juvenile development of O. bidens (Ⅱ) 注: 10.孵后108h仔鱼, 10-a.示肠道内食物, 10-b.示肛门; 11.孵后5d仔鱼, 11-a.示尾鳍鳍条, 11-b.示黑色素; 12.孵后6d仔鱼, 12-a.示肠道内食物, 12-b.示尾鳍; 13.孵后10d龄仔鱼, 13-a.示口裂处的黑色素, 13-b.示尾鳍上黄色点状色素; 14.孵后14d仔鱼, 14-a.示两鳔室, 14-b.示背鳍; 15.孵后16d仔鱼, 15-a.示臀鳍鳍条, 15-b.示腹鳍鳍条; 16.孵后17d稚鱼, 16-a.示鳞片 |
孵后17d稚鱼, 全长12.46±0.76mm, 各鳍发育完整, 腹鳍之后的体侧开始出现鳞片(图 3: 16, 16-a), 进入稚鱼期。孵后23d稚鱼, 全长达17.09±1.08mm, 外部形态特征与成鱼相近。
2.3 仔稚鱼生长特性经回归分析, 仔稚鱼的全长生长方程为: y= 0.0169x2+0.1031x+5.4831 (R2=0.9696) (y为全长, x为孵后时间)(图 4)。仔鱼孵化后12h内全长生长最快, 尤其是肛前距的生长; 仔鱼孵化后12h—6d内全长生长较慢, 肛前距的生长也如此。孵后6d之后的仔稚鱼全长生长加快, 肛前距的生长也加快; 孵后24—40h仔鱼头长生长较快, 孵后40—6d仔鱼头长生长较慢, 孵后6d之后的仔稚鱼头长生长加快。孵后0—12h仔鱼眼径增长较快, 孵后12—6d仔鱼眼径增长较慢, 孵后6—23d仔稚鱼眼径增长加快; 孵后24—6d仔鱼体高增长较慢, 孵后6—23d仔稚鱼体高增长加快(表 2)。
| 发育阶段 | 孵出时间 | 样本数 | 全长(mm) | 肛前长(mm) | 头长(mm) | 眼径(mm) | 体高(mm) | 卵黄囊长(mm) |
| 早期仔鱼 | 0h | 10 | 3.74±0.04 | 2.79±0.04 | — | 0.24±0.01 | — | 2.53±0.04 |
| 12h | 10 | 5.41±0.25 | 3.62±0.15 | — | 0.35±0.02 | — | 2.84±0.14 | |
| 24h | 10 | 5.62±0.06 | 3.67±0.15 | 0.67±0.04 | 0.37±0.02 | 0.80±0.08 | 2.76±0.13 | |
| 32h | 10 | 5.64±0.22 | 3.69±0.17 | 0.92±0.02 | 0.37±0.02 | 0.81±0.03 | 2.69±0.10 | |
| 40h | 9 | 6.03±0.14 | 3.86±0.08 | 1.03±0.06 | 0.39±0.04 | 0.82±0.07 | 2.66±0.08 | |
| 48h | 6 | 6.20±0.19 | 3.92±0.11 | 1.04±0.02 | 0.44±0.02 | 0.83±0.01 | 2.64±0.10 | |
| 60h | 10 | 6.25±0.11 | 3.97±0.11 | 1.08±0.07 | 0.44±0.02 | 0.83±0.02 | 2.62±0.08 | |
| 72h | 10 | 6.40±0.22 | 3.99±0.16 | 1.09±0.04 | 0.46±0.02 | 0.84±0.02 | 2.45±0.08 | |
| 84h | 10 | 6.45±0.08 | 4.02±0.06 | 1.11±0.24 | 0.46±0.02 | 0.84±0.03 | 2.30±0.05 | |
| 96h | 10 | 6.49±0.19 | 4.04±0.13 | 1.13±0.04 | 0.46±0.02 | 0.84±0.04 | 1.67±0.19 | |
| 晚期仔鱼 | 108h | 10 | 6.52±0.18 | 4.05±0.11 | 1.20±0.03 | 0.46±0.02 | 0.84±0.03 | — |
| 120h | 10 | 6.69±0.21 | 4.14±0.11 | 1.22±0.03 | 0.48±0.02 | 0.85±0.01 | — | |
| 6d | 10 | 6.88±0.19 | 4.18±0.10 | 1.26±0.03 | 0.49±0.01 | 0.87±0.02 | — | |
| 8d | 10 | 7.00±0.28 | 4.19±0.18 | 1.31±0.06 | 0.56±0.03 | 0.91±0.07 | — | |
| 10d | 10 | 8.27±0.28 | 4.68±0.18 | 1.60±0.06 | 0.58±0.03 | 1.02±0.07 | — | |
| 12d | 10 | 9.21±0.45 | 5.58±0.31 | 1.89±0.09 | 0.65±0.03 | 1.42±0.10 | — | |
| 16d | 10 | 9.94±0.91 | 6.06±0.51 | 2.16±0.27 | 0.73±0.08 | 1.83±0.23 | — | |
| 稚鱼 | 17d | 9 | 12.46±0.76 | 7.39±0.47 | 2.83±0.21 | 0.90±0.04 | 2.34±0.12 | — |
| 19d | 9 | 13.46±1.11 | 7.61±0.58 | 3.08±0.30 | 0.95±0.06 | 2.52±0.34 | — | |
| 23d | 6 | 17.09±1.08 | 9.51±0.72 | 4.07±0.35 | 1.16±0.06 | 3.40±0.29 | — |
 |
| 图 4 马口鱼仔稚鱼的生长 Fig. 4 Growth of larva and juvenile of O. bidens |
鱼类卵子大小因物种不同存在差异, 同种因亲鱼不同、环境变化及产卵时间不同, 卵子大小也有差异(Bobe et al, 2010), 通常个体较大的亲鱼所产卵也较大(DeMartini, 1991)。马口鱼卵子(卵径1.00— 1.20mm), 比同亚科的唐鱼(Tanichthys albonubes) (刘汉生等, 2008)卵子(卵径0.86—0.89mm)及鲌亚科的翘嘴红鲌卵子(卵径0.72—1.18mm)(顾志敏等, 2008)稍大, 比鲃亚科的宽口光唇鱼卵子(卵径1.55—2.11mm) (严太明等, 1999)、光唇鱼卵子(卵径1.55—2.11mm) (姜建湖等, 2012)、半刺厚唇鱼[Acrossocheilius (Lissochilichthys) hemispinus]卵子(卵径1.74—1.97mm)(陈熙春, 2013)、中华倒刺鲃卵子(卵径1.8—2.0mm)(黄洪贵, 2009a)、倒刺鲃卵子(卵径2.87—3.12mm)(易祖盛等, 2004)、黑棘倒刺鲃[Spinibarbus caldwelli (Nichols)]卵子(卵径1.9—2.1mm)(苏敏等, 2002)及大鳞鲃卵子(卵径1.5mm)(徐伟等, 2011), 鮈亚科的稀有鮈>鲫卵子(卵径1.25—1.70mm)(常剑波等, 1995)及泰山赤鳞鱼卵子(卵径1.65—1.82mm)(陈红菊等, 2008), 裂腹鱼亚科的齐口裂腹鱼卵子(卵径2.9—3.0mm)(吴青等, 2004)和细鳞裂腹鱼卵子(卵径2.6—3.0mm)(陈礼强等, 2008)小。卵的大小还受亲鱼营养水平、产卵批次等的影响(Hsiao et al, 1994; Tyler et al, 1994), 繁殖之前对亲鱼进行营养强化培育, 同时控制产卵批次, 可获得较大的优质卵。卵的大小是影响早期发育的主要因素之一, 仔鱼初次摄食及卵黄耗尽时间与卵径大小呈正相关(Knutsen et al, 1985)。马口鱼卵径较小, 初次摄食及卵黄耗尽时间较早, 育苗生产上应在仔鱼孵后第4d开始投喂开口饵料, 确保开口仔鱼能及时摄食。马口鱼受精卵沉性、无粘性, 孵化采用了姜建湖等(2012)孵化光唇鱼受精卵的方式, 即将受精卵散布于孵化框中, 平放于孵化池水体表层, 在微流水、微充气条件下孵化, 取得了较高孵化率。
马口鱼胚胎发育过程中未见克氏囊(尾泡), 与倒刺鲃(易祖盛等, 2004)、厚颌鲂(王剑伟等, 2005)、齐口裂腹鱼(吴青等, 2004)及唐鱼(陈国柱等, 2004)等相似, 而宽口光唇鱼(严太明等, 1999)及云南光唇鱼(唐安华等, 1982)等的胚胎发育过程中有该器官的形成。马口鱼胚胎眼晶体出现在肌肉效应期之后、心脏原基期之前, 此现象也存在于黑脊倒刺鲃(苏敏等, 2002)、齐口裂腹鱼(吴青等, 2004)、厚颌鲂(王剑伟等, 2005)及光唇鱼(姜建湖等, 2012)等的胚胎发育过程中, 而稀有鮈鲫(常剑波等, 1995)、倒刺鲃(易祖盛等, 2004)、唐鱼(陈国柱等, 2004)及细鳞裂腹鱼(陈礼强等, 2008)等的胚胎眼晶体出现在肌肉效应期之前。马口鱼胚胎耳石出现在心脏原基期之后, 与厚颌鲂(王剑伟等, 2005)及倒刺鲃(易祖盛等, 2004)等种类相似, 而黑脊倒刺鲃(苏敏等, 2002)、泰山赤鳞鱼(陈红菊等, 2008)及光唇鱼(姜建湖等, 2012)胚胎在眼晶体出现期耳囊中就可见耳石。
马口鱼仔鱼发育经历早期仔鱼和晚期仔鱼两阶段, 早期仔鱼无摄食能力, 以卵黄囊内的卵黄为发育所需营养, 亦称之为卵黄囊期仔鱼; 卵黄囊消失之后, 进入晚期仔鱼阶段, 主要进行器官发生与发育。马口鱼仔鱼孵后84h一鳔室形成, 逐渐开始平游, 仔鱼孵后96h大多数开始平游, 这比光唇鱼(姜建湖等, 2012)早, 比黑脊倒刺鲃(唐良华等, 2006)、翘嘴红鲌(顾志敏等, 2008)等晚。马口鱼仔鱼孵后108h卵黄囊几乎消失, 其肠道内可见食物, 即已经开始摄食, 比瓣结鱼(谢恩义等, 2002)、黑脊倒刺鲃(唐良华等, 2006)、细鳞裂腹鱼(陈礼强等, 2008)及光唇鱼(姜建湖等, 2012)开口摄食时间早, 仔鱼在短时间内具有摄食能力, 可快速获取饵料, 以便在繁殖期内取得摄食优势, 提高生存能力。马口鱼人工育苗生产中, 应及时投喂适口饵料, 以提高仔鱼培育的成活率, 本研究以小型活体枝角类及桡足类为开口饵料, 仔鱼培育成活率较高。仔鱼完成运动器官的发育后开始进入稚鱼期, 主要进行鱼体生长、性腺发生与分化。
马口鱼仔稚鱼的生长有阶段性差异, 孵后12h内卵黄囊消耗大, 生长最快; 孵后12h—6d自身卵黄囊为营养(4d内)转向外源营养初期(4—6d)生长较慢; 孵后6d之后已适应从外界摄取饵料为营养, 生长又加快。据研究, 光唇鱼晚期仔鱼生长速度 > 早期仔鱼 > 稚鱼(姜建湖等, 2012), 与马口鱼类似, 均显示早期发育异速生长特性。
4 结论马口鱼卵较小, 初次摄食及卵黄耗尽时间较早。受精卵在26—28℃水温下孵育, 37h孵化出仔鱼。胚胎发育过程中未见克氏囊(尾泡), 眼晶体出现在肌肉效应期之后、心脏原基期之前, 耳石出现在心脏原基期之后。仔鱼孵后96h大多数开始平游, 孵后108h开始摄食。仔稚鱼生长方程为: y=0.0169x2+0.1031x+ 5.4831 (R2=0.9696) (y为全长, x为孵后时间), 显示异速生长特性。本研究积累了马口鱼早期生活史资料, 为马口鱼人工繁殖及鱼苗培育提供理论参考。
| 王剑伟, 谭德清, 李文静, 2005. 厚颌鲂人工繁殖初报及胚胎发育观察. 水生生物学报, 29(2): 130–136 |
| 母丹利, 袁思平, 薛聪顺, 等, 2016. 马口鱼消化道的形态及组织学结构特征. 宁波大学学报(理工版), 29(3): 39–44 |
| 刘汉生, 易祖盛, 林小涛, 2008. 唐鱼的繁殖行为和胚胎发育研究. 水生态学杂志, 29(2): 22–27 |
| 严太明, 何学福, 贺吉胜, 1999. 宽口光唇鱼胚胎发育的研究. 水生生物学报, 23(6): 636–640 |
| 苏敏, 林丹军, 尤永隆, 2002. 黑脊倒刺鲃胚胎发育的观察. 福建师范大学学报(自然科学版), 18(2): 80–84 |
| 李军, 张海明, 1993. 团头鲂胚胎及仔稚鱼发育观察. 水产科技情报, 20(4): 158–163 |
| 李强, 李小芳, 蓝昭军, 等, 2007. 北江马口鱼的年龄、生长和生活史类型的研究. 华南农业大学学报(自然科学版), 28(2增): 78–88 |
| 李强, 蓝昭军, 赵俊, 等, 2010. 广东北江马口鱼个体生殖力研究. 四川动物, 29(3): 440–445 |
| 吴青, 王强, 蔡礼明, 等, 2004. 齐口裂腹鱼的胚胎发育和仔鱼的早期发育. 大连水产学院学报, 19(3): 218–221 |
| 陈礼强, 吴青, 郑曙明, 等, 2008. 细鳞裂腹鱼胚胎和卵黄囊仔鱼的发育. 中国水产科学, 15(6): 927–934 |
| 陈红菊, 姜运良, 宋憬愚, 等, 2008. 泰山赤鳞鱼胚胎发育的研究. 水生生物学报, 32(6): 926–933 |
| 陈国柱, 方展强, 马广智, 2004. 唐鱼胚胎发育观察. 中国水产科学, 11(6): 489–496 |
| 陈宜瑜, 1982. 马口鱼类分类的重新整理. 海洋与湖沼, 13(3): 293–299 |
| 陈熙春, 2013. 半刺厚唇鱼胚胎与胚后发育观察. 福建水产, 35(3): 181–186 |
| 易祖盛, 陈湘粦, 王春, 等, 2004. 倒刺鲃胚胎发育的研究. 中国水产科学, 11(1): 65–69 |
| 金柏, 金克伟, 1985. 铁甲水库马口鱼生物学的初步研究. 水产科学, 4(4): 8–12 |
| 姜志强, 王喜庆, 刘建, 等, 1995. 碧流河水库马口鱼的食性及渔业对策. 水产科学, 14(3): 35–38 |
| 姜建湖, 张德明, 竺俊全, 等, 2012. 光唇鱼(Acrossocheilus fasciatus)胚胎及仔、稚鱼的发育. 海洋与湖沼, 43(2): 280–287 DOI:10.11693/hyhz201202013013 |
| 姜景田, 2010. 马口鱼人工繁殖及养殖试验. 江西水产科技,(1): 27–28, 23 |
| 顾志敏, 朱俊杰, 贾永义, 等, 2008. 太湖翘嘴红鲌胚胎发育及胚后发育观察. 中国水产科学, 15(2): 204–214 |
| 徐伟, 耿龙武, 李池陶, 等, 2011. 大鳞鲃的人工繁殖、胚胎发育和耐盐碱测定. 水产学报, 35(2): 255–260 |
| 唐安华, 何学福, 1982. 云南光唇鱼Acrossocheilus yunanensis (Regan)的胚胎和胚后发育的初步观察. 西南师范学院学报,(1): 91–99 |
| 唐良华, 苏敏, 尤永隆, 等, 2006. 黑脊倒刺鲃仔、稚、幼鱼的生长发育研究. 淡水渔业, 36(1): 46–49 |
| 黄洪贵, 2009a. 中华倒刺鲃胚胎与仔鱼发育的观察. 江西农业大学学报, 31(6): 1087–1092 |
| 黄洪贵, 2009b. 倒刺鲃仔稚鱼发育的观察. 上海海洋大学学报, 18(5): 565–570 |
| 常剑波, 王剑伟, 曹文宣, 1995. 稀有鲫胚胎发育研究. 水生生物学报, 19(2): 97–103 |
| 董根, 杨建敏, 王卫军, 等, 2013. 短蛸(Octopus ocellatus)胚胎发育生物学零度和有效积温的研究. 海洋与湖沼, 44(2): 467–481 DOI:10.11693/hyhz201302032032 |
| 谢恩义, 阳清发, 何学福, 2002. 瓣结鱼的胚胎及幼鱼发育. 水产学报, 26(2): 115–121 |
| Bobe J, Labbé C, 2010. Egg and sperm quality in fish. General and Comparative Endocrinology, 165(3): 535–548 DOI:10.1016/j.ygcen.2009.02.011 |
| DeMartini E E, 1991. Annual variations in fecundity, egg size, and the gonadal and somatic conditions of queenfish Seriphus politus (Sciaenidae). Fishery Bulletin, 89(1): 9–18 |
| He Y F, Wu X B, Zhu Y J, et al, 2014. Effect of rearing temperature on growth and thermal tolerance of Schizothorax (Racoma) kozlovi larvae and juveniles. Journal of Thermal Biology, 46: 24–30 DOI:10.1016/j.jtherbio.2014.09.009 |
| Houman S, Hossein V G, Oryan S, et al, 2002. Biological characteristics of growth and reproduction in Barbus capito in south coasts of the Caspian Sea-Gilan Province. Journal of Marine Sciences and Technology, 1(4): 85–98 |
| Hsiao S, Greeley M S Jr, Wallace R A, 1994. Reproductive cycling in female Fundulus heteroclitus. The Biological Bulletin, 186(3): 271–284 DOI:10.2307/1542273 |
| Johansson F, 2006. Body shape differentiation among mitochondrial-DNA lineages of Zacco platypus and Opsariichthys bidens (Cyprinidae) from the Changjiang and Xijiang river drainage areas in southern China. Acta Zoologica Sinica, 52(5): 948–953 |
| Knutsen G M, Tilseth S, 1985. Growth, development, and feeding success of Atlantic cod larvae Gadus morhua related to egg size. Transactions of the American Fisheries Society, 114(4): 507–511 DOI:10.1577/1548-8659(1985)114<507:GDAFSO>2.0.CO;2 |
| Perdices A, Sayanda D, Coelho M M, 2005. Mitochondrial diversity of Opsariichthys bidens (Teleostei, Cyprinidae) in three Chinese drainages. Molecular Phylogenetics and Evolution, 37(3): 920–927 DOI:10.1016/j.ympev.2005.04.020 |
| Tyler C R, Nagler J J, Pottinger T G, et al, 1994. Effects of unilateral ovariectomy on recruitment and growth of follicles in the rainbow trout, Oncorhynchus mykiss. Fish Physiology and Biochemistry, 13(4): 309–316 DOI:10.1007/BF00003435 |