 2017, Vol. 48
2017, Vol. 48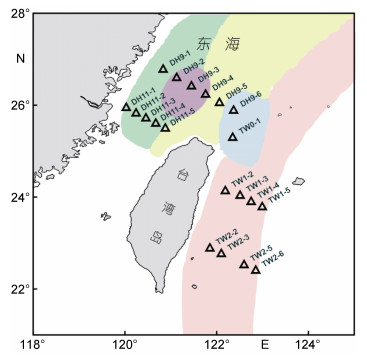


2014年春季黑潮与邻近海域甲藻组成及其对水团的指示作用

http://dx.doi.org/10.11693/hyhz20170400098
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 岑竞仪, 欧林坚, 李思, 张华, 王建艳, 吕颂辉. 2017.
- CEN Jing-Yi, OU Lin-Jian, LI Si, ZHANG Hua, WANG Jian-Yan, LÜ Song-Hui. 2017.
- 2014年春季黑潮与邻近海域甲藻组成及其对水团的指示作用
- THE SPECIES COMPOSITION OF DINOFLAGELLATE AND ITS ROLE OF INDICATION TO KUROSHIO AND ADJACENT WATERS IN SPRING 2014
- 海洋与湖沼, 48(5): 1022-1029
- Oceanologia et Limnologia Sinica, 48(5): 1022-1029.
- http://dx.doi.org/10.11693/hyhz20170400098
-
文章历史
- 收稿日期:2017-04-20
- 收修改稿日期:2017-07-03
引用本文
岑竞仪, 欧林坚, 李思, 张华, 王建艳, 吕颂辉. 2017. 2014年春季黑潮与邻近海域甲藻组成及其对水团的指示作用[J]. 海洋与湖沼, 48(5): 1022-1029
CEN Jing-Yi, OU Lin-Jian, LI Si, ZHANG Hua, WANG Jian-Yan, LÜ Song-Hui. 2017. THE SPECIES COMPOSITION OF DINOFLAGELLATE AND ITS ROLE OF INDICATION TO KUROSHIO AND ADJACENT WATERS IN SPRING 2014[J]. Oceanologia et Limnologia Sinica, 48(5): 1022-1029.
2014年春季黑潮与邻近海域甲藻组成及其对水团的指示作用
1.
暨南大学生命科学技术学院 广州 510632;
2. 水体富营养化与赤潮防治广东普通高校重点实验室 广州 510632;
3. 北京自然博物馆 北京 100050
2. 水体富营养化与赤潮防治广东普通高校重点实验室 广州 510632;
3. 北京自然博物馆 北京 100050
摘要:我国东部海域受近岸流、台湾暖流和黑潮的影响,不同水团间环境因子差异较大,甲藻作为初级生产者主要受环境影响,研究分析不同水团浮游甲藻群落组成及分布情况具有重要的生态意义。2014年5-6月,本研究对台湾以东黑潮主流径及东海200m等深线内陆架区进行采样调查,分析测站浮游甲藻的群落结构组成。研究结果表明,网采样品共鉴定出浮游甲藻17属81种,甲藻丰度范围为(0.45-1207.86)×103cell/m3。根据聚类分析结果,将海区浮游甲藻划分为3个群落类型,分别为受沿岸水团影响的群落Ⅰ、受台湾暖流水团影响的群落Ⅱ和黑潮水团影响的群落Ⅲ。不同水团间甲藻群落结构具有一定差异性,其中甲藻丰度变化趋势为沿岸水团>台湾暖流水团>黑潮水团,沿岸水团甲藻主要以广温广盐种为主,台湾暖流水团甲藻以广温高盐种为主,黑潮水团甲藻以高温高盐种为主。
关键词:甲藻群落 黑潮 指示种
THE SPECIES COMPOSITION OF DINOFLAGELLATE AND ITS ROLE OF INDICATION TO KUROSHIO AND ADJACENT WATERS IN SPRING 2014
1.
College of Life Science and Technology, Jinan University, Guangzhou 510632, China;
2. Key Laboratory of Aquatic Eutrophication and Control of Harmful Algae Blooms of Guangdong Higher Education Institutes, Guangzhou 510632, China;
3. Department of Science Research, Beijing Museum of Natural History, Beijing 100050, China
2. Key Laboratory of Aquatic Eutrophication and Control of Harmful Algae Blooms of Guangdong Higher Education Institutes, Guangzhou 510632, China;
3. Department of Science Research, Beijing Museum of Natural History, Beijing 100050, China
Abstract:
The East China Sea is affected by coastal current, Taiwan Warm Current and Kuroshio, and the environmental factors are different among its water masses. As a primary producer, dinoflagellates are sensitive to variation in the environment. Understanding the species composition of dinoflagellate is ecologically important. We conducted a survey in the main body of Kuroshio off eastern Taiwan Island on East China Sea shelf within 200 m in depth from May to June 2014. The results show that a total of 17 genera and 81 species of dinoflagellates were identified from the netz-samplers, and the abundance was from 0.45×103 to 1207.86×103cells/m3. In the cluster diagram, the phytoplankton dinoflagellates could be divided to three communities, including Group Ⅰ that is coastal current affected, Group Ⅱ in the Taiwan Warm Current (TWCW), and Group Ⅲ in the Kuroshio. The dinoflagellates grouping reveals that the coastal water mass ranked the first place in dinoflagellate diversity, followed by TWCW and Kuroshio in turn. In addition, the species composition of dinoflagellate was different from each other in ecological type in these water masses.
Key words:
dinoflagellate community
Kuroshio
indicator species
黑潮(Kuroshio)是大洋环流系中的一支西边界强流, 北半球副热带环流系的重要组成部分, 是强度仅次于墨西哥暖流的世界第二大洋流, 它以高温、高盐、流幅窄、流速强、流量大而著称, 发源于北赤道, 经菲律宾, 紧贴台湾东部进入东海, 后经琉球群岛, 沿日本列岛南部流去(张然等, 2016; 张灿影等, 2017)。黑潮在流经吕宋海峡时以分支或流套的形式入侵南中国海, 并部分流经台湾海峡, 而后在台湾东北部分出一个分支形成台湾暧流(宋军等, 2016)。黑潮运载着巨大的水量、热量和能量, 流向中纬度地带, 形成独特的高温、高盐特征, 影响着东海海域生物区系分布, 了解黑潮流系及其影响海域的浮游生物群落结构, 对于认识中国沿海生态的健康状态具有积极作用。
甲藻是海洋浮游植物中很重要的一个类群, 它们具有微弱的活动能力或没有活动能力而随波逐流, 其对栖息生境中各种环境因子, 特别是水动力因子的依赖性强, 不同生境中甲藻种类组成不尽相同, 因此研究甲藻的种类组成对大洋海流和水团分布具有一定的指示作用(顾新根等, 1995; 刘东艳等, 2000)。黑潮流系中浮游植物的群落结构, 具有因地而异、垂直分布不均匀的特点(伍伯瑜, 1986)。针对我国黑潮海域浮游植物研究, 规模较大的国际合作调查研究项目有1965—1977年间“黑潮及邻近海域的合作研究”、1986—1992年间实施的中日黑潮合作调查研究项目和1995—1998年间开展的中日副热带环流合作调查研究项目, 并取得了丰硕的成果, 出版了大量的资料报告(张灿影等, 2017)。随后我国学者也对东海海域、台湾海峡、吕宋海峡等黑潮影响海域的浮游植物进行了研究, 而对于多种水团都涵括的大面调查还较少(林更铭等, 2007; 谭激扬等, 2013; 周茜茜等, 2015)。本研究基于2014年春季(5—6月)对台湾以东黑潮主流径以及东海陆架海区(主要为中国东部受黑潮影响的海域, 属于东海近岸海域、台湾西北和东北海域, 涵括黑潮主流径、台湾暖流和东海沿岸流三种水团)开展的调查, 研究了该区域甲藻种类组成, 初步分析了其对不同水团的指示作用, 可为阐明黑潮对东中国海生态环境影响提供依据。
1 材料与方法 1.1 采样测站2014年5—6月, 搭乘“科学一号”对台湾以东黑潮主流径及东海200m等深线内的陆架区(120.03°—122.99°E, 22.40°—26.77°N)进行采样研究, 共设立四个断面20个测站(图 1)。
浮游甲藻样品采用浅Ⅲ小型浮游生物网(网口直径37cm, 网口面积0.1m2, 网身长140cm, 网目孔径76μm)由底层到表层进行垂直拖网采样。当水深超过200m时, 从200m处向上进行拖网。采集到的样品在现场用5%的缓冲福尔马林固定保存。在实验室浓缩后进行种类鉴定并在显微镜(Olympus BX61, 日本)下用浮游植物计数框进行计数。样品的采集、处理以及数据处理均按照《海洋调查规范-第六部分海洋生物调查》(GB/T 12763.6-2007) 执行。
1.3 数据处理群聚结构划分采用等级聚类分析(Cluster)和非线性多维标度排序(non-metric multidimensional scaling, NMDS)实现, 以各调查测站每种浮游甲藻的丰度为指标, 测站为样本, 使用丰度数据构建Bray-Curtis相似矩阵, 建立距离矩阵, 采用类平均法进行聚类, 作树状图, 并进行了NMDS分析验证聚类结果的可信程度。用相似性检验(analysis of similarity, ANOSIM)非参数多元分析方法检验两个群落之间是否具有显著性差异。用胁强系数(Stress)来检验NMDS分析结果的优劣, 当Stress<0.2时, 可认为NMDS的二维点图有一定的解释意义, Stress<0.1时, 认为是一个好的排序, Stress<0.05时, 认为NMDS图具有很好的代表性(Clarke et al, 2001)。甲藻群落结构分析采用Primer V.6软件运算, 调查海区测站平面分布图采用Golden Software Surfer 11.0软件作图。
细胞丰度采用浓缩计数法计算公式:
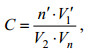
其中C为单位体积海水中标本总量; n'为取样计数个数; V1'为水样浓缩后体积; V2'为原采水量(滤水量以采样网口面积乘垂直拖网距离估算); Vn'表示取样计数的体积。
优势度采用McNaughton指数(Y)计算公式:
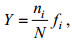
其中N表示各采样点所有物种个体总数; ni代表第i种的个体总数; fi表示该物种在各个采样点出现的频率, 当Y>0.02时, 该物种为群落中的优势种。
多样性指数H'(Shannon-Wiener index)计算公式(Shannon et al, 1949):
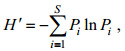
其中Pi表示第i种的个数与该样品总个数之比值; S为样品种类数。
均匀度指数J计算公式(Pielou, 1969)为:
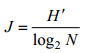
对2014年春季调查海区各测站浮游甲藻种类和丰度进行聚类分析和非参数多维标度排序, 发现在10%的Bray-Curtis相似性水平上, 将调查海区各测站划分为东海近岸海域群落(Ⅰ)和外海海域群落。在20%相似性水平上, 外海海域群落可进一步分为受台湾暖流影响海域群落(Ⅱ)和受黑潮主流域影响群落(Ⅲ), 同一群聚内部测站间的相似性水平较低。通过ANOSIM分析方法进行检验, 可知群落Ⅰ与群落Ⅱ差异性较明显(R=0.716, P=0.002), 群落Ⅲ与群落Ⅰ和Ⅱ差异性极明显(R=0.977, P=0.001; R=0.843, P=0.001)。经NMDS分析排序的胁强系数为0.07(<0.1), 说明NMDS图是一个好的排序(图 2)。由NMDS图可以看出群落Ⅰ与群落Ⅲ距离最远, 群落间种类组成差异最大。而群落Ⅰ与Ⅱ距离较近, 群落内组成具有一定的相似之处。
 |
| 图 2 调查海域春季浮游甲藻群落Bray-Curtis聚类图(a)和非线性多维标度排序分析(b) Fig. 2 Bray-Curtis cluster(a) and non-metric multidimensional scaling analysis(b) of dinoflagellate community structure in the study area in spring |
通过对调查海域浮游甲藻种类及丰度聚类分析和非参数多维标度排序可把浮游甲藻群落分为三类, 分别为:东海近岸海域群落(Ⅰ)、台湾暖流海域群落(Ⅱ)和黑潮主流域群落(Ⅲ), 与调查海域水团自然分布情况(图 1)相拟。
2.2 不同海域的浮游甲藻种类组成调查期间共鉴定出浮游甲藻17属81种(含5种未定种), 其中新角藻属(Neoceratium)31种, 占总种类数的37.8%, 原多甲藻(Protoperidinium)属18种, 占总种类的22%。在调查期间整个海区浮游甲藻优势种有:矮胖新角藻(N. humile)、三角新角藻(N. tripos)、具尾鳍藻(Dinophysis caudata)、夜光藻(Noctiluca scintillans)和东海原甲藻(Prorocentrum donghaiense)。
群落Ⅰ近岸海域:共鉴定出浮游甲藻13属42种(含4种未定种), 其中新角藻属和原多甲藻属的种类数最多, 分别为17种(占总种类数40.48%)和12种(占总种类数28.57%)。其中分布最广的种类有:矮胖新角藻、梭状新角藻(N. fusus)、三角新角藻、叉状新角藻(N. furca)、具尾鳍藻、夜光藻, 生态类型多为广温广盐种。
群落Ⅱ台湾暖流流经海域:共鉴定出浮游甲藻17属49种(含4种未定种), 其中新角藻属和原多甲藻属种类数最多, 分别为22种(占总种类数44.90%)和13种(占总种类数26.53%)。
群落Ⅲ黑潮流经海域:共鉴定出浮游甲藻12属50种(含1种未定种), 其中新角藻属的种类数最多, 共25种(占总种类数50%)。
2.3 不同海域的浮游甲藻优势种调查海域从近岸到外海, 水系复杂, 受到黑潮、台湾暖流、近岸水等多重水团影响, 浮游甲藻种类丰富, 生态类型多样, 不同水团甲藻种类组成和优势种存在一定差异, 按照浮游甲藻群落聚类分析可分成3个类群(图 2)。群落Ⅰ近岸海域的优势种有矮胖新角藻、三角新角藻、具尾鳍藻、夜光藻、东海原甲藻。除东海原甲藻只在最靠近岸测站(DH9-1, DH1-1) 出现, 细胞丰度占浮游甲藻总细胞丰度的48.78%, 其他优势种在群落Ⅰ所有测站都存在; 除具尾鳍藻优势度(Y=0.04) 较低外, 其他优势种Y都大于0.1。群落Ⅱ台湾暖流流经海域的优势种与群落Ⅰ和Ⅲ差异较大, 主要特征是该区域优势种以外海种类为主, 优势种类数较多, 优势度均不高。群落Ⅲ黑潮流经海域的主要优势种为波状新角藻(N. trichoceros), 在所有群落Ⅲ测站都出现, 细胞丰度占总细胞丰度的22%, 优势度为Y=0.22, 其他优势种Y都小于0.1(表 1)。
表 1 不同群落甲藻优势种优势度(Y)比较 Tab. 1 Dominant dinoflagellates of different communities in the study area in spring
| 种名 | 群落Ⅰ | 群落Ⅱ | 群落Ⅲ | |
| 矮胖新角藻 | Neoceratium humile | 0.11 | - | - |
| 歧分新角藻 | N. carriense | - | - | 0.03 |
| 偏转新角藻 | N. deflexum | - | 0.03 | - |
| 大角新角藻 | N. macroceros | - | - | 0.05 |
| 马西里亚新角藻 | N. massiliense | - | 0.08 | 0.03 |
| 三角新角藻 | N. tripos | - | 0.09 | 0.22 |
| 兀鹰新角藻 | N. vultur | - | 0.03 | - |
| 梭状新角藻 | N. fusus | - | 0.05 | - |
| 波状新角藻 | N. trichoceros | 0.19 | 0.06 | - |
| 具尾鳍藻 | Dinophysis caudata | 0.04 | 0.06 | - |
| 轮状拟翼藻 | Diplopsalopsis orbicularis | - | 0.06 | - |
| 夜光藻 | Noctiluca scintillans | 0.10 | - | - |
| 东海原甲藻 | Prorocentrum donghaiense | 0.14 | - | - |
| 海洋原多甲藻 | Protoperidinium oceanicum | - | 0.05 | - |
| 斯氏扁甲藻 | Pyrophacus steinii | - | 0.08 | 0.08 |
| 注: “-”代表该甲藻在这个群落中不是优势种。 | ||||
如图 3所示, 调查海区网样浮游甲藻细胞丰度范围为(0.45—1207.86)×103cell/m3, 其中群落Ⅰ近岸海域细胞丰度变化范围为(23.26—1207.86)×103cell/m3, 群落Ⅱ台湾暖流流经海域变化范围为(0.67—15.19)× 103cell/m3; 群落Ⅲ黑潮流经海域变化范围为(0.45—1.94)×103cell/m3。调查海域网样浮游植物细胞丰度自外海到近海呈递增趋势。
 |
| 图 3 调查海域甲藻群落细胞密度分布 Fig. 3 The distribution of dinoflagellate abundance in the study area |
2014年春季, 调查海域的多样性指数(H')范围为1.28—2.89, 平均值为2.15, 总体变化趋势为近岸海域较低, 外海海域较高(图 4)。群落Ⅰ多样性指数范围为1.28—1.89, 平均值为1.60;群落Ⅱ多样性指数范围为2.44—2.89, 平均值为2.66;群落Ⅲ多样性指数范围为1.84—2.66, 平均值为2.30。群落Ⅰ多样性指数平均值比群落Ⅱ和Ⅲ都低, 反映了群落Ⅰ群类结构比群落Ⅱ和Ⅲ脆弱, 群落内优势种细胞丰度较高, 占总细胞丰度的百分比高(93.38%), 而近岸群落Ⅰ均匀度(J)均值为0.60, 比群落Ⅱ(J=0.9) 和群落Ⅲ(J=0.91) 都低, 也从侧面反映出群落Ⅰ优势种对群落结构影响较大。
 |
| 图 4 调查海域甲藻群落多样性指数(H')的水平分布 Fig. 4 Horizontal distribution of Shannon-Wiener diversity index of dinoflagellate communities in the study area |
甲藻对环境的适应性依照种类而有所不同, 生活在不同水系甲藻的组成和分布不一样, 反之甲藻的群落结构也可以反映环境情况(陆斗定, 1991; 李瑞香等, 1992)。本研究以调查海域甲藻种类及丰度进行聚类分析、非线性多维标度排序分析和群落相似性分析, 可以把调查海域浮游甲藻分为3种群落, 聚类分析结果与本次调查期间海区水团自然分布情况相一致。通过ANOSIM对3个群落进行相似性分析, 从结果可以反映出群落Ⅰ与群落Ⅲ群落组成差异性极大(R=0.977), 而群落Ⅱ与Ⅲ群落结构较为相近。其中群落Ⅰ(DH9-1、DH9-2、DH9-3、DH9-4和DH11-1、DH11-2、DH11-3、DH11-4测站), 主要位于东海近岸海域, 海区同时受陆源淡水输入以及外海台湾暖流高温高盐水的影响, 温度和盐度变化范围较大, 平均盐度低于33(卢汐等, 2016), 甲藻群落组成以近岸低盐广温种和广温广盐种为主; 群落Ⅱ(TW0、DH9-4、DH9-5、DH9-6和DH11-5测站)主要位于台湾以北海域, 受台湾暖流影响, 水团表层温度较高, 盐度高于33, 其中TW0表层(0—30m)温度变化范围为21.22—23.65℃, 盐度变化范围为34.33—34.52(卢汐等, 2016), 甲藻群落组成主要为广温广盐和高温高盐种类; 群落Ⅲ(TW1和TW2断面)主要位于台湾以东海域, 为黑潮主流径, 海域以高盐(>34) 为特征, 表层(0—30m)盐度为34.59±0.14, 次表层(30—50m)平均盐度为34.73±0.09(卢汐等, 2016), 甲藻群落组成主要以高温高盐种类为主。
3.2 不同水团甲藻群落结构分析调查海域水团复杂, 可分为三大水团:沿岸水团、台湾暖流水团和黑潮水团, 不同水团浮游植物群落的差异主要受水团环境因子的影响。东海黑潮区浮游植物种类组成、丰度变化与温度、盐度变化密切相关, 黑潮水团甲藻种类丰富, 以高温高盐为主, 其中狭高温高盐种类可以作为其指示种; 台湾暖流水团以广温广盐种和高温高盐种为主, 其中近岸暧水种可以作为其指示种; 沿岸水团以低盐近岸类群为主, 其中夜光藻为代表种(陆斗定, 1991; 刘亚林等, 2016)。本研究结果显示, 沿岸水团(群落Ⅰ)以低盐近岸种和广温种为主(占总丰度93.38%), 贡献最大的分别为东海原甲藻(49.83%)、三角新角藻、矮胖新角藻和夜光藻, 其中东海原甲藻只出现在离岸最近的DH9-1和DH11-1测站, 夜光藻在沿岸水团所有的测站都出现, 丰度也比较高, 可作为沿岸水团的指示种; 台湾暖流水团(群落Ⅱ)以暧水种为主(占总丰度68.8%), 此外还出现较多的广温广盐种, 常见种有偏转新角藻(N. deflexum)、柔软新角藻(N. molle)、马西里亚新角藻(N. massiliense)、波状新角藻、兀鹰新角藻(N. vultur)、梭状新角藻、三角新角藻、具尾鳍藻、轮状拟翼藻(Diplopsalopsis orbicularis)、扁平原多甲藻(Protoperidinium depressum)、叉分原多甲藻(P. divergens)、海洋原多甲藻(P. oceanicum)、新月梨甲藻(P. lunula)、斯氏扁甲藻(Pyrophacus steinii); 黑潮水团(群落Ⅲ), 以高温高盐种为主(占总丰度83.04%), 其中主要优势种为波状新角藻(占总丰度22%), 常见种有蜡台新角藻(N. candelabrum)、大角新角藻(N. macroceros)、波状新角藻、斯氏扁甲藻。不同水团甲藻生态类群丰度和组成的差异较大, 反映了不同生态类群对不同环境的适应。
陆斗定等(1990)对1986年春季黑潮区与邻近海域甲藻丰度进行研究, 发现台湾西北部海域甲藻丰度(>1.2×103cell/m3)比台湾以东黑潮主干道的甲藻丰度(<0.8×103cell/m3)高, 而本次调查中相对应海域的丰度分别为大于5×103cell/m3和小于2×103cell/m3, 比1986年调查期间的细胞丰度高。本研究调查期间海域不同水团甲藻丰度变化趋势表现为沿岸水团>台湾暖流水团>黑潮水团。黑潮水团甲藻组成种类多, 但丰度较低, 种类以高温高盐种为主。
3.3 黑潮水团指示种生物的生存与海洋环境条件密切相关, 海洋环境的变化直接影响到生物生态类型的分布及其数量的改变(苗育田等, 1998), 黑潮水团具有高温高盐等水文特征, 在其中分布的一些生物对黑潮水团具有指示作用。在对中国东海黑潮区及相关海域研究中, 部分学者以一些狭高温高盐生态类型或一些只分布于黑潮水团的浮游植物作为黑潮水团的指示种(杨清良等, 1999; Chen et al, 2003; 刘海娇等, 2015)。甲藻具有微弱的运动能力和特殊的营养类型, 在大洋中它们往往是浮游植物群落中的主要类群, 对大洋海流和水团分布有良好的指示作用, 部分种类能作为大洋水团和海流的指示种(Wood, 1963; Abé et al, 1981; 刘东艳等, 2000)。在我国最早(1965—1977年)对黑潮海域的研究中, 提出以一些狭高温高盐甲藻种类作为东海陆架的黑潮指示种(郭玉洁等, 1982; 毛兴华等, 1984); 随后在1986—1988年通过中日合作黑潮调查研究, 国内学者陆斗定(1991)通过对四个航次浮游甲藻分析, 提出以狭高温高盐种和一些热带罕见甲藻种类(共25种)作为黑潮水系指示种; 李瑞香等(1992)在对1989年黑潮区域甲藻的研究中, 以只局限于黑潮区域和对马暖流的狭高温高盐种双管藻(Amphisolenia spp.)、鸟尾藻(Ornithocercus spp.)、梨甲藻(Pyrocystis spp.)和新角藻等作为黑潮指示种; 近几年在针对东海海域的相关研究中, 证实了鸟尾藻属、梨甲藻属、角甲藻属(Ceratocorys)部分狭高温高盐种主要分布于黑潮及受其影响的海域, 能对黑潮水系起到指示作用(王丹等, 2006; Lin et al, 2012; 谭激扬等, 2013; 周茜茜等, 2015)。
通过历史数据统计, 在我国黑潮及其影响海域研究中, 统计出黑潮指示种41种, 隶属于6属(见表 2)。与历史数据比较, 本调查期间共鉴定出黑潮指示种19种, 其中黑潮主干流出现14种, 台湾暖流水团出现12种, 沿岸水团出现6种, 其中有5种出现在DH9-3测站, 而此测站处于60m等深线附近地形变化处, 温度、盐度垂向分布均一, 为陆架垂直混合剧烈的区域, 呈现出台湾海峡水流的温盐特征(卢汐等, 2016)。刘东艳等(2000)在对琉球群岛及其邻近海域甲藻研究中把具尾鳍藻作为琉球群岛岛链以西的黑潮指示种, 但此种在中国沿岸未受黑潮影响的海域也有分布且属于广布种, 在本次调查中具尾鳍藻在沿岸水团所有测站和台湾暖流水团都有分布, 而在黑潮水团海域并没有分布, 它不是黑潮水团特有的种类。除具尾鳍藻外, 在本次调查中只局限于黑潮水团的甲藻种类有15种, 其中三叉双管藻(Amphisolenia thrinax)、尖新角藻(N. belone)、板状新角藻(N. platycorne)、方鸟尾藻(O. quadratus)和钩梨甲藻(P. hamulus)5种被认为是黑潮指示种, 另外美丽新角藻(N. pulchellum)、楔形鳍藻(D. cuneus)和优美梨甲藻(P. elegans)3种狭高温高盐种也只分布于此海域, 以上种类可作为黑潮主干径指示种; 分布于黑潮水团和台湾暖流水团的种类有10种, 其中二齿双管藻(A. bidentata)、二裂新角藻(N. biceps)和柔软新角藻等广温高盐种和扭状新角藻(N. contortum)、偏转新角藻、网纹新角藻(N. hexacanthum)、蛙指新角藻(N. ranipes)、新月梨甲藻、长刺角甲藻(C. horrida)等狭高温高盐种可以作为黑潮水团及台湾暖流水团指示种。
表 2 黑潮指示种 Tab. 2 The indicator species to the Kuroshio
| 种名 | 拉丁文 |
| 四齿双管藻 | Amphisolenia schauinslandi |
| 三叉双管藻* | A. thrinax |
| 尖新角藻* | Neoceratium belone |
| 毕氏新角藻 | N. bigelowii |
| 脑形新角藻 | N. cephalatum |
| 扭状新角藻* | N. contortum |
| 偏转新角藻* | N. deflexum |
| 叉状新角藻矮胖变种 | N. furca var. eugrammum |
| 曲肘新角藻 | N. geniculatum |
| 瘤状新角藻* | N. gibberum |
| 圆头新角藻* | N. gravidum |
| 网纹新角藻* | N. hexacanthum |
| 矮胖新角藻 | N. humile |
| 剑峰角藻* | Ceratium incisum |
| 科氏新角藻* | N. kofoidii |
| 马西里亚新角藻* | N. massiliense |
| 板状新角藻 | N. platycorne |
| 蛙指新角藻* | N. ranipes |
| 锥形新角藻 | N. schroeteri |
| 波状新角藻 | N. trichoceros |
| 三角新角藻印度变种* | N. tripos var. indicum |
| 兀鹰新角藻* | N. vultur |
| 长刺角甲藻* | Ceratocorys horrida |
| 具尾鳍藻* | Dinophysis caudata |
| 勇士鳍藻 | D. mitra |
| 大鸟尾藻 | Ornithocercus magnificus |
| 方鸟尾藻* | O. quadratus |
| 美丽鸟尾藻 | O. splendidus |
| 四叶鸟尾藻 | O. steinii |
| 瑟氏鸟尾藻* | O. thurnii |
| 纺锤梨甲藻双锥变型 | Pyrocystis fusiformis f.bicornia |
| 尖梨甲藻 | P. acuta |
| 钩梨甲藻* | P. hamulus |
| 钩梨甲藻异肢变种 | P. hamulus var.inaeaqualis |
| 钩梨甲藻半圆变种 | P. hamulus var. semicircuralis |
| 新月梨甲藻* | P. lunula |
| 粗梨甲藻 | P. robusta |
| 矛形梨甲藻 | P. lanceolata |
| 光甲多甲藻 | Peridium pallidum |
| 注: *表示本调查期间鉴定出的种类; 相关记录参考已报道文献(毛兴华等, 1984; 林金美. 1994; 陆斗定等, 1990, 1991; 李瑞香等, 1992; 刘东艳等, 2000; 刘亚林等, 2016)。 | |
(1) 调查期间海域不同水团甲藻丰度变化趋势表现为沿岸水团>台湾暖流水团>黑潮水团。黑潮水团甲藻呈种类多, 丰度较低特点。
(2) 三叉双管藻、尖角藻、板状新角藻、方鸟尾藻、钩梨甲藻、美丽新角藻、楔形鳍藻、和优美梨甲藻等狭高温高盐种可作为黑潮主干径指示种; 二齿双管藻、二裂角藻、柔软新角藻、扭状新角藻、偏转新角藻、网纹新角藻、蛙指新角藻、新月梨甲藻和长刺角甲藻等狭高温高盐种可以作为黑潮水团及台湾暖流水团指示种。
致谢 本研究中的海洋调查和样品采集工作得到了厦门大学环境与生态学院各位老师、同学及考察船全体工作人员的支持, 谨此致谢。
参考文献
| 王丹, 孙军, 安佰正, 等, 2008. 2006年秋季东海陆架浮游植物群集. 应用生态学报, 19(11): 2435–2442 |
| 毛兴华, 李瑞香, 1984. 东海北部陆架区浮游甲藻的分布及其生态特征. 海洋学报, 6(5): 672–677 |
| 卢汐, 宋金明, 袁华茂, 等, 2016. 黑潮主流径海域海水中的无机碳及其对东海陆架区的影响. 海洋与湖沼, 47(1): 16–28 |
| 伍伯瑜, 1986. 黑潮流系-Ⅲ黑潮流系的浮游生物及其他. 海洋通报, 5(3): 74–78 |
| 林金美, 1994. 东海浮游甲藻类的分布. 海洋学报, 16(2): 110–115 |
| 刘东艳, 孙军, 钱树本, 2000. 琉球群岛及其邻近海域的浮游甲藻-1997年夏季的种类组成和丰度分布. 见: 中国海洋学文集-西北太平洋副热带环流研究(二). 北京: 中国海洋学会 |
| 刘亚林, 黄秀清, 蒋晓山, 等, 2016. 2014年冬季浙江中部海域网采浮游植物群集. 海洋科学, 40(4): 46–53 |
| 刘海娇, 傅文诚, 孙军, 2015. 2009-2011年东海陆架海域网采浮游植物群落的季节变化. 海洋学报, 37(10): 106–122 DOI:10.3969/j.issn.0253-4193.2015.10.011 |
| 李瑞香, 俞建銮, 1992. 东海黑潮区甲藻的分布及其对水系的指示作用. 见: 国家海洋局科技司. 黑潮调查研究论文选(四). 北京: 海洋出版社, 182-190 |
| 杨清良, 林更铭, 林金美, 1999. 1996年春季副热带环流区浮游植物生态的初步研究. 海洋科学, 21(5): 120–128 |
| 宋军, 郭俊如, 鲍献文, 等, 2016. 东海黑潮与陆架海之间的水交换研究. 海洋通报, 35(2): 178–186 DOI:10.11840/j.issn.1001-6392.2016.02.008 |
| 张然, 徐海明, 张百超, 2016. 东中国海黑潮海洋锋的季节变化特征及其成因. 气象科学, 36(2): 203–211 |
| 张灿影, 冯志纲, 张晓琨, 等, 2017. 基于文献计量的黑潮研究进展分析. 世界科技研究与发展, DOI:10.16507/j.issn.1006-6055.2017.03.002 |
| 陆斗定, 1991. 东海黑潮指示性甲藻的分布特征. 见: 国家海洋局科技司. 黑潮调查研究论文选(三). 北京: 海洋出版社, 287-296 |
| 陆斗定, 徐芝敏, 蒋加伦, 1990. 1986年日本南部浮游甲藻群落组成特点. 见: 国家海洋局科技司. 黑潮调查研究论文选(二). 北京: 海洋出版社, 278-283 |
| 陆斗定, 蒋加伦, 徐芝敏, 1990. 1986年春季东海黑潮区浮游甲藻种类组成及其分布特征的初步分析. 见: 国家海洋局科技司. 黑潮调查研究论文选(一). 北京: 海洋出版社, 229-238 |
| 苗育田, 于洪华, 何德华, 等, 1998. 正常天气系统下的东海水系与指示生物对应关系的分析. 海洋通报, 17(2): 13–21 |
| 林更铭, 杨清良, 2007. 台湾海峡小型浮游植物的物种多样性和分布特征. 生物多样性, 15(1): 31–45 |
| 周茜茜, 陈长平, 梁君荣, 等, 2015. 中国东部陆架边缘海网采浮游植物种类组成和季节变化. 生物多样性, 23(1): 23–32 |
| 顾新根, 袁骐, 沈焕庭, 等, 1995. 长江口最大浑浊带浮游植物的生态研究. 中国水产科学, 2(1): 16–27 |
| 郭玉洁, 杨则禹, 1982. 1976年夏季东海陆架区浮游植物生态的研究. 海洋科学集刊, 19: 11–32 |
| 谭激扬, 黄良民, 谭烨辉, 等, 2013. 水团对吕宋海峡浮游植物群落结构的影响. 海洋学报, 35(6): 178–189 |
| Abé T H, Saito M, 1981. Studies on the Family Peridinidae.An Unfinished Monograph of the Armoured Dinoflagellata. Seto Marine Biological Laboratory, Special Publication Series, 6: 421 |
| Chen Y L L, Chen H Y, Lin Y H, 2003. Distribution and downward flux of Trichodesmium in the South China Sea as influenced by the transport from the Kuroshio Current. Marine Ecology ProgressSeries, 259: 47–57 DOI:10.3354/meps259047 |
| Clarke K R, Warwick R M, 2001. A further biodiversity index applicable to species lists:Variation in taxonomic distinctness. Marine Ecology Progress Series, 216: 265–278 DOI:10.3354/meps216265 |
| Lin G M, Yang Q L, Lin W H, et al, 2012. Distribution characteristic and variation trend of planktonic dinoflagellate in the Taiwan Strait from 2006 to 2007. Marine Science Bulletin, 14(1): 68–79 |
| Pielou E C, 1969. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, 1-286 |
| Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1-125 |
| Wood E J F, 1963. Dinoflagellates in the Australian region. Ⅱ. Recent collections.Division of Fisheries and Oceanography, Technical Paper No. 14, Melbourne, Australia:Commonwealth Scientific and Industrial Research Organization, 14:1-55 |