 2018, Vol. 49
2018, Vol. 49中国海洋湖沼学会主办。
文章信息
- 徐鹏, 黄菊, 蔡厚才, 陈万东, 章守宇. 2018.
- XU Peng, HUANG Ju, CAI Hou-Cai, CHEN Wan-Dong, ZHANG Shou-Yu. 2018.
- 南麂列岛养殖功能海域秋季潮致混合特征及其对营养盐浓度的影响
- The characteristics of tide-induced mixing and its iMPACT ON nutrient concentration in BREADING GROUNDS IN Autumn AROUND Nanji Archipelago, EAST CHINA SEA
- 海洋与湖沼, 49(1): 17-23
- Oceanologia et Limnologia Sinica, 49(1): 17-23.
- http://dx.doi.org/10.11693/hyhz20170500111
-
文章历史
- 收稿日期:2017-05-04
- 收修改稿日期:2017-06-14




2. 南麂列岛国家海洋自然保护区研究所 温州 325400
2. Institute of Nanji Archipelago National Nature Reserve, Wenzhou 325400, China
随着经济社会的快速发展, 我国海洋蓝色经济在过去几十年中得到了长足进步。然而长期以来, 受生态环境恶化与过度捕捞的影响, 我国近海渔业资源已严重衰退(王大海, 2014)。为了应对渔业资源的衰退, 海水养殖在我国迅猛发展并已成为我国海产品的主要来源(林丽华, 2005)。海水养殖资源在空间分布上具有地域性, 这就在一定程度上导致了养殖海域附近生态环境问题。为了实现海水养殖产业的可持续发展, 需厘清养殖海域内物质循环与能量流动规律, 其中水动力过程对营养盐的输运是一个重要方面(魏皓等, 2010)。
海底沉积物是水体中营养盐的一个重要来源(刘素美等, 2000; 潘建明等, 2002), 湍流混合可将位于水体底层的营养盐卷挟至水体上层, 从而支撑浮游植物的生长(Sharples et al, 2001; 魏皓等, 2002; Jin et al, 2013)。在近岸浅海, 潮过程是最重要的动力因子之一(Munk et al, 1998), 潮致混合对营养盐的垂向输运具有重要贡献(Souza et al, 2001; Castro et al, 2011)。通常情况下, 正压潮致混合在底边界层内比较显著, 且随离底距离的增加而强度减弱(如徐鹏等, 2013)。当斜压过程介入时, 潮致混合表现出与正压情况下不同的特点(如Wang et al, 2014)。但上述这些认识多是基于在天然海域的观测, 在养殖功能海域, 特别是在有大型人工养殖设施存在的情况下, 潮致混合如何响应, 对营养盐的垂向输运又有何影响, 目前仍鲜有报道。
南麂列岛位于浙江省东南部, 属强潮陆架海区。该列岛地处亚热带, 海洋自然环境条件优越, 有“贝藻王国”的美誉(孙建璋等, 2009)。以保护海洋生物多样性为目标, 以海洋贝藻类及其生态环境为主要保护对象, 南麂列岛经国务院批准成为我国首批五个海洋类自然保护区中的一个。同时南麂列岛还是浙江省重要的水产养殖基地, 是该省首批四大深水网箱规模化养殖示范基地之一(蔡厚才等, 2004)。大黄鱼是南麂列岛深水网箱养殖的重要鱼种, 且养殖规模在近几年迅速扩大(庄定根, 2014)。本文将通过分析实测数据, 阐明在大型人工养殖设施影响下的潮致混合特征及其对营养盐浓度的影响, 从而为相似海域可持续发展的海水养殖提供参考和依据。
1 现场观测2017年11月16日17时, 研究团队于南麂岛东北大黄鱼深水网箱养殖区进行现场数据采集(图 1), 整个观测过程持续25小时。观测采用海床基观测与船基观测同步进行的方式, 其中海床基观测平台布放RDI Workhorse 1200kHz声学多普勒流速剖面仪(ADCP)一台, RBR温深仪(TD)一台, 船基观测则主要进行温盐剖面观测与水样采集。ADCP采用仰视工作方式, 并用beam坐标记录流速。设置其发射声波频率为2Hz, 数据输出频率为0.5Hz, 数据垂直分辨率为0.5m。TD设置为1Hz连续采样。船基温盐剖面观测利用SBE 19plus温盐深仪(CTD)完成, 采样频率为4Hz, 每小时观测一次。水样采集同样采用整点观测的方式, 对瞬时水深表、中、底三层分别进行采样。现场在样品中加入三氯甲烷后用保温箱带回实验室分析营养盐浓度。
 |
| 图 1 观测站位 Fig. 1 Map of the study site 注:左上角子图中的红框标示了南麂列岛的位置 |
观测站位紧靠养殖网箱, 该处平均水深约17.7m。养殖网箱呈圆柱形, 直径约19.1m, 高约7.5m (图 2)。受周围岛屿遮蔽作用影响, 观测期间风速不大, 无明显波浪。南麂海域潮波以半日潮为主(图 3), 观测期间最大潮差约5.4m。由于周围岛屿岸线的限制, 观测站位处潮流为往复流, 流向沿东南-西北方向(图片未展示)。观测站位处潮位未表现出明显的不对称结构, 但落潮流(流向约152°)却显著强于涨潮流(流向约332°)。观测期间最大落潮流速约为0.55m/s, 而最大涨潮流速约只有0.15m/s(图 3)。值得注意的是, 一个潮周期内两次转流时潮流等值线具有明显不同的类型, 一种为由涨至落时的竖直型, 即潮流自底至表近乎同落; 另一种为由落至涨时的倾斜型, 即底层潮流先落。这种转流时潮流不同的时空结构将对流速剪切与雷诺应力等产生重要影响。无论落潮流与涨潮流, 都在转流后不久达到其各自峰值, 分别大致对应高、低潮时。
 |
| 图 2 观测站位处深水养殖网箱照片 Fig. 2 The maricultureal cages in the study site |
 |
| 图 3 ADCP采集的流速剖面 Fig. 3 Profiles of currents recorded by ADCP 注:图中上端空白处黑线表示水位, 下端彩图表示流速, 其中正、负值分别表示西北、东南方向流速; 彩图中两条粗黑线分别标注了落潮阶段流速等值线为竖直和倾斜的两种状态; 五角星则标注了这两个时刻深水网箱底端所处的深度; 这种设定同样应用到本文其他图片中 |
雷诺应力是表征湍流强度一个重要参量(Wang et al, 2014)。利用声学流速剖面观测资料并根据“方差法”可以计算水体中雷诺应力。该方法最早由Lohrmann等(1990)提出。Stacey等(1999)与Lu等(1999)对该方法进行了发展, 并首先将其应用到陆架浅海。目前, 该方法已被广泛用于陆架浅海与河口海湾中雷诺应力的估计(Rippeth et al, 2002).
方差法将ADCP在beam坐标系下记录的原始流速bi (i=1, 2, 3, 4)进行雷诺分解, 即分解为平均流速与湍流脉动之和, bi =bi +bi′(i=1, 2, 3, 4)。利用bi与笛卡儿坐标系下3个正交流速分量u, v, w间的几何关系以及雷诺应力的定义, 可得
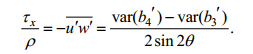 (1)
(1)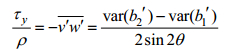 (2)
(2)其中, τx与τy分别为雷诺应力矢量在x与y方向的分量; ρ为海水密度; var(bi′)为bi′的方差; θ为ADCP 4个beam方向与仪器中心轴(即z方向)的夹角, 在本实验中为20°。在计算得到x与y方向的雷诺应力后, 再将其投影到潮流方向, 得到沿潮流方向的雷诺应力。
2.2 全自动间断化学分析仪测定营养盐浓度本文利用第2代CleverChem全自动间断化学分析仪进行营养盐浓度分析。该仪器实际上是将传统的比色法机械化, 自动化, 共包括自动取样器、化学反应单元、检测器和数据处理系统四个部分。它采用目前世界上最先进的第二代全自动间断化学分析技术, 用吸光度表示反应终点, 采用比色管直读方式, 样品与试剂在独立的比色管中反应和成色, 完全做到间断分析, 可以在无人看管的情况下一次测定64个样品的9个参数。反应过程由电脑控制, 自动化程度高, 操作简便, 减少了人为干扰, 监测结果精确可靠, 已被广泛用于生物(Kim et al, 2008; Van Verseveld et al, 2009)、环境(Santoro et al, 2008)、水处理(Van Ginkel et al, 2008)等领域。
3 结果与分析图 4展示了利用“方差法”算得的沿潮流方向雷诺应力的时空分布特征。需要说明的是, 这里展示的结果是将原始数据乘以了一个校正系数(1.3)。尽管本文所用的ADCP设置(2s/ensemble, 0.5s/ping, 0.5m/bin)被广泛地应用于近海雷诺应力观测(Rippeth et al, 2002; Williams et al, 2004), 但这种设置实际上是对数据时空分辨率与数据体量的一种兼顾。Rippeth等(2002)曾将此设置下的结果与同步观测得到的一个更高时空分比率(0.5s/ensemble, 0.5s/ping, 0.25m/bin)的结果作比较, 从而将校正系数确定为1.3。

受潮流不对称的影响, 雷诺应力呈现出明显的落潮占优特点。涨潮时雷诺应力量值较小, 接近于噪声水平, 而落潮时雷诺应力量值显著增大, 可达8×10-4m2/s2(图 4)。与转流时潮流具有两种不同的时空结构相对应, 雷诺应力在转流时的量值与垂向结构也显著不同。从量值上来讲, 高潮时(由涨转落)雷诺应力较低潮时(由落转涨)明显偏小, 前者最大值约是后者的1/2。从垂向结构来讲, 高潮时水体中部与近底层雷诺应力没有明显的位相差(观测期间第2个高潮时这种空间结构更清晰), 而低潮时雷诺应力在垂直方向上呈现出明显的位相差, 各水层雷诺应力峰值自底至表陆续出现。
与在天然海域观测到的正压潮致混合特征不同(如Rippeth et al, 2002), 观测站位处雷诺应力最大值并没有被限制在近底层, 而是出现在水体中层(图 4)。高潮近底层雷诺应力约为4×10-4m2/s2, 而水体中部的值可达6×10-4m2/s2(观测期间第2个高潮时)。低潮时近底层雷诺应力与高潮时相差不大, 但水体中部的值可高达8×10-4m2/s2。造成这种影响的可能原因有两个, 一为人工养殖网箱的影响, 二为斜压过程的影响。接下来本文将结合同步采集到的盐度、流速数据对其进行分析。
观测期间, 站位处盐度随潮位相在28.1—28.9范围内变化, 其中涨潮时盐度增大, 落潮时盐度降低(图 5a)。考虑到观测海域附近潮流为往复流, 盐度的这一变化说明该海域在11月份具有明显的水平盐度梯度, 这与更大空间范围的实测结果是相符的(Chen et al, 2006)。该盐度梯度方向由西北指向东南, 即与落潮流同向。由盐度剖面的时间变化可推断(图 5a), 高潮时观测站位处水平盐度梯度较弱, 而低潮时则显著增强。在垂向上, 低潮时西北方向的低盐水平流至观测站位, 使水体呈现强层化, 浮力频率平方介于O(10-3.5/s2)—O(10-3.0/s2); 高潮时高盐水重新控制观测站位, 水体层化转弱, 浮力频率的平方介于O(10-4.5/s2)—O(10-4.0/s2)(图 5b)。
 |
| 图 5 调查期间同步采集的盐度(a)、浮力频率平方(b)、流速剪切(c)、理查德森数(d)剖面 Fig. 5 Profiles of salinity (a), the square of buoyance frequency (b), currents shear (c), and Richardson number (d) simultaneously collected during the observation |
受潮流不对称影响, 涨潮时流速剪切较小, 而落潮时流速剪切较大(图 5c)。在近底层, 涨潮时流速剪切约为-0.02/s, 而落潮时显著增大为0.05/s。落潮阶段大部分时间内, 流速剪切峰值被限制在距底4m以内, 直至低潮转流时, 流速剪切峰值自底至表陆续出现。流速剪切在高、低潮时不同的时空结构与观测站位处水平盐度梯度密切相关。高潮时, 观测站位处水平盐度梯度较小, 故而斜压压强梯度力不起主要作用。同时, 水体在垂向上混合较为均匀。因此, 高潮时观测站位处流速剪切表现出更多正压潮的特点, 即其峰值被限制在近底层, 且其值随着离底距离的增加而减小。当低潮时, 即潮流由落转涨时, 观测站位处具有明显的水平盐度梯度, 斜压压强梯度力不可忽略且随距底距离的减小而增大。而正压压强梯度力在水体不同层次是相等的。在这样的受力情况下, 底层的水体将较上层的水体先达到受力平衡, 进而发生流向的反转, 流向反转之处随时间增长而离底距离增加。表底流速位相不一致生成强剪切, 剪切峰值自底至表陆续出现。高、低潮位转流时流速剪切不同的时空结构进一步决定了图 4中雷诺应力的深度-时间分布。
本文进一步利用浮力频率和流速剪切数据计算了水体理查德森数(图 5d)。结果显示, 涨潮阶段各层水体理查德森数多大于1.00, 水体稳定。落潮阶段高、低潮时理查德森数具有不同的垂向结构。高潮时, 只有近底层水体理查德森数小于0.25;低潮时, 则是近乎整个水柱理查德森数小于0.25。在高、低潮时刻之间, 有一段时间近底层水体理查德森数大于1.00, 这是由于落潮时低盐水平流至观测站位, 层化增强导致的。尽管理查德森数剖面图 1小时的时间分辨率比较低, 但可以大致看出其与雷诺应力的时空结构是比较吻合的。落潮阶段高潮时, 潮致混合具有正压特点, 雷诺应力峰值被限制于近底层; 落潮阶段低潮时, 潮致混合具有斜压特点, 雷诺应力峰值自底至表陆续出现。在高、低潮之间, 受层化作用影响, 潮致混合较弱。
综上, 斜压作用解释了低潮时雷诺应力峰值自底至表陆续出现以及高、低潮之间雷诺应力量值较小的现象, 但无法解释水体中层雷诺应力信号被明显放大的现象。本文推断雷诺应力峰值出现在水体中层极有可能是受养殖网箱的影响。图 4中雷诺应力峰值出现于养殖网箱底部附近且随着养殖网箱离底距离的减小而逐渐下移的现象可以很好的佐证这一推断。
由于表、中、底三层营养盐浓度变化趋势一致, 本文对其进行了深度平均。结果显示, 南麂列岛养殖功能海域秋季营养盐浓度非常高, 其中SiO32-浓度最高, 平均为20.79μmol/L, 其次为NO3-与PO43-, 平均浓度分别为4.10与1.34μmol/L, NO2-浓度最低, 仅为0.03μmol/L(图 6)。由于NH4+浓度过低, 仪器并未检测出来。观测站位处N/P约为3, 远小于Redfiled比值16, 所以该海域11月份对于浮游植物生长为N限制。总体上来讲, NO3-与盐度呈负相关关系, 即当近岸低盐海水流至观测站位时, NO3-浓度升高, 可达4.6μmol/L, 而当外海高盐水控制观测站位时, NO3-浓度降低至4.0μmol/L以下。由于农业生产过程中大量化肥的使用, 导致东亚河流中NO3-浓度远超过PO43-(Zhang et al, 2007)。受长江冲淡水影响, 观测站位处NO3-浓度与盐度相关性明显。
 |
| 图 6 垂向平均盐度(a)、垂向平均营养盐浓度(b)与最底层、垂向平均雷诺应力(c)时间序列 Fig. 6 Time series of depth-averaged salinity (a), depth-averaged concentrations of nutrients (b), and the lowest-bin Reynolds stress (c) 注:子图(b)、(c)中阴影部分标注了雷诺应力高值发生的时间段 |
NO2-、PO43-与SiO32-等浓度与盐度相关性不大, 说明它们受局地过程影响更大。在雷诺应力量值较大的时间段内(图 6阴影标注范围), 此三类营养盐浓度与雷诺应力均呈现出双峰结构, 说明雷诺应力是对该海域营养盐浓度有重要影响的一个局地动力因子。需要说明的是, 图 6c每一段阴影部分标注的雷诺应力中共有2个峰值, 第一、二个峰值分别主要由正压与斜压过程产生。正压、斜压过程产生的雷诺应力均可造成营养盐浓度的上升。值得注意的是, 7小时左右观测范围内最底层雷诺应力峰值显著, 但垂向平均的雷诺应力峰值并不明显, 即在此时刻养殖网箱对水体中层雷诺应力并无明显增强作用。在这样的情况下, 营养盐浓度仍然明显上升, 说明对营养盐浓度上升起关键作用的是近底层雷诺应力, 而不是养殖网箱的影响。
4 结论南麂列岛深水网箱养殖海域潮流表现出显著的落潮占优现象, 最大涨潮流只有0.15m/s, 而最大落潮流高达0.55m/s。由于落潮阶段存在正压潮向斜压潮的转换过程, 高潮转流时潮流表底位相差不大, 近乎同落; 低潮转流时, 潮流位相差明显, 自底至表陆续转流。与潮流特征相似, 雷诺应力一方面具有落潮占优的特点。涨潮时雷诺应力量值较小, 接近于噪音水平, 而落潮时雷诺应力量值显著增大, 可达8×10-4m2/s2。另一方面在高、低潮转流时呈现出不同的时空结构。高潮转流时, 雷诺应力峰值被限制于近底层; 而在低潮转流时, 其峰值自底至表陆续出现。同时, 养殖网箱对雷诺应力的垂向分布影响明显, 可以显著增强其在水体中层的量值。
南麂列岛深水网箱养殖海域内营养盐浓度很高, 其中SiO32-浓度最高, 平均为20.79μmol/L, 其次为NO3-与PO43-, 平均浓度分别为4.10与1.34μmol/L, NO2-浓度最低, 仅为0.03μmol/L。N/P约为3, 远小于Redfiled比值16, 所以该海域11月份对于浮游植物生长为N限制。NO3-浓度与盐度相关性明显, 说明其受水团控制。SiO32-, PO43-与NO2-则主要受局地过程影响, 雷诺应力对其浓度影响明显。对营养盐浓度变化起关键作用的是近底层雷诺应力, 而非由养殖网箱增强的水体中层雷诺应力。
| 王大海, 2014. 海水养殖业发展规模经济及规模效率研究. 青岛: 中国海洋大学博士学位论文 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D548497 |
| 庄定根, 2014. 南麂岛大黄鱼产业化养殖品质改良技术开发——大黄鱼健康养殖技术研究. 宁波: 宁波大学硕士学位论文 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D604977 |
| 刘素美, 张经, 陈洪涛, 2000. 黄海和东海生源要素的化学海洋学. 海洋环境科学, 19(1): 68–74 |
| 孙建璋, 庄定根, 王铁杆, 等, 2009. 南麂列岛铜藻场建设设计与初步实施. 现代渔业信息, 24(7): 25–28 |
| 林丽华, 2005. 红树林种植-养殖系统内浮游植物及营养盐的动态研究. 广州: 中山大学硕士学位论文 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y767750 |
| 徐鹏, 刘志宇, 毛新燕, 等, 2013. 强潮狭长海湾中垂直涡黏性系数与底拖曳系数的估计. 中国海洋大学学报, 43(8): 1–7 |
| 蔡厚才, 叶鹏, 谢秉笑, 等, 2004. 南麂海区真鲷深水网箱养殖技术研究. 浙江海洋学院学报(自然科学版), 23(4): 347–350, 377 |
| 潘建明, 周怀阳, 扈传昱, 等, 2002. 夏季珠江口沉积物中营养盐剖面分布和界面交换通量. 海洋学报, 24(3): 52–59 |
| 魏皓, 王磊, 林以安, 等, 2002. 黄海中部营养盐的贯跃层输运. 海洋科学进展, 20(3): 15–20 |
| 魏皓, 赵亮, 原野, 等, 2010. 桑沟湾水动力特征及其对养殖容量影响的研究——观测与模型. 渔业科学进展, 31(4): 65–71 |
| Castro L R, Cáceres M A, Silva N, et al, 2011. Short-term variations in mesozooplankton, ichthyoplankton, and nutrients associated with semi-diurnal tides in a Patagonian Gulf. Continental Shelf Research, 31(3-4): 282–292 |
| Chen X Y, Wang X H, Guo J S, 2006. Seasonal variability of the sea surface salinity in the East China Sea during 1990-2002. Journal of Geophysical Research:Oceans, 111(C5): C05008 |
| Jin J, Liu S M, Ren J L, et al, 2013. Nutrient dynamics and coupling with phytoplankton species composition during the spring blooms in the Yellow Sea. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 97: 16–32 |
| Kim J K, Kraemer G, Yarish C, 2008. Field scale evaluation of seaweed aquaculture as a nutrient bioextraction strategy in Long Island Sound and the Bronx River Estuary. Journal of Experimental Marine Biology and Ecology, 30(7): 432–440 |
| Lohrmann A, Hackett B, Røed L P, 1990. High resolution measurements of turbulence, velocity and stress using a pulse-to-pulse coherent sonar. Journal of Atmospheric and Oceanic Technology, 7(1): 19–37 DOI:10.1175/1520-0426(1990)007<0019:HRMOTV>2.0.CO;2 |
| Lu Y Y, Lueck R G, 1999. Using a broadband ADCP in a tidal channel. Part Ⅰ:Mean flow and shear. Journal of Atmospheric and Oceanic Technology, 16(11): 1556–1567 DOI:10.1175/1520-0426(1999)016<1556:UABAIA>2.0.CO;2 |
| Munk W, Wunsch C, 1998. Abyssal recipes Ⅱ:energetics of tidal and wind mixing. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 45(12): 1977–2010 DOI:10.1016/S0967-0637(98)00070-3 |
| Rippeth T P, Williams E, Simpson J H, 2002. Reynolds stress and turbulent energy production in a tidal channel. Journal of Physical Oceanography, 32(4): 1242–1251 DOI:10.1175/1520-0485(2002)032<1242:RSATEP>2.0.CO;2 |
| Santoro A E, Casciotti K L, Francis C A, 2008. Activity, abundance and diversity of nitrifying archaea and bacteria in the central California Current. Environmental Microbiology, 12(7): 1989–2006 |
| Sharples J, Moore C M, Abraham E R, 2001. Internal tide dissipation, mixing, and vertical nitrate flux at the shelf edge of NE New Zealand. Journal of Geophysical Research:Oceans, 106(C7): 14069–14081 DOI:10.1029/2000JC000604 |
| Souza A J, Pineda J, 2001. Tidal mixing modulation of sea-surface temperature and diatom abundance in Southern California. Continental Shelf Research, 21(6-7): 651–666 DOI:10.1016/S0278-4343(00)00105-9 |
| Stacey M T, Monismith S G, Burau J R, 1999. Measurements of Reynolds stress profiles in unstratified tidal flow. Journal of Geophysical Research:Oceans, 104(C5): 10933–10949 DOI:10.1029/1998JC900095 |
| Van Ginkel S W, Ahn C H, Badruzzaman M, et al, 2008. Kinetics of nitrate and perchlorate reduction in ion-exchange brine using the membrane biofilm reactor (MBfR). Water Research, 42(15): 4197–4205 DOI:10.1016/j.watres.2008.07.012 |
| Van Verseveld W J, McDonnell J J, Lajtha K, 2009. The role of hillslope hydrology in controlling nutrient loss. Journal of Hydrology, 367(3-4): 177–187 DOI:10.1016/j.jhydrol.2008.11.002 |
| Wang J N, Greenan B J W, Lu Y Y, et al, 2014. Layered mixing on the New England Shelf in summer. Journal of Geophysical Research, 119(9): 5776–5796 |
| Williams E, Simpson J H, 2004. Uncertainties in estimates of Reynolds stress and TKE production rate using the ADCP variance method. Journal of Atmospheric and Oceanic Technology, 21(2): 347–357 DOI:10.1175/1520-0426(2004)021<0347:UIEORS>2.0.CO;2 |
| Zhang J, Liu S M, Ren J L, et al, 2007. Nutrient gradients from the eutrophic Changjiang (Yangtze River) Estuary to the oligotrophic Kuroshio waters and re-evaluation of budgets for the East China Sea shelf. Progress in Oceanography, 74(4): 449–478 DOI:10.1016/j.pocean.2007.04.019 |