 2018, Vol. 49
2018, Vol. 49

2004-2016年夏季长江口网采浮游植物及其影响因素

http://dx.doi.org/10.11693/hyhz20171100289
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 黄海燕, 王秋璐, 许艳, 刘捷, 曲艳敏. 2018.
- HUANG Hai-Yan, WANG Qiu-Lu, XU Yan, LIU Jie, QU Yan-Min. 2018.
- 2004-2016年夏季长江口网采浮游植物及其影响因素
- DISTRIBUTION OF SUMMER COMMUNITY OF NET-COLLECTED PHYTOPLANKTON FROM 2004 TO 2016 AND THE FACTORS IN THE CHANGJIANG RIVER ESTUARY
- 海洋与湖沼, 49(2): 319-330
- Oceanologia et Limnologia Sinica, 49(2): 319-330.
- http://dx.doi.org/10.11693/hyhz20171100289
-
文章历史
- 收稿日期:2017-11-14
- 收修改稿日期:2017-11-27
引用本文
黄海燕, 王秋璐, 许艳, 刘捷, 曲艳敏. 2018. 2004-2016年夏季长江口网采浮游植物及其影响因素[J]. 海洋与湖沼, 49(2): 319-330
HUANG Hai-Yan, WANG Qiu-Lu, XU Yan, LIU Jie, QU Yan-Min. 2018. DISTRIBUTION OF SUMMER COMMUNITY OF NET-COLLECTED PHYTOPLANKTON FROM 2004 TO 2016 AND THE FACTORS IN THE CHANGJIANG RIVER ESTUARY[J]. Oceanologia et Limnologia Sinica, 49(2): 319-330.
2004-2016年夏季长江口网采浮游植物及其影响因素
国家海洋信息中心 天津 300171
摘要:基于2004-2016年夏季长江口生态监控区13年的生态环境调查数据,开展网采浮游植物年际变化及其与环境因子关系研究。结果表明:(1)共鉴定出浮游植物7门263种,以硅藻占绝对优势,甲藻次之,物种数呈增加趋势,其中硅藻种类数占有比下降,而甲藻的种类数占有比呈上升趋势;(2)总细胞丰度、硅藻细胞丰度和甲藻细胞丰度都呈上升趋势,除2014年外,其余年份硅藻细胞丰度都占绝对优势并与总细胞丰度的分布曲线和变化趋势基本一致;(3)水平分布模式及其年际变化受环流变化和水团消长的影响,总体趋势是河口区细胞丰度较低,由河口到外海呈逐渐增大趋势,部分年份呈现先增大后减小的趋势;(4)生物多样性多年呈下降趋势,物种组成随空间和时间变化较大,只有较少的种类能够保持稳定,多数物种只是偶尔被检出;(5)中肋骨条藻有10年为区域第一优势种,41种常见种中有31种细胞丰度呈增加趋势,10种细胞丰度呈下降趋势。(6)浮游植物细胞丰度与活性磷酸盐和无机氮具有相关性,种类数、多样性指数除了与溶解氧、氨氮、叶绿素a无相关性外与其他10种环境因子都相关。本研究能够较好反映研究区域浮游植物的本底情况及演变趋势。
关键词:长江口 浮游植物 年际变化 影响因素
DISTRIBUTION OF SUMMER COMMUNITY OF NET-COLLECTED PHYTOPLANKTON FROM 2004 TO 2016 AND THE FACTORS IN THE CHANGJIANG RIVER ESTUARY
National Marine Data and Information Service, Tianjin 300171, China
Abstract:
Based on ecological environment survey data collected in the Changjiang (Yangtze) River estuary ecological monitoring area in summer from 2004 to 2016, the interannual variation of net-collected phytoplankton and its relationship to environmental factors were studied. Seven phyla including 263 species of phytoplankton were identified and dominated by diatoms, followed by dinoflagellates. In the past 13 years, the number of species tended to increase; the contribution of diatom diversity to total phytoplankton diversity declined, and the proportion of dinoflagellates diversity increased correspondingly. The cell abundance of the total species, including diatom and dinoflagellates displayed an increasing trend. The diatom cell abundance dominated except for 2014, and the distribution and variation patterns were consistent with the total cell abundance in the rest of the year. The horizontal distribution pattern of cell abundance and its interannual variation were affected by circulation change and water consumption. The cell abundance was relatively low in the estuary and increased toward the open sea; the overall cell abundance increased gradually but fluctuated in some years. The biodiversity slightly decreased, and the community composition changed greatly in temporal and spatial scopes. Only a few species remained stable during the period, and most could be detected occasionally. Skeletonema costatum was dominant species in at least 10 years. Of 41 common species, 31 showed an increasing trend and 10 decreased in cell abundance. In addition, phytoplankton cell abundance was correlated with phosphate and inorganic nitrogen. The species number and diversity index were related to 10 out of 13 studied environmental factors, exclusive of dissolved oxygen, ammonia nitrogen, and chlorophyll a. This study would be of great significance to understand the current situation and evolution trend of phytoplankton.
Key words:
Changjiang River estuary
phytoplankton
interannual variation
influential factors
长江口生态监控区是全国范围内实施生态监控区制度的18个典型生态系统之一。监控区位于上海市近岸海域, 作为河口与海洋的交汇区域, 受长江冲淡水、沿岸流、台湾暖流和黑潮的影响, 加上航道整治、滩涂围垦以及长江上游大型水利工程建设引起的水动力条件变化, 造就了复杂多变的环境。长江流域承载流经地区的工、农业废水和城市污水, 将大量生源要素(碳、氮、磷等)、有机污染物、重金属等从陆地带入长江口海域, 对该区域生态环境造成了极大的影响, 水体富营养化严重、赤潮频发(周名江等, 2003; 范丽等, 2008; 张璇, 2012; 尹艳娥等, 2014; 张桂成, 2015)。
浮游植物是赤潮发生的生物基础, 作为初级生产者, 其种群变动和群落结构直接影响海洋生态系统的结构和功能。对长江口及其毗邻海域浮游植物研究的报导已有很多, 内容涵盖种类组成、数量分布、生物多样性、浮游植物群落与环境因子关系等(Gao et al, 2005; Kong et al, 2009; Zhu et al, 2009; Yang et al, 2014; Jiang et al, 2015; 李云等, 2007; 栾青杉等, 2007; 王丹等, 2008; 何青等, 2009; 章飞燕, 2009; 栾青杉等, 2010; 唐峰华等, 2010; 王江涛等, 2012; 赵冉等, 2013; 林峰竹等, 2008; 江志兵等, 2014; 刘海娇等, 2015; 林梅等, 2017), 但由于受到样本和数据的限制, 这些研究主要是基于一次调查或者是短时期内群落结构变化, 长期趋势变化分析只能依赖历史文献材料收集, 所分析的材料往往存在不同年份调查区域、调查时间不一致的情况, 即使是在同一年份, 也存在调查站位个数有限, 调查资料不完整等问题, 不足以反映研究区域浮游植物多年连续变化情况。本文基于研究区域连续13年同一时间段的大面调查数据, 开展浮游植物种类组成、物种变化、数量变化、群落特征指数及其变化趋势研究, 并结合水深、盐度、pH、溶解氧、无机氮、活性磷酸盐、活性硅酸盐等13种环境因子开展相关性分析, 长期连续的年际调查资料是研究浮游植物年际变化的珍贵数据资源, 可以弥补以往研究的不足。本研究有助于摸清长江口生态监控区浮游植物的本底情况及演变趋势, 为赤潮发生机制及生态系统变化研究提供基础资料。
1 材料与方法 1.1 材料浮游植物数据来源于2004—2016年海洋环境监测长江口生态监控区8月份由底至表垂直拖网调查, 网型为Ⅲ型浮游生物网(网目孔径76μm), 鉴定分析方法按《海洋监测规范》(中华人民共和国国家质量监督检验检疫总局, 2007)第7部分执行, 采用浓缩计数法计数。数据空间范围为121.32°—123°E, 30.75°— 31.75°N, 除2011和2012年站位为17个, 其余各年份站位都为20个, 各年份站位在空间上分布上略有差异, 共计30个不重复站位, 其中有8、3、7、1、1、1、1、4、4个站位分别调查了13年、12年、11年、10年、9年、4年、3年、2年、1年数据。如图 1所示。
 |
| 图 1 2004—2016年夏季网采浮游植物站位分布图 Fig. 1 Phytoplankton net-sampling stations in summer from 2004 to 2016 |
水质数据来源于相应站位调查, 水质因子分析均按《海洋监测规范》执行, 包括水深、盐度、pH、溶解氧、悬浮物、化学耗氧量、活性磷酸盐、氨氮、硝酸盐、亚硝酸盐、无机氮、活性硅酸盐和叶绿素a。
1.2 数据处理分析本文以物种站位检出率(F)、丰度百分比(C)研究浮游植物物种变化特征。
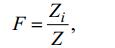 (1)
(1)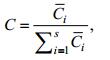 (2)
(2)Zi为第i种出现的站位数, Z为总站位数; 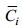
本文以区域物种优势度Y最大的物种为第一优势种(简称优势种), 物种多样性指数H'(Shannon- Wiener index)、物种丰富度指数D (Margalef’s index)和均匀度指数J(Pielou index)研究浮游植物群落特征。
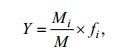 (3)
(3)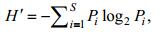 (4)
(4)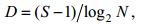 (5)
(5)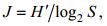 (6)
(6)Mi表示所有样品中第i种的总个体数, M表示所有样品中所有物种的总个体数, fi为第i种在所有样品中出现的频率; S表示样品中的种类数, N表示样品中物种总个体数, Pi表示第i种在该样品中占的比例。
群落与环境因子分析主要利用多元统计分析软件SPSS12进行Pearson相关性分析。
2 结果 2.1 种类组成2004—2016年长江口生态监控区夏季网采浮游植物(以下简称浮游植物)共鉴定出7门263种(定种224种), 其中硅藻171种(定种149种)、甲藻62种(定种59种)、金藻4种(定种3种)、红藻1种、蓝藻12种(定种5种)、裸藻1种、绿藻12种(定种8种)。各年份种类组成上硅藻占绝对优势, 占浮游植物种类数62%—74%, 呈下降趋势; 甲藻占浮游植物种类数19%—29%, 呈上升趋势。2004—2016年物种变化范围为60—113种, 2004和2008年最高, 2009年最低, 各年份物种数在波动中呈增加趋势(图 2)。
 |
| 图 2 2004—2016年夏季浮游植物种类变化 Fig. 2 Variation of phytoplankton species in summer from 2004 to 2016 |
2004—2016年各站位浮游植物物种数统计如图 3所示, 各站位物种变化范围为1—58种, 2012年各站位间物种数差异最大, 为54种, 2007年各站位间物种数差异最小, 为22种, 2004年有一个离群值, 为51种。
 |
| 图 3 2004—2016年夏季各站位浮游植物种类数变化 Fig. 3 Variation in the number of phytoplankton species in the stations in summer from 2004 to 2016 |
2004—2016年夏季长江口各浮游植物物种年检出状况如图 4所示, 所有年份都检出的物种有18种占6.8%, 包括硅藻12种, 甲藻5种和蓝藻1种; 出现5—12年的物种有85种, 占32%;出现1—4年的物种有160种, 占61%, 其中仅出现1年的物种有88种。
 |
| 图 4 2004—2016年夏季浮游植物各物种年检出数 Fig. 4 The time of presence of phytoplankton species in summer from 2004 to 2016 |
近13年, 各物种站位检出率在0.4%—85%;站位检出率≥50%的物种有11种, 由硅藻和甲藻组成, 其中中肋骨条藻和琼氏圆筛藻站位检出率在80%以上, 洛氏角毛藻、虹彩圆筛藻和纺锤角藻站位检出率在60%以上; 站位检出率<10%的共有212种, 占全部物种数的80.6%, 站位检出率<1%的有185种, 占70%。各物种站位检出率如图 5所示。
 |
| 图 5 2004—2016年夏季各浮游植物站位检出率 Fig. 5 The frequency of occurrence of phytoplankton species in summer from 2004 to 2016 |
近13年中检出年份数≥10年的物种有41种, 这些常见种的检出年份数、站位检出率、丰度百分比及各年份各物种细胞丰度及变化情况如表 1所示。这些常见浮游植物种包括硅藻29种, 甲藻10种, 蓝藻和绿藻各1种, 站位检出率范围7%—85%, 站位检出率≥50%的有11种, 丰度百分比范围从 < 0.01%到93.6%, 以中肋骨条藻占绝对优势, 站位检出率和丰度百分比都为最大, 丰度百分比≤0.01%的有25种。各常见浮游植物中细胞丰度呈增加趋势的有31种, 呈下降趋势的10种。中肋骨条藻、旋链角毛藻和尖刺拟菱形藻物种的丰度百分比之和达96.1%, 因此, 其余260种浮游植物细胞丰度之和小于4%。
表 1 2004—2016年浮游植物常见物种的细胞数量年际变化(单位: cells/m3) Tab. 1 Annual variation in the cell numbers of common phytoplankton species in summer from 2004 to 2016 (unit: cells/m3)
| 物种名称 | 年份数 | 站检出率(%) | 丰度百分比(%) | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 趋势 | ||
| 硅藻门 |  |
 |
|||||||||||||||||
| 中肋骨条藻Skeletonema costatum | 13 | 85 | 93.6 | ||||||||||||||||
| 琼氏圆筛藻Coscinodiscus jonesianus | 13 | 83 | 0.11 | ||||||||||||||||
| 虹彩圆筛藻Coscinodiscus oculus-iridis | 13 | 63 | 0.02 | ||||||||||||||||
| 洛氏角毛藻Chaetoceros lorenzianus | 13 | 61 | 0.45 | ||||||||||||||||
| 菱形海线藻Thalassionema nitzschioides | 13 | 45 | 0.15 | ||||||||||||||||
| 旋链角毛藻Chaetoceros curvisetus | 13 | 41 | 1.06 | ||||||||||||||||
| 布氏双尾藻Ditylum brightwellii | 13 | 35 | 0.01 | ||||||||||||||||
| 扁面角毛藻Chaetoceros compressus | 13 | 31 | 0.37 | ||||||||||||||||
| 丹麦细柱藻Leptocylindrus danicus | 13 | 20 | 0.01 | ||||||||||||||||
| 圆海链藻Thalassiosira rotula | 13 | 20 | 0.01 | ||||||||||||||||
| 扭鞘藻Streptotheca thamesis | 13 | 20 | < 0.01 | ||||||||||||||||
| 蜂窝三角藻Triceratium favus | 13 | 18 | < 0.01 | ||||||||||||||||
| 伏氏海毛藻Thalassiothrix frauenfeldii | 12 | 59 | 0.07 | ||||||||||||||||
| 中华盒形藻Biddulphia sinensis | 12 | 50 | 0.01 | ||||||||||||||||
| 透明辐杆藻Bacteriastrum hyalinum | 12 | 31 | 0.02 | ||||||||||||||||
| 窄隙角毛藻Chaetoceros affinis | 12 | 24 | 0.08 | ||||||||||||||||
| 颗粒直链藻Melosira granulata | 12 | 20 | 0.03 | ||||||||||||||||
| 刚毛根管藻Rhizosolenia setigera | 11 | 30 | < 0.01 | ||||||||||||||||
| 笔尖形根管藻Rhizosolenia styliformis | 11 | 23 | 0.01 | ||||||||||||||||
| 冕孢角毛藻Chaetoceros diadema | 11 | 13 | 0.05 | ||||||||||||||||
| 小细柱藻Leptocylindrus minimus | 11 | 11 | < 0.01 | ||||||||||||||||
| 具槽直链藻Melosira sulcata | 11 | 9 | < 0.01 | ||||||||||||||||
| 尖刺拟菱形藻Pseudo-nitzschia pungens | 10 | 50 | 1.48 | ||||||||||||||||
| 齿角毛藻Chaetoceros denticulatus | 10 | 18 | 0.01 | ||||||||||||||||
| 短角弯角藻Eucampia zoodiacus | 10 | 17 | 0.12 | ||||||||||||||||
| 翼根管藻纤细变型Rhizosolenia alata f. gracillima | 10 | 17 | < 0.01 | ||||||||||||||||
| 爱氏角毛藻Chaetoceros eibenii | 10 | 12 | < 0.01 | ||||||||||||||||
| 有翼圆筛藻Coscinodiscus bipartitus | 10 | 9 | < 0.01 | ||||||||||||||||
| 密聚角毛藻Chaetoceros coarctatus | 10 | 7 | < 0.01 | ||||||||||||||||
| 甲藻门 | |||||||||||||||||||
| 纺锤角藻Ceratium fusus | 13 | 65 | 0.07 | ||||||||||||||||
| 三角角藻Ceratium tripos | 13 | 57 | 0.01 | ||||||||||||||||
| 夜光藻Noctiluca scintillans | 13 | 52 | 0.02 | ||||||||||||||||
| 叉角藻Ceratium furca | 13 | 52 | 0.01 | ||||||||||||||||
| 具尾鳍藻Dinophysis caudata | 13 | 46 | < 0.01 | ||||||||||||||||
| 五角多甲藻Peridinium pentagonum | 12 | 28 | < 0.01 | ||||||||||||||||
| 锥形多甲藻Peridinium conicum | 12 | 27 | < 0.01 | ||||||||||||||||
| 扁平多甲藻Peridinium depressum | 11 | 25 | < 0.01 | ||||||||||||||||
| 三叉角藻Ceratium trichoceors | 10 | 13 | < 0.01 | ||||||||||||||||
| 锥状斯克里普藻Scrippsiella trochoidea | 10 | 12 | < 0.01 | ||||||||||||||||
| 蓝藻门 | |||||||||||||||||||
| 微囊藻Microcystis sp. | 13 | 15 | < 0.01 | ||||||||||||||||
| 绿藻门 | |||||||||||||||||||
| 单角盘星藻具孔变种 | 11 | 14 | 0.01 | ||||||||||||||||
| Pediastrum simplex var. duodenarium | |||||||||||||||||||
注:通过线性回归分析物种细胞数量变化趋势, “  ”代表斜率k > 0; “↘”代表斜率k < 0; ”代表斜率k > 0; “↘”代表斜率k < 0;  | |||||||||||||||||||
2004—2016年夏季长江口浮游植物平均细胞丰度范围为(110—93030)×104cells/m3, 硅藻平均细胞丰度范围为(104—93021)×104cells/m3, 甲藻平均细胞丰度范围为(2—94)×104cells/m3。长江口浮游植物平均细胞丰度、硅藻平均细胞丰度和甲藻平均细胞丰度都呈上升趋势, 总细胞丰度在2012年最高, 2004和2014年最低。除2014年外, 其余年份硅藻细胞丰度都占绝对优势, 与总细胞丰度分布曲线和变化趋势基本一致(图 6)。
 |
| 图 6 2004—2016年夏季浮游植物细胞丰度 Fig. 6 Cell abundance of phytoplankton in summer from 2004 to 2016 |
2004—2016年夏季长江口各站位浮游植物细胞丰度分布情况见图 7, 明显可见2016、2012、2009和2008年各站位细胞丰度水平较其他年份略高, 2015、2014和2004年各站位细胞丰度水平较低。2011年各站位细胞丰度水平差异较大, 有5个站位细胞丰度明显高于其他站位。各站位细胞丰度分布状况由河口到外海呈羽状分布, 总体呈逐渐增大趋势, 部分年份呈现先增大后减小的趋势。
 |
| 图 7 2004—2016年夏季网采浮游植物细胞丰度分布 Fig. 7 Distribution in cell abundance of net-phytoplankton in summer from 2004 to 2016 |
中肋骨条藻、旋链角毛藻、纺锤角藻和尖刺拟菱形藻为2004—2016年夏季长江口浮游植物优势种(表 2), 各年份优势种优势度范围为0.2—0.89, 细胞丰度范围为(49—92955)×104cells/m3, 站位检出率范围35%—100%, 丰度百分比范围为31%—99.9%。13年中, 中肋骨条藻有10年为区域第一优势种, 且在2006、2007、2008、2011、2012和2016年处于绝对优势, 旋链角毛藻、纺锤角藻和尖刺拟菱形藻分别在2013、2014和2015年成为该区域第一优势种, 且优势度都较低。
表 2 2004—2016年夏季网采浮游植物优势种 Tab. 2 Dominant species of net-phytoplankton in summer from 2004 to 2016
| 年份 | 优势种 | 优势度 | 细胞丰度(104cells/m3) | 站位检出率(%) | 丰度百分比(%) |
| 2004 | 中肋骨条藻 | 0.35 | 49 | 80 | 44 |
| 2005 | 中肋骨条藻 | 0.63 | 2387 | 90 | 70 |
| 2006 | 中肋骨条藻 | 0.72 | 1666 | 85 | 85 |
| 2007 | 中肋骨条藻 | 0.79 | 3414 | 85 | 93 |
| 2008 | 中肋骨条藻 | 0.84 | 5656 | 100 | 84 |
| 2009 | 中肋骨条藻 | 0.39 | 1604 | 85 | 61 |
| 2010 | 中肋骨条藻 | 0.26 | 182 | 85 | 31 |
| 2011 | 中肋骨条藻 | 0.88 | 92955 | 88 | 99.9 |
| 2012 | 中肋骨条藻 | 0.79 | 11973 | 94 | 84 |
| 2013 | 旋链角毛藻 | 0.2 | 461 | 35 | 58 |
| 2014 | 纺锤角藻 | 0.28 | 66 | 75 | 38 |
| 2015 | 尖刺拟菱形藻 | 0.34 | 387 | 75 | 45 |
| 2016 | 中肋骨条藻 | 0.89 | 12044 | 95 | 94 |
2004—2016年长江口物种多样性指数范围为0.73—2.28, 丰富度指数范围0.51—1.28, 均匀度指数范围0.21—0.6;各站位物种多样性指数范围为0— 4.03, 丰富度指数范围0—3.04, 均匀度指数范围0— 0.99(图 8)。站位多样性指数0值出现在2009年和2011年, 因其中某些站位只检测到一个物种或者某一物种细胞数量太大造成多样性指数接近0值。2009、2010、2012和2016年各站位物种多样性指数分布呈左偏, 2004和2010年呈右偏, 其余年份分布较对称; 均匀度指数在2010、2012和2015年出现离群值, 丰富度指数在2004、2009和2016年出现离群值。经线性回归分析, 物种多样性指数和均匀度指数在波动中总体程下降趋势, 丰富度指数在波动中总体呈上升趋势。
 |
| 图 8 2004—2016年夏季网采浮游植物多样性指数H' (a), 均匀度指数J (b), 和丰富度指数D (c) Fig. 8 Diversity indexes of net-phytoplankton in summer from 2004 to 2016 a. H' (Shannon-Wiener index); b. J (Pielou index); c. D (Margalef's index) |
利用SPSS12分析软件对各个站位的浮游植物细胞丰度、硅藻细胞丰度和甲藻细胞丰度、种类数、物种多样性指数、丰富度指数、均匀度指数与水深、盐度、pH、溶解氧、悬浮物、化学耗氧量、活性磷酸盐、氨氮、硝酸盐、亚硝酸盐、无机氮、活性硅酸盐和叶绿素a共13项相关环境因子开展进行相关性分析(表 3)。结果表明, 近13年夏季长江口网采浮游植物细胞丰度与活性磷酸盐、硝酸盐和无机氮正相关; 甲藻细胞丰度与盐度正相关, 与活性磷酸盐、氨氮、硝酸盐、无机氮呈负相关, 与活性硅酸盐呈显著负相关; 硅藻细胞丰度与活性磷酸盐、硝酸盐和无机氮正相关; 浮游植物各站位物种数与水深、盐度、pH显著正相关, 与悬浮物、化学耗氧量、活性磷酸盐、硝酸盐、无机氮和活性硅酸盐呈显著负相关关系, 与亚硝酸盐呈负相关关系; 物种多样性指数与水深、盐度显著正相关, 与悬浮物、化学耗氧量、活性磷酸盐、硝酸盐、亚硝酸盐、无机氮呈显著负相关关系, 与活性硅酸盐呈负相关关系; 丰富度指数与水深、盐度、pH显著正相关, 与悬浮物、化学耗氧量、活性磷酸盐、硝酸盐、亚硝酸盐、无机氮和活性硅酸盐呈显著负相关关系; 均匀度指数与盐度呈正相关, 与化学耗氧量、亚硝酸盐呈显著负相关, 与无机氮呈负相关关系。
表 3 浮游植物群落与环境参数Pearson相关系数 Tab. 3 The Pearson correlation coefficient of phytoplankton abundance and environmental factors
| 总细胞丰度 | 甲藻细胞丰度 | 硅藻细胞丰度 | 物种数 | 物种多样性指数 | 丰富度指数 | 均匀度指数 | ||
| 水深 | Pearson相关性 | –0.065 | 0.093 | –0.065 | 0.547** | 0.303** | 0.487** | 0.161 |
| 显著性(双侧) | 0.464 | 0.292 | 0.464 | 0.000 | 0.000 | 0.000 | 0.068 | |
| N | 130 | 130 | 130 | 130 | 130 | 130 | 130 | |
| 盐度 | Pearson相关性 | –0.125 | 0.169* | –0.125 | 0.613** | 0.379** | 0.585** | 0.199* |
| 显著性(双侧) | 0.108 | 0.029 | 0.108 | 0.000 | 0.000 | 0.000 | 0.010 | |
| N | 166 | 166 | 166 | 166 | 166 | 166 | 166 | |
| pH | Pearson相关性 | –0.057 | 0.060 | –0.057 | 0.332** | 0.135 | 0.239** | 0.045 |
| 显著性(双侧) | 0.520 | 0.496 | 0.520 | 0.000 | 0.126 | 0.006 | 0.615 | |
| N | 130 | 130 | 130 | 130 | 130 | 130 | 130 | |
| 溶解氧 | Pearson相关性 | 0.125 | –0.126 | 0.125 | –0.004 | –0.126 | –0.091 | –0.126 |
| 显著性(双侧) | 0.087 | 0.084 | 0.087 | 0.957 | 0.084 | 0.212 | 0.084 | |
| N | 189 | 189 | 189 | 189 | 189 | 189 | 189 | |
| 悬浮物 | Pearson相关性 | 0.116 | –0.101 | 0.116 | –0.490** | –0.232** | –0.478** | 0.019 |
| 显著性(双侧) | 0.111 | 0.168 | 0.110 | 0.000 | 0.001 | 0.000 | 0.794 | |
| N | 189 | 189 | 189 | 189 | 189 | 189 | 189 | |
| 总细胞丰度 | 甲藻细胞丰度 | 硅藻细胞丰度 | 物种数 | 物种多样性指数 | 丰富度指数 | 均匀度指数 | ||
| 化学耗氧量 | Pearson相关性 | 0.055 | –0.004 | 0.055 | –0.251** | –0.348** | –0.335** | –0.301** |
| 显著性(双侧) | 0.571 | 0.968 | 0.571 | 0.008 | 0.000 | 0.000 | 0.001 | |
| N | 110 | 110 | 110 | 110 | 110 | 110 | 110 | |
| 活性磷酸盐 | Pearson相关性 | 0.175* | –0.180* | 0.175* | –0.571** | –0.276** | –0.505** | –0.104 |
| 显著性(双侧) | 0.016 | 0.013 | 0.016 | 0.000 | 0.000 | 0.000 | 0.156 | |
| N | 189 | 189 | 189 | 189 | 189 | 189 | 189 | |
| 氨氮 | Pearson相关性 | 0.029 | 0.195* | 0.029 | –0.113 | –0.125 | –0.126 | –0.108 |
| 显著性(双侧) | 0.706 | 0.011 | 0.706 | 0.145 | 0.108 | 0.104 | 0.164 | |
| N | 168 | 168 | 168 | 168 | 168 | 168 | 168 | |
| 硝酸盐 | Pearson相关性 | 0.177* | –0.158* | 0.177* | –0.519** | –0.250** | –0.471** | –0.071 |
| 显著性(双侧) | 0.016 | 0.031 | 0.016 | 0.000 | 0.001 | 0.000 | 0.333 | |
| N | 186 | 186 | 186 | 186 | 186 | 186 | 186 | |
| 亚硝酸盐 | Pearson相关性 | –0.024 | 0.010 | –0.024 | –0.161* | –0.200** | –0.192** | –0.203** |
| 显著性(双侧) | 0.748 | 0.895 | 0.748 | 0.027 | 0.006 | 0.008 | 0.005 | |
| N | 189 | 189 | 189 | 189 | 189 | 189 | 189 | |
| 无机氮 | Pearson相关性 | 0.174* | –0.162* | 0.174* | –0.621** | –0.348** | –0.583** | –0.175* |
| 显著性(双侧) | 0.023 | 0.035 | 0.023 | 0.000 | 0.000 | 0.000 | 0.022 | |
| N | 170 | 170 | 170 | 170 | 170 | 170 | 170 | |
| 活性硅酸盐 | Pearson相关性 | 0.047 | –0.233** | 0.047 | –0.567** | –0.201* | –0.483** | –0.005 |
| 显著性(双侧) | 0.601 | 0.008 | 0.600 | 0.000 | 0.022 | 0.000 | 0.954 | |
| N | 129 | 129 | 129 | 129 | 129 | 129 | 129 | |
| 叶绿素a | Pearson相关性 | 0.007 | 0.106 | 0.007 | 0.115 | –0.050 | 0.027 | –0.087 |
| 显著性(双侧) | 0.921 | 0.145 | 0.921 | 0.114 | 0.490 | 0.708 | 0.235 | |
| N | 190 | 190 | 190 | 190 | 190 | 190 | 190 | |
| 注: N:样本数; **: P < 0.01极显著相关; *: P < 0.05显著相关 | ||||||||
本文对2004—2016年长江口夏季网采浮游植物数据开展分析, 共计254组浮游植物数据, 并采用与之对应站位的190组水质各参数数据开展相关性分析, 缺少的为2011年部分站位及2012—2014年全部站位的水质数据, 主要原因是由于方案的调整, 未对对应站位开展水质调查。
种类组成上, 硅藻占绝对优势, 甲藻次之, 此外还有少量的金藻、红藻、蓝藻、裸藻和绿藻。近13年物种数在波动中呈增加趋势, 硅藻种类数占有比例下降, 而甲藻的种类数占有比呈上升趋势。王金辉等(2004)分析了1990—2002年间长江口海域浮游植物的年际变化, Jiang等(2014)研究了过去50年长江口海域浮游植物的演替, 林梅等(2017)分析了1986—2014年春季长江口海域浮游植物年际变化, 虽然在各自研究时间段中浮游植物种类数变化趋势有所差异, 但都一致认为, 硅藻种类数占有比呈下降趋势, 而甲藻的种类数占有比呈上升趋势。近半个世纪以来, 长江口及其邻近海域悬沙含量降低, 增加了水体的透明度, 氮、磷营养盐含量也明显增加, 而硅浓度显著降低(辛明, 2014)。透明度和氮、磷营养盐含量的增加有利于浮游植物的生长, 促进种类数的上升, 而硅浓度的降低将限制硅藻群落的发展, 使得硅藻种类占有比下降, 再加上近些年对甲藻研究鉴定技术的进步, 发现了更多种类的甲藻, 使得甲藻种类占有比上升。
各年际间物种数和年内各站位物种数差异都较大, 10年都有出现的物种占15.6%, 出现1—4年次的物种占61%, 站位检出率≥50%的物种占4.2%, 站位检出率<1%的物种占70%。中肋骨条藻在2004— 2012年、2016年皆为本研究区域的第一优势种, 旋链角毛藻和尖刺拟菱形藻分别在2013和2015年成为第一优势种, 这三种硅藻的细胞丰度百分比之和达96.1%, 因此, 其余260种浮游植物细胞丰度之和小于4%。由此可见, 长江口夏季浮游植物物种组成随空间和时间变化较大, 只有较少的种类能够保持稳定, 多数物种因调查时间和区域不同而只是偶尔被检出, 这与长江口复杂的生态环境有关。根据盐度、光照、流沙等水体理化性质, 长江河口生态系统自西向东划分为长江口咸淡水生态系(河口区, 122°E以西)、河口半咸水生态系(冲淡水区122°—122.5°E)、外海高盐生态系(外海区, 122.5°E以东), 其范围随着丰水期和枯水期有所变化(王金辉, 2002), 只有广适性的浮游植物才能在如此复杂的环境中占据主导地位。中肋骨条藻为广温广盐的典型代表种, 旋链角毛藻尖刺拟菱形藻亦为广温性沿岸种, 这三种硅藻皆为小型的链状群体种类(郭皓, 2004), 它们的生理特性使其能够适应长江口复杂的环境, 个体微小营群体生活, 使其在竞争中更占优势。
资料显示(国家海洋局, 2005, 2006, 2007, 2008, 2009, 2010, 2011, 2012, 2013, 2014, 2015, 2016, 2017), 长江口海域为在赤潮高发区, 2004—2016年每年都有赤潮发生, 引发赤潮的生物种主要为中肋骨条藻、东海原甲藻、米氏凯伦藻、红色中益虫和海链藻。中肋骨条藻在2005、2007、2008、2010、2011、2015、2016年发生赤潮, 东海原甲藻在2004、2006、2010、2012、2013、2014、2016年发生赤潮, 米氏凯伦藻在2006、2012、2013年发生赤潮。本研究数据中显示中肋骨条藻在各年份都有检测出且细胞丰度大, 说明该区域有发生中肋骨条藻赤潮的生物基础。而东海原甲藻仅出现在2007和2013年, 米氏凯伦藻仅在2007年被检测出, 一方面可能是由于赤潮发生时间主要集中在5—6月份, 8月份东海原甲藻和米氏凯伦藻由于密度较低所以在网采样品中未检测到; 另一方面是由于对东海原甲藻和米氏凯伦藻等单细胞浮游植物, Ⅲ型网对其捕获效率比较差, 网采数据在反应其实际数量方面存在不足。
硅藻在调查海域细胞丰度上占优势, 除2014年外, 整个调查海域浮游植物细胞丰度基本与硅藻细胞丰度呈现一致的分布趋势。在水平分布上, 不同年份有所差异, 总体趋势是河口区细胞丰度较低, 由河口到外海呈逐渐增大趋势, 部分年份呈现先增大后减小的趋势。浮游植物的细胞丰度的分布模式与长江冲淡水、台湾暖流水等水系的影响范围以及不同浮游植物物种的生理生态习性有较大的关系(栾青杉等, 2010)。研究发现(郭术津, 2012), 长江口近岸海域浮游植物细胞丰度高值区多集中于海水盐度为15—23的海域, 并且随长江冲淡水的扩展和回缩东西偏移, 再加上不同的年份东海的水团分布有所差别, 如长江径流量的年度变化、台湾暖流北上的范围以及黑潮对东海大陆架入侵的年度差异等, 这些差异会明显影响长江口海域理化因子如温度、盐度和营养盐等的分布趋势, 从而造成长江口细胞丰度分布模式的差异。
在群落结构上, 虽然近13年物种多样性指数和均匀度指数均在波动中总体呈下降趋势, 但物种的丰富度呈增加趋势, 各年份内各站位间多样性指数差异较大。一般而言, 物种多样性指数越高、种类丰富度越大、个体分布越均匀, 代表这个海洋生态系统越健康, 而在本研究中, 丰富度指数和其它多样性指数变化趋势不同, 主要原因在于长江口是典型的河口区, 浮游植物优势种和其它物种的丰度相差很悬殊, 导致物种丰富度指数失去了对群落多样性的判别能力(孙军等, 2004)。与环境因子的相关性表明, 多样性指数与悬浮物、化学耗氧量、活性磷酸盐、无机氮和硅酸盐呈显著负相关关系, 浮游植物群落结构的变化也预示着近年来长江口海域富营养水平加剧。
4 结论研究结果表明, 2004—2016年夏季共鉴定出浮游植物7门263种, 以硅藻占绝对优势, 甲藻次之, 物种数呈增加趋势, 其中硅藻种类数占有比例下降, 而甲藻的种类数占有比呈上升趋势。总细胞丰度范围、硅藻平均细胞丰度和甲藻平均细胞丰度都呈上升趋势, 除2014年外, 其余年份硅藻细胞丰度都占绝对优势并与总细胞丰度的分布曲线和变化趋势基本一致。水平分布模式及其年际变化受环流变化和水团消长的影响, 总体趋势是河口区细胞丰度较低, 由河口到外海呈逐渐增大趋势, 部分年份呈现先增大后减小的趋势。生物多样性多年呈下降趋势, 物种组成随空间和时间变化较大, 只有较少的种类能够保持稳定, 多数物种因调查时间和站位不同而只是偶尔被检出。中肋骨条藻有10年为区域第一优势种, 41种常见种中有31种细胞丰度呈增加趋势, 10种细胞丰度呈下降趋势。浮游植物细胞丰度与活性磷酸盐和无机氮具有相关性, 种类数、多样性指数除了与溶解氧、氨氮、叶绿素a无相关性外与其他10种环境因子都相关。本研究使用的是同一区域连续13年同一时间段的大面调查数据, 是研究浮游植物年际变化的珍贵数据资源, 能够较好反映长江口生态监控区浮游植物物种的本底情况及演变趋势。
致谢 感谢国家海洋局东海分局提供海洋环境监测数据。
参考文献
| 中华人民共和国国家质量监督检验检疫总局, 2007. GB 17378-2007海洋监测规范. 北京: 中国标准出版社, |
| 尹艳娥, 沈新强, 蒋玫, 等, 2014. 长江口及其邻近海域富营养化趋势分析及与环境因子关系. 生态环境学报, 23(4): 622–629 |
| 王丹, 孙军, 周锋, 等, 2008. 2006年6月长江口低氧区及邻近水域浮游植物. 海洋与湖沼, 39(6): 619–627 DOI:10.11693/hyhz200806012 |
| 王江涛, 曹婧, 2012. 长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 31(3): 310–315 |
| 王金辉, 2002. 长江口3个不同生态系的浮游植物群落. 青岛海洋大学学报, 32(3): 422–428 |
| 王金辉, 黄秀清, 刘阿成, 等, 2004. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 23(1): 32–39 |
| 刘海娇, 傅文诚, 孙军, 等, 2015. 2009-2011年东海陆架海域网采浮游植物群落的季节变化. 海洋学报, 37(10): 106–122 DOI:10.3969/j.issn.0253-4193.2015.10.011 |
| 孙军, 刘东艳, 2004. 多样性指数在海洋浮游植物研究中的应用. 海洋学报, 26(1): 62–75 |
| 江志兵, 曾江宁, 李宏亮, 等, 2014. 长江口及其邻近陆架区夏季网采浮游植物及其影响因素. 海洋学报, 36(6): 112–123 |
| 何青, 孙军, 2009. 长江口及其邻近水域网采浮游植物群落. 生态学报, 29(7): 3928–3938 |
| 张璇, 2012. 长江口及邻近海域营养盐的历史演变及其在赤潮中的作用研究. 青岛: 中国海洋大学硕士学位论文 |
| 张桂成, 2015. 长江口及其邻近海域溶解有机氮的生物可利用性及其在赤潮爆发过程中的作用研究. 青岛: 中国海洋大学博士学位论文, 2015 |
| 李云, 李道季, 唐静亮, 等, 2007. 长江口及毗邻海域浮游植物的分布与变化. 环境科学, 28(4): 719–729 |
| 辛明, 2014. 长江口海域关键环境因子的长期变化及其生态效应. 青岛: 中国海洋大学博士毕业论文 |
| 周名江, 颜天, 邹景忠, 2003. 长江口邻近海域赤潮发生区基本特征初探. 应用生态学报, 14(7): 1031–1038 |
| 国家海洋局, 2005. 2004年中国海洋环境质量公报. 北京: 国家海洋局 http://www.mlr.gov.cn/zwgk/tjxx/201004/t20100401_143693.htm |
| 国家海洋局, 2006. 2005年中国海洋环境质量公报. 北京: 国家海洋局 http://www.mlr.gov.cn/zwgk/tjxx/201004/t20100401_143690.htm |
| 国家海洋局, 2007. 2006年中国海洋环境质量公报. 北京: 国家海洋局 http://www.mlr.gov.cn/zwgk/tjxx/201004/t20100401_143687.htm |
| 国家海洋局, 2008. 2007年中国海洋环境质量公报. 北京: 国家海洋局. https://wenku.baidu.com/view/f20d77a1b0717fd5360cdcf1.html |
| 国家海洋局, 2009. 2008年中国海洋环境质量公报. 北京: 国家海洋局. http://www.mlr.gov.cn/zwgk/tjxx/201003/t20100325_712755.htm |
| 国家海洋局, 2010. 2009年中国海洋环境质量公报. 北京: 国家海洋局 https://wenku.baidu.com/view/e33323d376a20029bd642db6.html |
| 国家海洋局, 2011. 2010年中国海洋环境质量公报. 北京: 国家海洋局. https://wenku.baidu.com/view/c2c3d486ec3a87c24028c4d7.html |
| 国家海洋局, 2012. 2011年中国海洋环境质量公报. 北京: 国家海洋局 https://wenku.baidu.com/view/bf214630ee06eff9aef807eb.html |
| 国家海洋局, 2013. 2012年中国海洋环境质量公报. 北京: 国家海洋局 |
| 国家海洋局, 2014. 2013年中国海洋环境质量公报. 北京: 国家海洋局 |
| 国家海洋局, 2015. 2014年中国海洋环境质量公报. 北京: 国家海洋局 |
| 国家海洋局, 2016. 2015年中国海洋环境质量公报. 北京: 国家海洋局 |
| 国家海洋局, 2017. 2016年中国海洋环境质量公报. 北京: 国家海洋局 |
| 林梅, 李杨, 2017. 长江口海域春季浮游植物的年际变化. 海洋与湖沼, 48(2): 303–311 |
| 林峰竹, 吴玉霖, 于海成, 等, 2008. 2004年长江口浮游植物群落结构特征分析. 海洋与湖沼, 39(4): 401–410 |
| 范丽, 程金平, 郭广勇, 2008. 长江口污染物种类及及其来源. 江苏环境科技, 21(S1): 10–12, 15 |
| 赵冉, 孙军, 宋书群, 等, 2013. 2006年春季长江口及其邻近水域浮游植物. 海洋通报, 32(4): 421–428 DOI:10.11840/j.issn.1001-6392.2013.04.009 |
| 唐峰华, 伍玉梅, 樊伟, 等, 2010. 长江口浮游植物分布情况及与径流关系的初步探讨. 生态环境学报, 19(12): 2934–2940 DOI:10.3969/j.issn.1674-5906.2010.12.029 |
| 栾青杉, 孙军, 宋书群, 等, 2007. 长江口夏季浮游植物群落与环境因子的典范对应分析. 植物生态学报, 31(3): 445–450 |
| 栾青衫, 孙军, 2010. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系. 生态学报, 30(18): 4967–4975 |
| 郭皓, 2004. 中国近海赤潮生物图谱. 北京: 海洋出版社, |
| 郭术津, 2012. 东海浮游植物群集研究. 青岛: 中国海洋大学硕士学位论文 |
| 章飞燕, 2009. 长江口及邻近海域浮游植物群落变化的历史对比及其环境因子研究. 上海: 华东师范大学硕士毕业论文 |
| Gao X L, Song J M, 2005. Phytoplankton distributions and their relationship with the environment in the Changjiang Estuary, China. Marine Pollution Bulletin, 50(3): 327–335 DOI:10.1016/j.marpolbul.2004.11.004 |
| Jiang Z B, Liu J J, Chen J F, et al, 2014. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River)estuary during the past 50 years. Water Research, 54: 1–11 DOI:10.1016/j.watres.2014.01.032 |
| Jiang Z B, Chen J F, Zhou F, et al, 2015. Controlling factors of summer phytoplankton community in the Changjiang (Yangtze River) Estuary and adjacent East China Sea shelf. Continental Shelf Research, 101: 71–84 DOI:10.1016/j.csr.2015.04.009 |
| Kong F Z, Xu Z J, Yu R C, et al, 2016. Distribution patterns of phytoplankton in the Changjiang River estuary and adjacent waters in spring 2009. Chinese Journal of Oceanology and Limnology, 34(5): 902–914 DOI:10.1007/s00343-016-4202-6 |
| Yang S, Han X R, Zhang C S, et al, 2014. Seasonal changes in phytoplankton biomass and dominant species in the Changjiang River Estuary and adjacent seas:general trends based on field survey data 1959-2009. Journal of Ocean University of China, 13(6): 926–934 DOI:10.1007/s11802-014-2515-7 |
| Zhu Z Y, Ng W M, Liu S M, et al, 2009. Estuarine phytoplankton dynamics and shift of limiting factors:A study in the Changjiang (Yangtze River) Estuary and adjacent area. Estuarine, Coastal and Shelf Science, 84(3): 393–401 DOI:10.1016/j.ecss.2009.07.005 |