 2018, Vol. 49
2018, Vol. 49


大弹涂鱼(Boleophthalmus pectinirostris) piscidin 1的分子特征及功能分析

http://dx.doi.org/10.11693/hyhz20171100297
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 管峰, 李长红, 聂力, 苗亮, 陈炯. 2018.
- GUAN Feng, LI Chang-Hong, NIE Li, MIAO Liang, CHEN Jiong. 2018.
- 大弹涂鱼(Boleophthalmus pectinirostris) piscidin 1的分子特征及功能分析
- PISCIDIN 1 OF MUDSKIPPER BOLEOPHTHALMUS PECTINIROSTRIS: MOLECULAR CHARACTERIZATION AND FUNCTIONAL ANALYSIS
- 海洋与湖沼, 49(2): 422-431
- Oceanologia et Limnologia Sinica, 49(2): 422-431.
- http://dx.doi.org/10.11693/hyhz20171100297
-
文章历史
- 收稿日期:2017-11-26
- 收修改稿日期:2017-12-30
引用本文
管峰, 李长红, 聂力, 苗亮, 陈炯. 2018. 大弹涂鱼(Boleophthalmus pectinirostris) piscidin 1的分子特征及功能分析[J]. 海洋与湖沼, 49(2): 422-431
GUAN Feng, LI Chang-Hong, NIE Li, MIAO Liang, CHEN Jiong. 2018. PISCIDIN 1 OF MUDSKIPPER BOLEOPHTHALMUS PECTINIROSTRIS: MOLECULAR CHARACTERIZATION AND FUNCTIONAL ANALYSIS[J]. Oceanologia et Limnologia Sinica, 49(2): 422-431.
大弹涂鱼(Boleophthalmus pectinirostris) piscidin 1的分子特征及功能分析
1.
宁波大学海洋学院 宁波 315211;
2. 浙江万里学院生物与环境学院 宁波 315100
2. 浙江万里学院生物与环境学院 宁波 315100
摘要:摘要Piscidin类抗菌肽具有广谱的抗菌活性,在鱼类先天性免疫中扮演重要角色。本文对大弹涂鱼(Boleophthalmus pectinirostris)单核/巨噬细胞(monocytes/macrophages,MO/MΦ)转录组测序获得piscidin 1基因(Bppis1)cDNA全序列。Bppis1基因cDNA序列由327个核苷酸组成,开放阅读框为207bp,编码68个氨基酸,预测相对分子量为7.7kDa,等电点为5.51。氨基酸序列多重比对分析表明,Bppis1具有piscidin家族的特征结构,信号肽序列最为保守,终止于GEG序列后,与黄条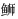 (Seriola lalandi)piscidin同源性最高,为40.8%;系统进化树分析表明,Bppis1属于piscidin 1类,与黄条
piscidin进化相关性最高。实时荧光定量PCR(quantitative real-time PCR,RT-qPCR)结果显示,Bppis1 mRNA在健康鱼鳃中表达量最高;迟缓爱德华氏菌(Edwardsiella tarda)感染后,大弹涂鱼肝、脾、肾、鳃和皮肤中Bppis1 mRNA表达量显著上调。体外抑菌实验结果表明,人工合成的Bppis1成熟肽抑菌活性较广泛,但对迟缓爱德华氏菌和3株革兰氏阳性菌无抑菌活性。大弹涂鱼MO/MΦ经1.0μg/mL Bppis1成熟肽处理后,对FITC标记的创伤弧菌(Vibrio vulnificus)的吞噬活性显著增加。综上,Bppis1在大弹涂鱼先天免疫中发挥重要作用,可能作为抵抗病原体入侵的潜在治疗药物。
(Seriola lalandi)piscidin同源性最高,为40.8%;系统进化树分析表明,Bppis1属于piscidin 1类,与黄条
piscidin进化相关性最高。实时荧光定量PCR(quantitative real-time PCR,RT-qPCR)结果显示,Bppis1 mRNA在健康鱼鳃中表达量最高;迟缓爱德华氏菌(Edwardsiella tarda)感染后,大弹涂鱼肝、脾、肾、鳃和皮肤中Bppis1 mRNA表达量显著上调。体外抑菌实验结果表明,人工合成的Bppis1成熟肽抑菌活性较广泛,但对迟缓爱德华氏菌和3株革兰氏阳性菌无抑菌活性。大弹涂鱼MO/MΦ经1.0μg/mL Bppis1成熟肽处理后,对FITC标记的创伤弧菌(Vibrio vulnificus)的吞噬活性显著增加。综上,Bppis1在大弹涂鱼先天免疫中发挥重要作用,可能作为抵抗病原体入侵的潜在治疗药物。
(Seriola lalandi)piscidin同源性最高,为40.8%;系统进化树分析表明,Bppis1属于piscidin 1类,与黄条
piscidin进化相关性最高。实时荧光定量PCR(quantitative real-time PCR,RT-qPCR)结果显示,Bppis1 mRNA在健康鱼鳃中表达量最高;迟缓爱德华氏菌(Edwardsiella tarda)感染后,大弹涂鱼肝、脾、肾、鳃和皮肤中Bppis1 mRNA表达量显著上调。体外抑菌实验结果表明,人工合成的Bppis1成熟肽抑菌活性较广泛,但对迟缓爱德华氏菌和3株革兰氏阳性菌无抑菌活性。大弹涂鱼MO/MΦ经1.0μg/mL Bppis1成熟肽处理后,对FITC标记的创伤弧菌(Vibrio vulnificus)的吞噬活性显著增加。综上,Bppis1在大弹涂鱼先天免疫中发挥重要作用,可能作为抵抗病原体入侵的潜在治疗药物。
关键词:大弹涂鱼 Piscidin 迟缓爱德华氏菌 抗菌活性 吞噬活性
PISCIDIN 1 OF MUDSKIPPER BOLEOPHTHALMUS PECTINIROSTRIS: MOLECULAR CHARACTERIZATION AND FUNCTIONAL ANALYSIS
1.
School of Marine Sciences, Ningbo University, Ningbo 315211, China;
2. Zhejiang Wanli University, Ningbo 315100, China
2. Zhejiang Wanli University, Ningbo 315100, China
Abstract:
Piscidin-like antimicrobial peptides, which have broad-spectrum activities, exert an important role in fish's innate immunity. In this study, we obtained the complete cDNA sequence of piscidin 1 (Bppis1) from mudskipper Boleophthalmus pectinirostris, with de novo transcriptome sequencing of mudskipper monocytes/macrophages (MO/MΦ). The full-length of cDNA sequence of Bppis1 was 327 nucleotides, containing a 207 nucleotides open reading frame (ORF) encoding 68 amino acids. The deduced molecular weight and isoelectric point of the deduced protein was 7.7kDa and 5.51 respectively. The multiple sequence alignment of complete amino acid sequences revealed that Bppis1 possessed the piscidin family signature motifs, and a conserved sequence in signal peptide terminating at the motif GEG. Bppis1 shared the highest amino acid sequence identity (40.8%) with that of yellowtail amberjack (Seriola lalandi). In addition, the phylogenetic tree confirmed that Bppis1 belonged to fish piscidin 1 cluster and was most closely related to yellowtail amberjack piscidin. Quantitative real-time PCR (RT-qPCR) analysis showed that Bppis1 mRNA had the highest expression level in gill of healthy mudskipper. Upon infection with Edwardsiella tarda, Bppis1 mRNA expression increased significantly in the liver, spleen, kidney, gill and skin. The result of antimicrobial test showed that the synthetic mature peptide of Bppis1 exhibited relatively broad antimicrobial activity in vitro, but no antimicrobial activity against Edwardsiella tarda and three gram-positive bacteria. After treatment with 1.0μg/mL Bppis1 mature peptide, the phagocytosis of FITC-Vibrio vulnificus by MO/MФ was enhanced significantly. Therefore, Bppis1 plays a critical role in the innate immune system of mudskipper, and may be a potential therapeutic agent against pathogen invasion.
Key words:
Boleophthalmus pectinirostris
piscidin
Edwardsiella tarda
antimicrobial activity
phagocytic activity
抗菌肽(antimicrobial peptides, AMPs)是动物先天免疫系统的重要组分之一, 在机体抵抗病原入侵时发挥重要作用。AMPs多是小分子量的短肽, 抗菌谱广, 主要通过形成离子通道直接破坏细胞膜以杀灭病原菌, 病原菌很难对其产生抗性(Brogden, 2005; Lai et al, 2009)。研究表明, AMPs除对细菌、真菌、病毒、原生动物和癌细胞等有良好的直接抑制或杀伤作用外, 还参与免疫激活、免疫调节、抗炎和免疫抑制等免疫过程(Lai et al, 2009; Choi et al, 2012; Hilchie et al, 2013; Wan et al, 2014; Hancock et al, 2016)。近年来, 国内外学者已从水产动物中分离鉴定了豹鳎毒素(pardaxin) (Lazarovici et al, 1986)、毒鱼豆素(piscidin) (Silphaduang et al, 2001)、cathelicidin (Lu et al, 2011)、肝脏表达抗菌肽-1 (liver express antimicrobial peptide 1, LEAP-1/hepcidin) (陈梅珍等, 2010; 王克坚, 2011)、肝脏表达抗菌肽-2 (liver express antimicrobial peptide 2, LEAP-2) (Li et al, 2014)、防御素(defensin) (Zhu et al, 2017)、epinecidin-1 (Pan et al, 2007)和scygonadin (王克坚, 2011)等多种AMPs。
Piscidin是一类广泛存在于鱼类中的AMP之一, 最早从杂交斑纹鲈鱼(Morone chrysops × Morone saxatilis)的肥大细胞中分离(Silphaduang et al, 2001)。此后, 在鳜鱼(Siniperca chuatsi) (Sun et al, 2007)、大西洋鳕(Gadus morhua L.) (Fernandes et al, 2010; Ruangsri et al, 2012)、南极独角雪冰鱼(Chionodraco hamatus) (Buonocore et al, 2012)、罗非鱼(Oreochromis niloticus) (Peng et al, 2012)、大黄鱼(Larimichthys crocea) (Niu et al, 2013; Zhou et al, 2014; Yang et al, 2016)、条石鲷(Oplegnathus fasciatus) (Umasuthan et al, 2016)和斜带石斑鱼(Epinephelus coioides) (Zhuang et al, 2017)等数种鱼类中均发现它的存在。已有的研究发现, piscidin通过破坏细菌、真菌和寄生虫的细胞膜发挥直接的抑制活性(Campagna et al, 2007; Sung et al, 2008; Hayden et al, 2015; Li et al, 2016; Umasuthan et al, 2016; Pan et al, 2017)。除具有直接的抗病原活性外, piscidin还能通过免疫调节功能参与机体内病原诱导的免疫应答(Huang et al, 2015; Lin et al, 2016; Pan et al, 2017)。由于与piscidin具有相似的初级或二级多肽结构以及基因结构, 从其他鱼类中鉴定的拟鲽毒素(pleurocidin) (Cole et al, 1997)、moronecidin (Lauth et al, 2002; Bae et al, 2014)、chrysophsin(Iijima et al, 2003)、dicentracin(Salerno et al, 2007?)、epinecidin (Yin et al, 2006; Pan et al, 2007)、myxinidin (Subramanian et al, 2009)和gaduscidin (Browne et al, 2011)等抗菌肽被建议归为piscidin家族。
大弹涂鱼(Boleophthalmus pectinirostris)是弹涂鱼的一种, 主要分布在中国、韩国和日本等地, 在我国主要盛产于东南沿海和台湾。它是一种能够适应不同盐度的两栖鱼类, 喜栖息于港湾和河口潮间带淤泥滩涂上, 摄食有机碎屑、底藻和小型底栖动物等, 营掘洞穴居生活, 具有较高的食用与药用价值。尽管大弹涂鱼生活的滩涂环境含有种类繁多、数量巨大的微生物群落, 但它们很少感染严重的细菌性疾病, 因此研究大弹涂鱼抗感染应答的免疫机制显得尤为重要。鉴于piscidin在动物抗感染免疫应答中的重要作用, 本研究初步研究大弹涂鱼(B. pectinirostris)的piscidin 1(Bppis1), 测得Bppis1基因的cDNA序列, 确定该基因结构特点、分析其氨基酸系统进化关系并确定mRNA在正常组织中表达情况, 研究迟缓爱德华氏菌(Edwardsiella tarda)感染后其组织中Bppis1 mRNA的表达变化; 人工合成Bppis1成熟肽, 分析其体外抑菌活性以及对大弹涂鱼单核/巨噬细胞(monocytes/ macrophages, MO/MΦ)功能的影响, 为进一步深入探讨piscidin对鱼类抵抗细菌感染的免疫作用提供依据。
1 材料与方法 1.1 实验材料与实验试剂30—40g健康大弹涂鱼(Boleophthalmus pectinirostris)购自浙江省宁波市路林市场, 规格均一、无病症。大弹涂鱼充气运回实验室, 在充分曝气、盐度为10‰、温度为24—26℃水环境中暂养以适应实验室环境。适应期间连续充气, 早晚换水各一次。
迟缓爱德华氏菌(Edwardsiella tarda)Et-CD由浙江省海水养殖研究所惠赠, 其他细菌菌株由本实验保存。RNAiso试剂、AMV逆转录酶、SYBR Premix Ex Taq试剂盒等购自TaKaRa公司(中国)。Ficoll购自Invitrogen公司(中国)。RPMI 1640培养基和胎牛血清购自Gibco公司(美国)。Bppis1成熟肽由吉尔生化(上海)有限公司合成, 纯度 > 95%。引物合成和序列测定由生工生物工程(上海)股份有限公司完成。
1.2 大弹涂鱼肾源MO/MΦ的分离培养大弹涂鱼肾源MO/MΦ的分离培养过程参考Chen等(2016)的方法, 具体步骤如下:用0.1% (V/V)的间氨基苯甲酸乙酯甲磺酸盐(MS-222)麻醉大弹涂鱼, 在无菌环境中抽血并迅速取出体肾, 放入无菌过滤筛网中, 再用1mL RPMI 1640培养基润湿体肾, 注射器活塞按压体肾碎片, 收集分离细胞。采用Ficoll密度梯度离心方法继续分离细胞, 获得含有MO/MΦ的白色细胞混悬层, 收集混悬细胞后用含2%胎牛血清的RPMI 1640培养基洗涤2次, 离心, 重复2次, 再用同样浓度胎牛血清的RPMI 1640培养基重新混悬细胞。使用血球计数板对分离细胞进行计数, 将MO/MΦ的浓度调至2.0×107 cells/mL, 每个35mm细胞培养皿加入2mL细胞, 置于5% CO2、24℃细胞培养箱中培养24h, 然后用RPMI 1640培养基除去未贴壁细胞, 重复2次, 用含10%胎牛血清的RPMI 1640培养基继续培养贴壁细胞。用Giemsa′s染液对贴壁细胞染色并镜检细胞的形态特征, 最终确认细胞培养皿中超过95%贴壁细胞属于MO/MΦ。
1.3 Bppis1 cDNA序列获得及分析应用Illumina二代测序平台构建正常无病症大弹涂鱼的MO/MΦ转录组数据库, 筛选取得Bppis1 cDNA序列。根据说明书, 从大弹涂鱼MO/MΦ中提取总RNA, 经AMV逆转录酶合成互补cDNA链, 通过特异性引物对PCR进行扩增并将扩增产物测序检验。使用SignalP 4.0软件(http://www.cbs.dtu.dk/services/SignalP-4.0/)对开放阅读框编码多肽的信号肽序列进行预测; 采用pI/Mw程序(http://web.expasy.org/compute_pi/)对基因编码多肽的分子量和等电点进行预测; 应用ClustalW程序(http://clustalw.ddbj.nig.ac.jp/)多重比对piscidin氨基酸序列; 使用MEGA 5.0软件构建piscidin氨基酸的系统进化树(Tamura et al, 2011)。构建系统发育进化树及比对多重序列所用piscidin的氨基酸序列详见表 1。
表 1 Piscidin氨基酸序列多重比对及系统发育进化树构建采用序列 Tab. 1 Protein sequences of piscidin for phylogenetic tree construction and multiple alignment
| NCBI登录号 | 基因名称 | 物种 | ||
| 拉丁名 | 英文名 | 中文名 | ||
| MF459711 | piscidin 1 | Boleophthalmus pectinirostris | Mudskipper | 大弹涂鱼 |
| AB703274 | piscidin | Oplegnathus fasciatus | Rock bream | 条石鲷 |
| EU741827 | piscidin l | Larimichthys crocea | Large yellow croaker | 大黄鱼 |
| JX412481 | piscidin 1 | Epinephelus malabaricus | Malabar grouper | 玛拉巴石斑鱼 |
| JQ823163 | piscidin l | Epinephelus coioides | Orange-spotted grouper | 斜带石斑鱼 |
| EU741828 | piscidin l | Epinephelus akaara | Red spotted grouper | 赤点石斑鱼 |
| GU592793 | piscidin l | Epinephelus fuscoguttatus | Brown marbled grouper | 褐点石斑鱼 |
| HQ437912 | piscidin l | Epinephelus bleekeri | Duskytail grouper | 布氏石斑鱼 |
| JN216987 | piscidin l | Epinephelus bruneus | Longtooth grouper | 褐石斑鱼 |
| FJ917596 | piscidin 1 | Gadus morhua L. | Atlantic cod | 大西洋鳕 |
| KY548530 | piscidin | Seriola lalandi | Yellowtail amberjack | 黄条
|
| HQ437913 | piscidin 2 | Epinephelus coioides | Orange-spotted grouper | 斜带石斑鱼 |
| JX412480 | piscidin 2 | Epinephelus malabaricus | Malabar grouper | 玛拉巴石斑鱼 |
| HQ184323 | piscidin 2β | Gadus morhua L. | Atlantic cod | 大西洋鳕 |
| HQ184322 | piscidin 2 | Gadus morhua L. | Atlantic cod | 大西洋鳕 |
| KX231319 | piscidin 3 | Morone chrysops | White bass | 白鲈 |
| KX231323 | piscidin 3 | Morone saxatilis | Striped bass | 条纹鲈 |
| KX231324 | piscidin 4 | Morone chrysops | White bass | 白鲈 |
| KX231320 | piscidin 4 | Morone saxatilis | Striped bass | 条纹鲈 |
| HM596029 | piscidin 4 | Morone chrysops ×Morone saxatilis | Hybrid striped bass | 杂交斑纹鲈鱼 |
| HM596030 | piscidin 5 | Morone chrysops ×Morone saxatilis | Hybrid striped bass | 杂交斑纹鲈鱼 |
| KX870851 | piscidin 5lt3 | Larimichthys crocea | Large yellow croaker | 大黄鱼 |
| KJ879923 | piscidin 5lt2 | Larimichthys crocea | Large yellow croaker | 大黄鱼 |
| KJ879922 | piscidin 5l | Larimichthys crocea | Large yellow croaker | 大黄鱼 |
| KX231326 | piscidin 6 | Morone chrysops | White bass | 白鲈 |
| KX231321 | piscidin 6 | Morone saxatilis | Striped bass | 条纹鲈 |
| KX231322 | piscidin 7 | Morone chrysops | White bass | 白鲈 |
腹腔注射迟缓爱德华氏菌感染大弹涂鱼的实验方法及感染剂量参考Chen等(2016), 具体步骤如下:收集对数生长期的迟缓爱德华氏菌, 用灭菌生理盐水洗涤3次, 将细菌浓度调整为1.0×105 CFU/mL。细菌感染组大弹涂鱼腹腔注射1.0×104 CFU/尾的迟缓爱德华氏菌, 对照组大弹涂鱼腹腔注入同等剂量的生理盐水, 在注射感染后4、8、12和24h (hours post infection, hpi)时分别收集两个组实验鱼的肝、脾、鳃、肾和皮肤等组织, 并立即放入液氮中冻存, 随后置-70℃超低温冰箱保存。
1.5 实时荧光定量PCR(quantitative real-time PCR, RT-qPCR)参考Chen等(2016)的方法抽提组织中总RNA、合成互补cDNA链并进行RT-qPCR检测目的基因mRNA的表达量。根据转录组测序结果获得的Bppis1序列设计检测引物, Bppis1-T(+): 5′-TGGTGGTGATG ATGGCTGAG-3′和Bppis1-T(-): 5′-CACTTGGAGGTA TTCG GGGC-3′, 预期扩增片段大小为172bp。根据大弹涂鱼18S rRNA基因设计检测引物, Bp18S rRNA-T(+): 5′-GGCCGTTCTTAGTTGGTGGA-3′和Bp18S rRNA-T(-): 5′-CCCGGACATCTAAGGGCAT C-3′, 预期扩增片段大小为112bp。确定的RT-qPCR检测体系共25μL, 包含上下游引物(10μmol/L)各1μL、cDNA模板0.5μL、SYBR Premix Ex Taq(2×)缓冲液12.5μL、灭菌水10μL。在ABI荧光定量PCR仪(Invitrogen, Foster City, CA, USA)上运行检测程序, 程序如下:预变性段共1个循环反应, 95℃ 5min; 扩增段共40个循环, 95℃ 30s, 60℃ 30s, 72℃ 30s;熔解段共1个循环反应, 94℃ 30s, 72℃ 60s, 95℃ 30s。每个样品反复检测4次。根据2–ΔΔCt方法计算Bppis1 mRNA的相对表达量(Livak et al, 2001)。
1.6 体外抑菌实验参考Li等(2014)的方法, 采用微量稀释法测定合成Bppis1肽对不同细菌的最低抑菌浓度(minimum inhibitory concentration, MIC), 略作改进。具体方法如下:采用二倍稀释法将合成Bppis1肽配制成1.563、3.125、6.25、12.5、25、50和100μg/mL共7个浓度; 将副溶血弧菌(Vibrio parahaemolyticus)、金黄色葡萄球菌(Staphylococcus aureus)、溶藻弧菌(Vibrio alginolyticus)、大肠杆菌(Escherichia coli)、创伤弧菌(Vibrio vulnificus)、哈维氏弧菌(Vibrio harveyi)、嗜水气单胞菌(Aeromonas hydrophila)、鳗弧菌(Vibrio anguillarum)、迟缓爱德华氏菌、单核细胞增生李斯特菌(Listeria monocytogenes)和海豚链球菌(Streptococcus iniae)等11种细菌培养至对数生长期, 用无菌生理盐水分别调整这些细菌的浓度为1.0×105 CFU/mL; 在96孔微孔板中每孔加入20μL 1.0×105 CFU/mL实验细菌的菌悬液, 然后加入80μL不同浓度的合成Bppis1肽, 混匀后在适当的温度条件下培养24h。采用Varioskan Flash全波长多功能酶标仪(Thermo Fisher Scientific, Waltham, MA, USA)测定OD600值。通过细菌沉降和600nm处吸光度值计算MIC, 同时设置卡那霉素为阳性对照。
1.7 吞噬实验参考Chen等(2016)的方法测定Bppis1对大弹涂鱼MO/MΦ吞噬能力的影响, 方法如下:分别将对数生长期的大肠杆菌和创伤弧菌标记异硫氰酸荧光素(fluorescein isothiocyanate, FITC), 分别命名为FITC-E. coli和FITC-V. vulnificus。将1.0或10.0μg/mL Bppis1肽加入到MO/MΦ中, 预处理8h, 然后以感染复数(multiplicity of infection, MOI)为10︰1的比例加入FITC-E. coli或FITC-V. vulnificus, 孵育30min, 灭菌PBS洗涤3次以除去未吞噬细菌, 台盼蓝淬灭细胞外荧光, 最后用FACS缓冲液(含0.2% BSA和0.1%叠氮化钠)重悬细胞。使用Gallios Flow Cytometer (Beckman Coulterr, Fullerton, CA, USA)对被吞噬细菌的荧光信号进行流式检测。大弹涂鱼MO/MΦ吞噬细菌的相对平均荧光强度(relative fluorescence intensity, MFI)用FlowJo软件分析。
2 结果 2.1 Bppis1 cDNA序列分析软件分析结果显示, Bppis1 cDNA全长327bp (GenBank登录号为MF459711), 包含一个完整的、长度为207bp的开放阅读框, 预测编码一个由68个氨基酸组成的、相对分子量大小为7.7kDa的多肽, 等电点为5.51。该多肽N-末端含一个由22个氨基酸组成的信号肽, 裂解位点为G20E21G22-F23F24, C-末端含一个由28个氨基酸组成的末端功能前区, 其成熟肽由18个氨基酸组成(图 1)。
氨基酸序列多重比对结果显示, 鱼类piscidin在信号肽区间序列最为保守, 终止于1个GEG或GEC序列后, 成熟肽和末端功能前区序列变异较大(图 1)。其中, 大弹涂鱼piscidin与其他已知物种piscidin序列之间同源性较低, 为5.8%—40.8%, 与黄条
取健康大弹涂鱼的肝、脾、肾、鳃和皮肤运行RT-qPCR检验程序, 结果显示Bppis1 mRNA在健康大弹涂鱼鳃组织中含量最高, 其余依次为皮肤、肝、脾和肾(图 3A)。与对照组对比, 当迟缓爱德华氏菌感染4h时, 脾、肾、肝和皮肤中Bppis1 mRNA的表达量显著提高, 分别是对照组表达量的2.17、3.17、3.32和20.76倍(P < 0.05);在8h时, 所有检测组织中Bppis1 mRNA的表达量均显著增加, 在肝、肾、鳃和皮肤中达到峰值, 分别为对照组的5.84、15.51、3.78和60.86倍(P < 0.05);在12h时, 脾中Bppis1 mRNA的表达量继续增加, 达到峰值, 为对照组的20.60倍(P < 0.05);在24h时, 肾、肝和鳃中Bppis1 mRNA的表达量与对照组无明显差异(图 3B—图 3F)。
 |
| 图 3 RT-qPCR分析Bppis1 mRNA的表达 Fig. 3 RT-qPCR analysis of Bppis1 mRNA 注:A. Bppis1 mRNA在健康大弹涂鱼中的表达; B—F: Bppis1 mRNA在迟缓爱德华氏菌感染大弹涂鱼组织中的表达。以Bppis1 mRNA与18S rRNA基因的比值作为Bppis1的相对表达量, *表示与对照组比较差异显著(P < 0.05) (n=4) |
利用酶标仪测定Bppis1肽对11种细菌的体外抑菌活性, 结果表明, Bppis1具备较广泛的抗菌活性, 对溶藻弧菌、副溶血弧菌和鳗弧菌的MIC为50μg/mL; 对哈维氏弧菌和嗜水气单胞菌的MIC为25μg/mL; 对创伤弧菌和大肠杆菌的MIC分别为12.5和6.25μg/mL, 并且抑菌效果优于对照组卡那霉素, 但对迟缓爱德华氏菌、金黄色葡萄球菌、单核细胞增生李斯特菌和海豚链球菌无明显的抑菌活性(表 2)。
表 2 Bppis1的体外抑菌MIC Tab. 2 MIC values of synthetic Bppis1 against bacteria in vitro
| 细菌名 | 分离株/菌株 | 培养基 | 培养温度(℃) | Bppis1 MIC (μg/mL) | 卡那霉素MIC (μg/mL) |
| 创伤弧菌 | ATCC279562 | TSB | 28 | 12.5 | 25 |
| 溶藻弧菌 | ATCC17749 | TSB | 28 | 50 | 25 |
| 副溶血弧菌 | ATCC33847 | TSB | 28 | 50 | 50 |
| 哈维氏弧菌 | ATCC33866 | TSB | 28 | 25 | 1.563 |
| 鳗弧菌 | ATCC19264 | TSB | 28 | 50 | 25 |
| 迟缓爱德华氏菌 | Et-CD | LB | 37 | — | 25 |
| 大肠杆菌 | K12 | LB | 37 | 6.25 | 50 |
| 金黄色葡萄球菌 | ATCC6538 | LB | 37 | — | 12.5 |
| 嗜水气单胞菌 | ATCC7966 | LB | 37 | 25 | 6.25 |
| 单核细胞增生李斯特菌 | ATCC19115 | BHI | 37 | — | 1.563 |
| 海豚链球菌 | ATCC29178 | BHI | 28 | — | 6.25 |
| 注:“—”表示Bppis1在100μg/mL时无抑菌活性 | |||||
采用Gallios Flow Cytometer检测合成Bppis1肽对大弹涂鱼MO/MΦ吞噬能力的影响。结果显示, 与对照组相比, 经1.0μg/mL的合成Bppis1肽处理后, MO/MΦ对FITC-E. coli吞噬活性无明显变化, 但对FITC-V. vulnificus吞噬活性显著增加, 为对照组的1.51倍; 经10.0μg/mL的合成Bppis1肽处理后, MO/MΦ对FITC-E. coli和FITC-V. vulnificus的吞噬活性均无明显变化(图 4)。
 |
| 图 4 合成Bppis1肽对大弹涂鱼MO/MΦ吞噬活性的影响 Fig. 4 Effect of synthetic Bppis1 on phagocytosis activity of mudskipper MO/MΦ 注:A和C:吞噬流式图; B和D:相对平均荧光强度。PBS处理组的相对平均荧光强度数值设定为100, 实验组的相对平均荧光强度用PBS处理组数值的倍数表示, *表示与对照组比较差异显著(P < 0.05) (n=3) |
Piscidin作为抗菌肽piscidin家族成员之一, 是鱼类先天免疫的重要组成部分。本研究从大弹涂鱼MO/MΦ中鉴定了Bppis1基因cDNA序列。序列分析结果表明, Bppis1具有与其他物种piscidin相似的结构特征, 由信号肽、成熟肽和末端功能前区组成, 其中信号肽序列最为保守, 以GEG序列终止, 成熟肽和末端功能前区序列变异较大。Bppis1与其他已知物种piscidin序列之间同源性较低, 与黄条
组织特征分析表明, Bppis1 mRNA在健康大弹涂鱼各组织中广泛表达, 且在鳃中表达量最高, 与已报道的南极独角雪冰鱼(Buonocore et al, 2012)、条石鲷(Bae et al, 2016; Umasuthan et al, 2016)、罗非鱼(Peng et al, 2012)、大黄鱼(Niu et al, 2013)等鱼类研究结果较为一致。鳃是鱼类抵抗病原入侵的重要屏障之一, Bppis1在鳃中表达量丰富, 揭示其在大弹涂鱼天然免疫应答中扮演重要角色。多篇文献表明鱼类piscidin的高表达与感染病原体紧密相联, 研究结果显示病原感染后piscidin的表达显著上调(Sun et al, 2007; Niu et al, 2013; Umasuthan et al, 2016; Zhuang et al, 2017)。例如, 斜带石斑鱼感染副溶血弧菌后, 脾中ecPis-1 mRNA表达在3h时显著上调(P < 0.05), 体肾中ecPis-2和ecPis-3 mRNA表达均在12h显著上调, 脾和头肾中ecPis-2和ecPis-4 mRNA在12h和24h时均显著上调(Zhuang et al, 2017);条石鲷感染条石鲷虹彩病毒(rock bream iridovirus, RBIV)、迟缓爱德华氏菌和海豚链球菌后, 鳃和头肾中Of-Pis1 mRNA表达均在3h时显著上调, 而且RBIV感染使得头肾中Of-Pis1 mRNA表达在12h达到峰值, 约为对照组的12倍(Umasuthan et al, 2016)。本研究中, 大弹涂鱼感染迟缓爱德华氏菌后, 肾、脾、肝、皮肤和鳃中Bppis1 mRNA表达结果与上述论文中的结论相似, 只是峰值数据和到达峰值的时间点略有差别, 肾、肝、皮肤和鳃中Bppis1 mRNA表达均在8h时达到峰值, 分别为对照组的15.51、5.84、60.86和3.78倍(P < 0.05), 脾中Bppis1 mRNA表达在12h达到峰值, 为对照组的20.6倍(P < 0.05), 揭示Bppis1可能在大弹涂鱼抗迟缓爱德华氏菌感染过程中发挥重要作用。
研究发现, piscidin对病毒、细菌、真菌和寄生虫均有抑制作用, 主要通过破坏细菌、真菌和寄生虫的细胞膜发挥直接的抑制活性(Chinchar et al, 2004; Campagna et al, 2007; Sung et al, 2008; Hayden et al, 2015; Li et al, 2016; Umasuthan et al, 2016; Pan et al, 2017)。体外抑菌实验结果表明, 人工合成或重组表达的piscidin成熟肽具有广泛的抗细菌活性, 对革兰氏阳性菌和阴性菌均有抑制作用(Bae et al, 2016; Umasuthan et al, 2016)。例如, 在条石鲷中, 重组表达的piscidin (pOf-Pis)对Vibrio tapetis、大肠杆菌和海豚链球菌均有较强抑菌活性, MIC分别为22.5、45和45µg/mL, 对迟缓爱德华氏菌抑菌活性较弱, MIC大于90µg/mL, 并且不能完全抑制迟缓爱德华氏菌的生长(Umasuthan et al, 2016);人工合成的piscidin (RbPisc)对海豚链球菌、哈维氏弧菌和奥氏弧菌(V. ordalii)的抑菌活性非常强, MIC均低于0.9µg/mL, 对大肠杆菌、溶藻弧菌、坎氏弧菌(V. campbellii)和创伤弧菌抗菌活性较强, MIC分别为1.9—3.9、0.9—1.9、1.9和3.9—7.8µg/mL, 对迟缓爱德华氏菌几乎没有抑制活性, MIC为250—500µg/mL (Bae et al, 2016)。在罗非鱼中, 人工合成的piscidin 3 (TP3)和piscidin 4 (TP4)对创伤弧菌、嗜水气单胞菌、溶藻弧菌、铜绿假单胞菌(Pseudomona aeruginosa)、无乳链球菌(Streptococcus galactiae)和粪肠球菌(Enterococcus faecalis)均有较强的抑菌活性, 而TP1、TP2和TP5没有明显的抑菌活性(Peng et al, 2012)。在大黄鱼中, 人工合成的piscidin (Lcpis5lt3)对单核细胞增生李斯特菌、金黄色葡萄球菌、鳗弧菌、溶藻弧菌、副溶血弧菌和嗜水气单胞菌具有明显的抗菌效果, 对迟缓爱德华氏菌、创伤弧菌、哈维氏弧菌和海豚链球菌无明显抑制作用(Yang et al, 2016)。本研究中, 人工合成的Bppis1成熟肽对大肠杆菌和弧菌科6种细菌有明显的抗菌活性, MIC为6.25—50μg/mL, 但对迟缓爱德华氏菌以及3株革兰氏阳性菌无抗菌活性, 与上述研究结果有明显差异, 说明不同鱼类的piscidin对相同细菌的抗菌活性不同, Bppis1可能仅具有抗革兰氏阴性菌活性。
新近研究表明, 罗非鱼TP3和TP4能通过影响免疫相关基因, 如转移生长因子(transforming growth factor β, TGF-β)和细胞因子类白细胞介素(interleukin, IL)-1β、IL-6、IL-8等mRNA的表达来发挥免疫调节功能(Lin et al, 2016; Pan et al, 2017)。此外, Huang等(2015)研究发现, 2.5—10µg/mL TP4不仅能刺激纤维原细胞系(Hs-68)和角化细胞系(HaCaT)的增殖, 也能影响TGF、血管上皮生长因子和表皮生长因子调节的伤口闭合活性, 从而增加耐甲氧西林金黄色葡萄球菌(methicillin-resistant Staphylococcus aureus, MRSA)感染大鼠的存活率。本研究发现, 大弹涂鱼MO/MΦ经1.0μg/mL的Bppis1处理后, 对创伤弧菌的吞噬作用显著增强, 揭示Bppis1也具有免疫调节功能, 可能通过促进免疫细胞的吞噬活性发挥作用, 但具体作用机制尚待进一步研究。
4 结论本研究测定了大弹涂鱼Bppis1基因cDNA序列, 序列分析揭示其与黄条
参考文献
| 王克坚., 2011. 海洋鱼类和青蟹抗菌肽hepcidin和scygonadin的研究. 厦门大学学报(自然科学版), 50(2): 418–424 |
| 陈梅珍, 陈炯, 陆新江, 等, 2010. 香鱼hepcidin基因的克隆、序列分析及组织表达特征. 动物学研究, 31(6): 595–600 |
| Bae J S, Jung J M, An C M, et al, 2016. Piscidin:Antimicrobial peptide of rock bream, Oplegnathus fasciatus. Fish & Shellfish Immunology, 51: 136–142 |
| Bae J S, Shim H S, Hwang S D, et al, 2014. Expression analysis and biological activity of moronecidin from rock bream, Oplegnathus fasciatus. Fish & Shellfish Immunology, 40(2): 345–353 |
| Brogden K A, 2005. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nature Reviews Microbiology, 3(3): 238-250 |
| Browne M J, Feng C Y, Booth V, et al, 2011. Characterization and expression studies of Gaduscidin-1 and Gaduscidin-2; paralogous antimicrobial peptide-like transcripts from Atlantic cod (Gadus morhua). Developmental & Comparative Immunology, 35(3): 399–408 |
| Buonocore F, Randelli E, Casani D, et al, 2012. A piscidin-like antimicrobial peptide from the icefish Chionodraco hamatus (Perciformes:Channichthyidae):molecular characterization, localization and bactericidal activity. Fish & Shellfish Immunology, 33(5): 1183–1191 |
| Campagna S, Saint N, Molle G, et al, 2007. Structure and mechanism of action of the antimicrobial peptide piscidin. Biochemistry, 46(7): 1771–1778 DOI:10.1021/bi0620297 |
| Chen J, Chen Q, Lu X J, et al, 2016. The protection effect of LEAP-2 on the mudskipper (Boleophthalmus pectinirostris) against Edwardsiella tarda infection is associated with its immunomodulatory activity on monocytes/macrophages. Fish & Shellfish Immunology, 59: 66–76 |
| Chinchar V G, Bryan L, Silphadaung U, et al, 2004. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology, 323(2): 268–275 DOI:10.1016/j.virol.2004.02.029 |
| Choi K Y, Chow L N, Mookherjee N., 2012. Cationic host defence peptides:multifaceted role in immune modulation and inflammation. Journal of Innate Immunity, 4(4): 361–370 |
| Cole A M, Weis P, Diamond G., 1997. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. Journal of Biological Chemistry, 272(18): 12008–12013 DOI:10.1074/jbc.272.18.12008 |
| Fernandes J M, Ruangsri J, Kiron V., 2010. Atlantic cod piscidin and its diversification through positive selection. PLoS One, 5(3): e9501 |
| Hancock R E, Haney E F, Gill E E., 2016. The immunology of host defence peptides:beyond antimicrobial activity. Nature Reviews Immunology, 16(5): 321–334 DOI:10.1038/nri.2016.29 |
| Hayden R M, Goldberg G K, Ferguson B M, et al, 2015. Complementary effects of host defense peptides piscidin 1 and piscidin 3 on DNA and lipid membranes:biophysical insights into contrasting biological activities. The Journal of Physical Chemistry B, 119(49): 15235–15246 DOI:10.1021/acs.jpcb.5b09685 |
| Hilchie A L, Wuerth K, Hancock R E., 2013. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nature Chemical Biology, 9(12): 761–768 DOI:10.1038/nchembio.1393 |
| Huang H N, Chan Y L, Wu C J, et al, 2015. Tilapia piscidin 4 (TP4) stimulates cell proliferation and wound closure in MRSA-infected wounds in mice. Marine Drugs, 13(5): 2813–2833 DOI:10.3390/md13052813 |
| Iijima N, Tanimoto N, Emoto Y, et al, 2003. Purification and characterization of three isoforms of chrysophsin, a novel antimicrobial peptide in the gills of the red sea bream, Chrysophrys major. European Journal of Biochemistry, 270(4): 675–686 DOI:10.1046/j.1432-1033.2003.03419.x |
| Lai Y, Gallo R L., 2009. AMPed up immunity:how antimicrobial peptides have multiple roles in immune defense. Trends in Immunology, 30(3): 131–141 DOI:10.1016/j.it.2008.12.003 |
| Lauth X, Shike H, Burns J C, et al, 2002. Discovery and characterization of two isoforms of moronecidin, a novel antimicrobial peptide from hybrid striped bass. Journal of Biological Chemistry, 277(7): 5030–5039 DOI:10.1074/jbc.M109173200 |
| Lazarovici P, Primor N, Loew L M., 1986. Purification and pore-forming activity of two hydrophobic polypeptides from the secretion of the Red Sea Moses sole (Pardachirus marmoratus). Journal of Biological Chemistry, 261(35): 16704–16713 |
| Li H X, Lu X J, Li C H, et al, 2014. Molecular characterization and functional analysis of two distinct liver-expressed antimicrobial peptide 2 (LEAP-2) genes in large yellow croaker (Larimichthys crocea). Fish & Shellfish Immunology, 38(2): 330–339 |
| Li Z P, Chen D W, Pan Y Q, et al, 2016. Two isoforms of piscidin from Malabar grouper, Epinephelus malabaricus:Expression and functional characterization. Fish & Shellfish Immunology, 57: 222–235 |
| Lin W C, Chang H Y, Chen J Y., 2016. Electrotransfer of the tilapia piscidin 3 and tilapia piscidin 4 genes into skeletal muscle enhances the antibacterial and immunomodulatory functions of Oreochromis niloticus. Fish & Shellfish Immunology, 50: 200–209 |
| Livak K J, Schmittgen T D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25(4): 402–408 DOI:10.1006/meth.2001.1262 |
| Lu X J, Chen J, Huang Z A, et al, 2011. Identification and characterization of a novel cathelicidin from ayu, Plecoglossus altivelis. Fish & Shellfish Immunology, 31(1): 52–57 |
| Niu S F, Jin Y, Xu X, et al, 2013. Characterization of a novel piscidin-like antimicrobial peptide from Pseudosciaena crocea and its immune response to Cryptocaryon irritans. Fish & Shellfish Immunology, 35(2): 513–524 |
| Pan C Y, Chen J Y, Cheng Y S, et al, 2007. Gene expression and localization of the epinecidin-1 antimicrobial peptide in the grouper (Epinephelus coioides), and its role in protecting fish against pathogenic infection. DNA and Cell Biology, 26(6): 403–413 DOI:10.1089/dna.2006.0564 |
| Pan C Y, Tsai T Y, Su B C, et al, 2017. Study of the antimicrobial activity of tilapia piscidin 3 (TP3) and TP4 and their effects on immune functions in hybrid tilapia (Oreochromis spp. ). PLoS One, 12(1): e0169678 DOI:10.1371/journal.pone.0169678 |
| Peng K C, Lee S H, Hour A L, et al, 2012. Five different piscidins from Nile tilapia, Oreochromis niloticus:analysis of their expressions and biological functions. PLoS One, 7(11): e50263 DOI:10.1371/journal.pone.0050263 |
| Ruangsri J, Salger S A, Caipang C M, et al, 2012. Differential expression and biological activity of two piscidin paralogues and a novel splice variant in Atlantic cod (Gadus morhua L. ). Fish & Shellfish Immunology, 32(3): 396–406 |
| Salerno G, Parrinello N, Roch P, et al, 2007. cDNA sequence and tissue expression of an antimicrobial peptide, dicentracin; a new component of the moronecidin family isolated from head kidney leukocytes of sea bass, Dicentrarchus labrax. Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology, 146(4): 521–529 DOI:10.1016/j.cbpb.2006.12.007 |
| Silphaduang U, Noga E J., 2001. Antimicrobials:Peptide antibiotics in mast cells of fish. Nature, 414(6861): 268–269 DOI:10.1038/35104690 |
| Subramanian S, Ross N W, MacKinnon S L., 2009. Myxinidin, a novel antimicrobial peptide from the epidermal mucus of hagfish, Myxine glutinosa L. Marine Biotechnology, 11(6): 748–757 DOI:10.1007/s10126-009-9189-y |
| Sun B J, Xie H X, Song Y, et al, 2007. Gene structure of an antimicrobial peptide from mandarin fish, Siniperca chuatsi (Basilewsky), suggests that moronecidins and pleurocidins belong in one family:the piscidins. Journal of Fish Diseases, 30(6): 335–343 DOI:10.1111/jfd.2007.30.issue-6 |
| Sung W S, Lee J, Lee D G., 2008. Fungicidal effect and the mode of action of piscidin 2 derived from hybrid striped bass. Biochemical and Biophysical Research Communications, 371(3): 551–555 DOI:10.1016/j.bbrc.2008.04.107 |
| Tamura K, Peterson D, Peterson N, et al, 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28(10): 2731–2739 DOI:10.1093/molbev/msr121 |
| Umasuthan N, Mothishri M S, Thulasitha W S, et al, 2016. Molecular, genomic, and expressional delineation of a piscidin from rock bream (Oplegnathus fasciatus) with evidence for the potent antimicrobial activities of Of-Pis1 peptide. Fish & Shellfish Immunology, 48: 154–168 |
| Wan M, van der Does A M, Tang X, et al, 2014. Antimicrobial peptide LL-37 promotes bacterial phagocytosis by human macrophages. Journal of Leukocyte Biology, 95(6): 971–981 DOI:10.1189/jlb.0513304 |
| Yang J, Lu X J, Chai F C, et al, 2016. Molecular characterization and functional analysis of a piscidin gene in large yellow croaker (Larimichthys crocea). Zoological Research, 37(6): 347–355 |
| Yin Z X, He W, Chen W J, et al, 2006. Cloning, expression and antimicrobial activity of an antimicrobial peptide, epinecidin-1, from the orange-spotted grouper, Epinephelus coioides. Aquaculture, 253(1): 204–211 |
| Zhou Q J, Su Y Q, Niu S F, et al, 2014. Discovery and molecular cloning of piscidin-5-like gene from the large yellow croaker (Larimichthys crocea). Fish & Shellfish Immunology, 41(2): 417–420 |
| Zhu J, Wang H, Wang J, et al, 2017. Identification and characterization of a β-defensin gene involved in the immune defense response of channel catfish, Ictalurus punctatus. Molecular Immunology, 85: 256–264 DOI:10.1016/j.molimm.2017.03.009 |
| Zhuang Z R, Yang X D, Huang X Z, et al, 2017. Three new piscidins from orange-spotted grouper (Epinephelus coioides):Phylogeny, expression and functional characterization. Fish & Shellfish Immunology, 66: 240–253 |