 2018, Vol. 49
2018, Vol. 49

投喂海水小球藻和三角褐指藻对泥蚶成贝镉排出的影响

http://dx.doi.org/10.11693/hyhz20170800205
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 许家辉, 陈世雄, 俞兴发, 葛亚明, 刘俊稚, 周超. 2018.
- XU Jia-Hui, CHEN Shi-Xiong, YU Xing-Fa, GE Ya-Ming, LIU Jun-Zhi, ZHOU Chao. 2018.
- 投喂海水小球藻和三角褐指藻对泥蚶成贝镉排出的影响
- EFFECTS OF FEEDING CHLORELLA SALINA AND PHAEODACTYLUM TRICORNUTUM ON CADMIUM DEPURATION FROM ADULT CLAM TEGILLARCA GRANOSA
- 海洋与湖沼, 49(3): 624-629
- Oceanologia et Limnologia Sinica, 49(3): 624-629.
- http://dx.doi.org/10.11693/hyhz20170800205
-
文章历史
- 收稿日期:2017-08-10
- 收修改稿日期:2017-11-12
引用本文
许家辉, 陈世雄, 俞兴发, 葛亚明, 刘俊稚, 周超. 2018. 投喂海水小球藻和三角褐指藻对泥蚶成贝镉排出的影响[J]. 海洋与湖沼, 49(3): 624-629
XU Jia-Hui, CHEN Shi-Xiong, YU Xing-Fa, GE Ya-Ming, LIU Jun-Zhi, ZHOU Chao. 2018. EFFECTS OF FEEDING CHLORELLA SALINA AND PHAEODACTYLUM TRICORNUTUM ON CADMIUM DEPURATION FROM ADULT CLAM TEGILLARCA GRANOSA[J]. Oceanologia et Limnologia Sinica, 49(3): 624-629.
投喂海水小球藻和三角褐指藻对泥蚶成贝镉排出的影响
1.
浙江海洋大学海洋科学与技术学院 舟山 316022;
2. 浙江海洋大学创新应用研究院 舟山 316022
2. 浙江海洋大学创新应用研究院 舟山 316022
摘要:通过监测泥蚶不同组织器官镉含量的变化及构建相应泥蚶镉排出动力学模型,研究了投喂两种饵料微藻——海水小球藻(Chlorella salina)和三角褐指藻(Phaeodactylum tricornutum)对泥蚶(Tegillarca granosa)成贝镉排出的影响。结果表明:投喂饵料微藻对泥蚶外套膜、鳃、内脏团和肌肉的镉排出均有明显促进作用(P < 0.05),且投喂混合藻种的效果优于投喂单一藻种。经10d暂养净化后,泥蚶不同组织器官的镉排出率大小依次为内脏团>鳃>外套膜>肌肉,其中投喂混合藻时可食用部分(即肌肉部分)的镉排出率和动力学模型拟合得到的排出速率均比不投喂饵料微藻的对照高出1倍左右[30.02% vs16.13%;0.036μg/(g·d)vs0.017μg/(g·d)];同时,通过上述动力学模型还可算得,投喂饵料微藻的泥蚶体内镉的生物半衰期可比对照组缩短14—22d,其食用部分预期达标时间可缩短12—13d。
关键词:饵料微藻 泥蚶 组织器官 镉排出 动力学模型
EFFECTS OF FEEDING CHLORELLA SALINA AND PHAEODACTYLUM TRICORNUTUM ON CADMIUM DEPURATION FROM ADULT CLAM TEGILLARCA GRANOSA
1.
College of Marine Science and Technology, Zhejiang Ocean University, Zhoushan 316022, China;
2. Innovation and Application Institute, Zhejiang Ocean University, Zhoushan 316022, China
2. Innovation and Application Institute, Zhejiang Ocean University, Zhoushan 316022, China
Abstract:
By monitoring cadmium content in different tissues of clam Tegillarca granosa and constructing corresponding kinetic model of cadmium depuration, the effects of feeding two species of microalgae——Chlorella salina and Phaeodactylum tricornutum on cadmium depuration from adult T. granosa was studied. The results show that both the bait microalgae promoted significantly the cadmium depuration in all tissues of T. granosa (P < 0.05), and the feeding of the two microalgae mixture performed even better. In our 10-day experiment, the cadmium depuration efficiency in different tissues was in the order of viscera > gill > mantle > muscle. Specifically, in the muscle, the edible tissue, both cadmium depuration efficiency and rate of the mixture feeding scheme are about one fold higher than the control[30.02% vs 16.13%; 0.036 μg/(g·d) vs 0.017 μg/(g·d)]. Moreover, according to the constructed kinetic functions of cadmium depuration, by feeding the bait microalgae, the biological half-life (B1/2) of cadmium in all tested tissues could be shortened by 14-22 days, while the expected time for cadmium content in muscle to meet the corresponding national standards would be shortened by 12-13 days. Therefore, this study provided a technical way for heavy metal depuration from living shellfish in terms of food safety, and scientific instructions to bait microalgae selection in shellfish culture in pollution ecology.
Key words:
microalgal diet
Tegillarca granosa
tissues
cadmium depuration
kinetic model
贝类营养丰富、风味独特, 深受国内外消费者喜爱。我国贝类资源丰富, 是世界贝类生产和出口第一大国。然而, 随着我国沿海经济的快速发展, 大量重金属污染物(汞、镉、铅、铬、类金属砷等)被排入水体, 造成近岸海域海水水质恶化。滤食性贝类因其在水生生态系统中的特定生态位, 及其摄食行为特殊、移动能力差、生活方式固定等生活特性, 成为对重金属富集最强的水产品, 许多地区贝类产品均存在不同程度重金属超标, 其中镉、铅超标尤为严重(母清林等, 2013; 蔡艳等, 2016)。与其它污染相比, 重金属污染具有长期性、累积性、潜伏性和不可逆性等特点, 危害大、治理成本高。重金属污染会扰乱贝类自身正常的生命活动, 降低贝类产品品质和产量, 从而制约我国水产养殖业发展; 还会通过食物链传递对人类健康及水体生态环境带来严重危害(Haberkorn et al, 2014)。因此, 重金属污染问题已成为威胁我国贝类养殖业发展和人群健康的重要安全隐患, 亟待解决。
目前能有效脱除贝类体内重金属的方法不为多见, 净水暂养尚可在一定程度上降低贝类体内重金属含量(崔正国等, 2015), 但耗时长且去除率有限; 向养殖水体投加某些化学添加剂可促进贝类重金属排出, 但存在成本高、影响水质等缺点而阻碍其推广(张双灵等, 2013; Chen et al, 2016)。微藻在贝类食物链中占有重要地位, 是大部分贝类育苗场的唯一食源(陈自强等, 2013)。值得注意的是, 微藻也具有超强的重金属富集能力, 很可能在贝类排出重金属中发挥重要作用, 但目前相关报道尚不多见(汪远丽等, 2012; He et al, 2014)。
泥蚶(Tegillarca granosa), 俗称血蚶、粒蚶等, 是我国浙江、福建、广东等东南沿海省市的主要养殖贝类和重要出口创汇水产品之一。本文以泥蚶为研究对象, 探讨了两种常用饵料微藻——海水小球藻和三角褐指藻对泥蚶不同组织器官中镉排出率的影响, 并通过一级动力学模型拟合得到泥蚶镉排出速率k2和生物半衰期B1/2, 预测了通过饲喂饵料微藻促进泥蚶镉排出使其达到食用安全限值的周期, 以期为贝类重金属污染防治提供科学依据和数据支持。
1 材料与方法 1.1 泥蚶来源及暂养实验泥蚶取自浙江省舟山市朱家尖某水产养殖场, 平均壳长(2.80±0.20)cm, 清洗并除掉外壳附着物, 于新鲜海水中暂养3d。期间不投喂饵料, 用增氧泵(EP-9000, 创星电器)连续曝气以保证水中有充足氧气, 每日更换新鲜海水, 并剔除死亡或行为异常的个体。新鲜海水取自舟山市长峙岛附近海域, 经沉淀、砂滤后使用, 其盐度20—22、pH 8.05—8.13、镉浓度小于0.001μg/L。
1.2 饵料微藻及预培养本研究所用饵料微藻为海水小球藻(Chlorella salina)和三角褐指藻(Phaeodactylum tricornutum), 由本校国家海洋设施养殖工程技术研究中心提供, 用f/2培养基于恒温光照培养箱内无菌培养扩繁(温度22℃, 光照强度6000lx, 光照周期12:12)。配制f/2培养基所用新鲜海水同样取自舟山市长峙岛附近海域, 经煮沸、抽滤后使用。
1.3 泥蚶镉富集及排出实验方案泥蚶镉暴露与富集:取暂养后的健康泥蚶放入含20L新鲜海水的塑料箱中(58cm×44cm×34cm), 用1.0g/L的镉母液(由CdCl2·2H2O、少量稀盐酸(v:v=1:4)及高纯水配制而成)调节海水使其中Cd2+浓度为100μg/L。暴露富集5d, 期间不投喂饵料, 每隔24h更换新鲜海水并重新染毒一次。
泥蚶镉排出:取暴露富集结束后的泥蚶转移至清洁新鲜海水中, 设不投喂微藻(对照)和投喂三角褐指藻、投喂海水小球藻及投喂两种混合藻(细胞密度1:1)四个处理组。每天定时投喂2次(藻细胞密度为2.0—4.0×104cell/mL), 每24h更换新鲜海水一次, 保持水温(20±2)℃, 连续曝气。每处理设3个平行, 每天取样一次, 用超纯水洗净贝壳外附着物后迅速–20℃冷冻保存。
1.4 样品镉含量测定及镉排出率计算将泥蚶样品从冰箱中取出解冻, 取出软体组织并分组织解剖(外套膜、肌肉、鳃、内脏团)、烘干、研碎。用分析天平(AL204, 瑞士梅特勒)准确称取(0.1±0.05)g干样品, 按照《海洋监测规范》(GB17378.6-2007)中的方法消解, 用电感耦合等离子体发射光谱仪(Agilent 5100 ICP-OES)测定镉含量。各组织中镉排出率通过式(1)计算:
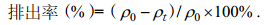 (1)
(1)式中, ρ0、ρt分别表示泥蚶组织初始镉含量与经过t时间后的泥蚶组织镉含量(μg/g)。
1.5 数据处理与动力学模型构建结果中所有数据均为三个平行的平均值, 用SPSS18.0统计软件Bonferroni法对不同处理的数据进行单因素方差分析(ANOVA), P < 0.05为差异显著, P < 0.01为差异极显著。
此外, 根据Kahle等(2002)研究结果, 泥蚶对镉的排出过程可视为一级动力学反应, 体内镉浓度(c)与排放时间(t)的关系可表示为方程式(2):
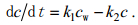 (2)
(2)式中, k1、k2分别为生物吸收速率常数和排出速率常数, cw为水中镉浓度(μg/L)。
由于排出实验在清洁海水中进行, 故cw近似为0;泥蚶对排出镉的重吸作用也可基本忽略, 即k1也视为0, 则方程式(2)可转换为方程式(3):
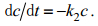 (3)
(3)对方程式(3)求解可得排出过程中任意t时刻泥蚶体内的镉含量(ct)为:
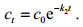 (4)
(4)式中, c0为初始时泥蚶体内的镉含量(μg/g), t为排出时间(d)。
最后, 将泥蚶体内镉含量ct与排出时间t按(4)式进行拟合, 即可得泥蚶体内镉排出速率常数k2。同时, 可用式(5)计算泥蚶体内镉的生物代谢半衰期B1/2(d):
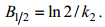 (5)
(5)投喂饵料微藻后泥蚶外套膜中的镉含量变化如图 1所示。可见, 对投喂饵料微藻处理组, 实验初期(0—2d)泥蚶外套膜镉排出不明显, 之后排出率持续增大, 而对照组泥蚶外套膜镉含量4d后才开始持续降低。经10d的净水养殖后, 对照组泥蚶外套膜的镉含量从13.33μg/g降至10.98μg/g, 排出率为17.62%;而各投喂饵料微藻处理组中泥蚶外套膜的镉排出率明显高于对照组(P < 0.05), 投喂三角褐指藻、海水小球藻和混合藻的泥蚶外套膜镉排出率分别为30.46%、28.15%、29.74%, 各饵料组间无明显差异(P > 0.05)。
 |
| 图 1 泥蚶外套膜中镉含量的变化 Fig. 1 Change of Cd content in the mantle of T. granosa |
外套膜具有辅助摄食、呼吸、运动等功能, 是泥蚶等双壳贝类与海水直接接触的组织器官, 其对重金属的富集程度相对较高, 且在自然条件下排出过程较为缓慢。如, 虾夷扇贝外套膜镉排出速率仅为0.008μg/(g·d)(王丽丽等, 2012), 菲律宾蛤仔外套膜的镉排出速率为0.013μg/(g·d)(姜会超等, 2014)。本实验中未投喂饵料微藻的对照组泥蚶外套膜镉排出速率为0.019μg/(g·d), 而投喂混合藻处理的泥蚶外套膜镉排出速率可达0.038μg/(g·d), 是对照组的2倍。研究表明, 双壳贝类外套膜或表皮等组织器官中重金属的富集、迁移过程非常复杂, 包括体表吸附、黏液滞留、脂质渗透等多种形式(Ettajani et al, 2001), 因此饵料微藻对泥蚶外套膜中镉排出的促进机理有待进一步深入研究。
2.2 饵料微藻对泥蚶鳃镉排出的影响投喂饵料微藻后泥蚶鳃的镉含量如图 2所示。在100μg/L镉暴露富集5d后, 泥蚶鳃中镉的平均含量为16.17μg/g, 高于外套膜中镉的富集量。实验期间, 各组泥蚶鳃内镉含量均呈持续下降, 但未投喂饵料微藻的对照组泥蚶腮内镉含量下降速率明显低于投喂饵料微藻的处理组。经过10d净化排放后, 对照组鳃镉的排出率为20.87%, 略高于外套膜; 与外套膜结果相似, 投喂饵料微藻显著促进了泥蚶鳃镉的排出(P < 0.05), 投喂三角褐指藻、海水小球藻、混合藻后的鳃镉排出率分别为44.12%、45.56%、47.44%。
 |
| 图 2 泥蚶鳃中镉含量的变化 Fig. 2 Change of Cd content in the gill of T. granosa |
鳃作为贝类的呼吸器官, 是金属离子从外界环境进入贝类体内的主要渠道之一, 也是金属在贝类体内暂存的主要富集部位之一。鳃丝有很大的比表面积, 上面分布着多种免疫活性酶触位点, 而饵料微藻中丰富的氨基酸与不饱和脂肪酸能够刺激其中超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)等免疫活性酶的表达(Chandurvelan et al, 2012)。同时, 饵料微藻内丰富的维生素C是一种良好的还原剂, 能够促进贝类体内氧化型谷胱甘肽的还原生成还原型谷胱甘肽, 从而利于与金属离子结合排出体外(He et al, 2014)。这些可能是投喂饵料微藻加速泥蚶鳃部镉排出的原因。
2.3 饵料微藻对泥蚶内脏团镉排出的影响如图 3所示, 在100μg/L镉暴露富集5d后, 泥蚶内脏团镉的平均含量高于外套膜和鳃, 为18.25μg/g。投喂饵料微藻极显著地促进了泥蚶内脏团中的镉排出(P < 0.01), 且投喂不同饵料微藻处理之间也存在显著差异(P < 0.05)。经过10d净水养殖后, 未投喂饵料微藻的对照组内脏团中镉的排出率为23.52%;而投喂三角褐指藻、海水小球藻和混合藻的泥蚶内脏团中镉排出率分别达45.16%、50.42%、54.92%, 即饵料微藻对内脏团镉排出促进程度的大小依次为混合藻 > 海水小球藻 > 三角褐指藻。
 |
| 图 3 泥蚶内脏团中镉含量的变化 Fig. 3 Change of Cd content in the viscera of T. granosa |
一般认为, 重金属与生物机体合成的金属硫蛋白(Metallothioneins, MTs)形成难溶性复合物而排出体外是生物解毒的重要机制(Moore et al, 2002)。由于泥蚶MTs合成场所主要位于消化腺上皮细胞和肝胰腺组织, 因此内脏团是泥蚶产生镉解毒作用的重要部位(解家松等, 2011)。此外, 肝胰腺也是泥蚶消化营养物质的重要场所, 具有旺盛的新陈代谢, 对体内有毒物质排出体外起着重要作用(张春丹等, 2012)。不同饵料微藻种类对泥蚶内脏团重金属排出作用的差异, 很可能与微藻细胞中营养成分和含量的不同有关:海水小球藻含有丰富的游离氨基酸、核酸、甾醇、亚麻酸等成分(Lee et al, 2013), 可为MTs合成和活性诱导等过程提供原料, 从而促进其与重金属的结合和排出; 三角褐指藻的核酸、氨基酸等含量不如海水小球藻, 但糖类、EPA等不饱和脂肪酸的含量较高(王晓燕等, 2014), 能给机体新陈代谢提供充足的能量, 从加速体内物质循环的角度有利于重金属污染物的排出。投喂混合藻可使上述营养物质得到互补, 因此更有利于重金属污染物的排出。
2.4 饵料微藻对泥蚶肌肉中镉排出的影响泥蚶肌肉中的镉含量变化如图 4所示。对比图 1—3可见, 暴露染毒后泥蚶肌肉中的镉含量显著低于外套膜、鳃和内脏团中的镉含量(P < 0.05)。这是因为多数贝类只有当内脏、鳃等组织器官重金属积累过度时才会在肌肉积累, 且只有较少部分随血液循环富集于肌肉中(张丽岩等, 2010; 霍礼辉等, 2012)。同时, 由于肌肉中缺乏对镉的调控机制和解毒途径, 使得肌肉与镉的结合度较其他组织器官更为牢固, 一旦富集短期内不易排出(Hervéfernández et al, 2010; 廖艳等, 2014), 这与本研究结论一致, 本实验中经过10d的净化排毒后, 对照组泥蚶肌肉镉排出率仅为16.13%, 明显低于外套膜、鳃和内脏团。而与投喂饵料微藻对泥蚶其他三个部位镉排出的影响规律一致, 投加饵料微藻同样显著促进了泥蚶肌肉的镉排出(P < 0.01), 投喂三角褐指藻、海水小球藻、混合藻组的泥蚶肌肉镉排出率分别为27.85%、26.11%、30.02%。肌肉是贝类产品的重要食用部位, 本研究结论有助于有效降低泥蚶肌肉镉含量以提升贝类食品安全。
 |
| 图 4 泥蚶肌肉中镉含量的变化 Fig. 4 Change of Cd content in the muscle of T. granosa |
根据一级动力学模型, 对投喂不同饵料微藻和对照条件下泥蚶各组织器官中的镉含量变化分别进行非线性拟合, 得到的镉排出动力学参数如表 1所示(R2>0.91)。
表 1 各处理下泥蚶各组织器官镉排出的动力学参数 Tab. 1 Kinetic parameters of Cd release from different tissue organs of T. granosa in different treatment
| 组织器官 | 处理 | k2(μg/g·d) | B1/2(d) | R2 |
| 外套膜 | 三角褐指藻 | 0.036 | 19 | 0.9123 |
| 海水小球藻 | 0.033 | 21 | 0.9813 | |
| 混合藻 | 0.038 | 18 | 0.9661 | |
| 对照组 | 0.020 | 36 | 0.9914 | |
| 鳃 | 三角褐指藻 | 0.062 | 11 | 0.9420 |
| 海水小球藻 | 0.066 | 10 | 0.9636 | |
| 混合藻 | 0.067 | 10 | 0.9755 | |
| 对照组 | 0.027 | 26 | 0.9515 | |
| 内脏团 | 三角褐指藻 | 0.066 | 10 | 0.9830 |
| 海水小球藻 | 0.074 | 9 | 0.9818 | |
| 混合藻 | 0.083 | 8 | 0.9850 | |
| 对照组 | 0.029 | 24 | 0.9802 | |
| 肌肉 | 三角褐指藻 | 0.036 | 19 | 0.9502 |
| 海水小球藻 | 0.033 | 21 | 0.9174 | |
| 混合藻 | 0.036 | 19 | 0.9249 | |
| 对照组 | 0.017 | 41 | 0.9204 |
由表 1可见, 投喂饵料微藻对泥蚶各主要组织器官中的镉排出速率k2均有明显促进作用(P < 0.05), 外套膜、鳃、内脏团和肌肉中的镉排出速率分别可比对照高出0.65—0.90倍、1.30—1.48倍、1.28—1.86倍、0.94—1.12倍; 且混合投喂饵料微藻的排出速率比单独投喂时更大, 即混合投喂对镉的促排作用更明显。此外, 动力学模型拟合结果显示, 投喂饵料微藻后各组织器官中镉的生物代谢半衰期B1/2能够比对照组缩短14—22d, 这从生物半衰期角度进一步展示了投喂饵料微藻对促进泥蚶镉排出的重要作用。
根据我国《食品安全国家标准食品中污染物限量》(GB2762-2017), 双壳类水产动物食用部分的镉限量标准为2.0μg/g(鲜重), 而本实验中泥蚶可食用部分(即肌肉部分)初始镉含量为3.018μg/g(鲜重, 实验所用泥蚶肌肉部分含水率为71.1%), 即本实验中泥蚶可食用部分初始镉含量已超标50.9%。运用上述动力学模型对本实验中泥蚶经暂养净化后其可食用部分镉含量达到国家标准要求所需天数进行了理论测算, 结果表明, 对照组泥蚶肌肉镉含量净化达标天数预计为25d, 投喂三角褐指藻、海水小球藻、混合藻的处理组泥蚶肌肉镉净化达标天数分别预计为12d、13d和12d, 分别比对照缩短13d、12d和13d。
综合以上数据, 泥蚶各组织器官对镉的富集能力与排出速率大小顺序均为内脏团 > 鳃 > 外套膜 > 肌肉, 该富集规律和霍礼辉等(2012)与王召根等(2013)的研究结果总体一致。饵料微藻对泥蚶不同组织器官镉排出促进程度的不同, 可能与其生物酶活性差异、解毒机理不同、营养物质分配不均等因素有关(陈肖肖等, 2013; Qiu, 2015)。此外, 泥蚶镉的富集与排出还可能受到饵料微藻种类、贝藻比例、饲喂时期和外界环境条件等诸多因素影响, 有待进一步研究。
3 结论本研究在构建的“饵料微藻-泥蚶”人工微宇宙体系中, 通过实验和动力学模型拟合探讨了投喂饵料微藻对泥蚶镉排出的影响, 取得了以下主要结论:投喂三角褐指藻、海水小球藻和二者混合藻对泥蚶各主要组织器官(内脏团、鳃、外套膜和肌肉)的镉排出均有明显促进作用, 能够明显提升镉的最大排出速率, 缩短其生物半衰期和净化达标时间, 且投喂混合藻的效果优于投喂单一藻。总之, 本研究能够从污染生态学角度为贝类养殖中的重金属污染防治和饵料微藻筛选等提供科学指导。
参考文献
| 王召根, 吴洪喜, 陈肖肖, 等, 2013. 年龄和环境条件对泥蚶富集重金属镉和铜的影响. 生态学报, 33(21): 6869–6875 |
| 王丽丽, 夏斌, 陈碧鹃, 等, 2012. 虾夷扇贝对镉的蓄积和排放规律. 海洋环境科学, 31(2): 159–162 |
| 王晓燕, 邢欢, 钟韵山, 等, 2014. 三角褐指藻和小球藻营养成分的对比分析. 天津科技大学学报, 29(2): 11–15 |
| 母清林, 王晓华, 佘运勇, 等, 2013. 浙江近岸海域贝类中重金属和贝毒污染状况研究. 海洋科学, 37(1): 87–91 |
| 张双灵, 张忍, 于春娣, 等, 2013. 贝类重金属脱除技术的研究现状与进展. 食品安全质量检测学报, 4(3): 857–862 |
| 张丽岩, 宋欣, 高玮玮, 等, 2010. Cd2+对青蛤(Cyclina sinensis)的毒性及蓄积过程研究. 海洋与湖沼, 41(3): 418–421 DOI:10.11693/hyhz201003019019 |
| 张春丹, 周君, 李晔, 等, 2012. 重金属胁迫对泥蚶(Tegillarca granosa)能量代谢酶转录水平的研究. 海洋与湖沼, 43(5): 919–923 DOI:10.11693/hyhz201205007007 |
| 汪远丽, 曲克明, 单宝田, 等, 2012. 重金属在小球藻-菲律宾蛤仔食物链上的传递与累积. 渔业科学进展, 33(1): 79–85 |
| 陈自强, 寿鹿, 廖一波, 等, 2013. 微藻饵料对双壳贝类幼体生长影响的研究进展. 科技通报, 29(7): 46–55 |
| 陈肖肖, 高业田, 吴洪喜, 等, 2013. 重金属Cd2+和Cu2+胁迫下泥蚶消化酶活性的变化. 生态学报, 33(24): 7690–7698 |
| 姜会超, 刘爱英, 宋秀凯, 等, 2014. 混合暴露条件下菲律宾蛤仔不同组织对几种重金属的富集与释放特征. 渔业科学进展, 35(3): 82–90 DOI:10.11758/yykxjz.20140312 |
| 崔正国, 苑旭洲, 崔毅, 等, 2015. 虾夷扇贝(Patinopecten yessoensis)对铅和镉的生物富集与释放规律. 渔业科学进展, 36(3): 116–124 DOI:10.11758/yykxjz.20150318 |
| 解家松, 许婷, 刘玮, 等, 2011. 泥蚶金属硫蛋白的鉴定、原核重组表达及其组织细胞分布. 中国水产科学, 18(5): 955–964 |
| 廖艳, 蒋志红, 吴晓萍, 等, 2014. 企鹅珍珠贝肉的一般化学组成及镉的分布. 食品科学, 35(2): 201–204 DOI:10.7506/spkx1002-6630-201402038 |
| 蔡艳, 周亦君, 吴晓艺, 等, 2016. 3种海洋贝类重金属污染及食用风险评价研究. 核农学报, 30(6): 1126–1134 DOI:10.11869/j.issn.100-8551.2016.06.1126 |
| 霍礼辉, 林志华, 尤仲杰, 等, 2012. 泥蚶不同组织器官对重金属(Cu、Pb、Cd)的富集规律. 海洋环境科学, 31(3): 395–399 |
| Chandurvelan R, Marsden I D, Gaw S, et al, 2012. Impairment of greenlipped mussel (Perna canaliculus) physiology by waterborne cadmium:Relationship to tissue bioaccumulation and effect of exposure duration. Aquatic Toxicology, 124-125: 114–124 DOI:10.1016/j.aquatox.2012.07.013 |
| Chen C F, Shen W L, Gu H L, et al, 2016. Vitellogenin's putative role in Tegillarca granosa's cadmium detoxification. Genes & Genomics, 39(2): 143–154 |
| Ettajani H, Berthet B, Amiard J C, et al, 2001. Determination of cadmium partitioning in microalgae and oysters:contribution to the assessment of trophic transfer. Archives of Environmental Contamination and Toxicology, 40(2): 209–221 DOI:10.1007/s002440010165 |
| Haberkorn H, Lambert C, Le Goïc N, et al, 2014. Cellular and biochemical responses of the oyster Crassostrea gigas to controlled exposures to metals and Alexandrium minutum. Aquatic Toxicology, 147: 158–167 DOI:10.1016/j.aquatox.2013.12.012 |
| He J S, Chen J P, 2014. A comprehensive review on biosorption of heavy metals by algal biomass:Materials, performances, chemistry, and modeling simulation tools. Bioresource Technology, 160: 67–78 DOI:10.1016/j.biortech.2014.01.068 |
| Hervé-Fernández P, Houlbrèque F, Boisson F, et al, 2010. Cadmium bioaccumulation and retention kinetics in the Chilean blue mussel Mytilus chilensis:seawater and food exposure pathways. Aquatic Toxicology, 99(4): 448–456 DOI:10.1016/j.aquatox.2010.06.004 |
| Kahle J, Zauke G P, 2002. Bioaccumulation of trace metals in the copepod Calanoides acutus, from the Weddell Sea (Antarctica):comparison of two-compartment and hyperbolic toxicokinetic models. Aquatic Toxicology, 59(1-2): 115–135 DOI:10.1016/S0166-445X(01)00245-4 |
| Lee Y J, Hong Y J, Kim J Y, et al, 2013. Dietary Chlorella protects against heterocyclic amine-induced aberrant gene expression in the rat colon by increasing fecal excretion of unmetabolized PhIP. Food & Chemical Toxicology, 56: 272–277 |
| Moore M N, Allen J I, 2002. A computational model of the digestive gland epithelial cell of marine mussels and its simulated responses to oil-derived aromatic hydrocarbons. Marine Environmental Research, 54(3-5): 579–584 DOI:10.1016/S0141-1136(02)00166-6 |
| Qiu Y W, 2015. Bioaccumulation of heavy metals both in wild and mariculture food chains in Daya Bay, South China. Estuarine Coastal & Shelf Science, 163: 7–14 |