 2018, Vol. 49
2018, Vol. 49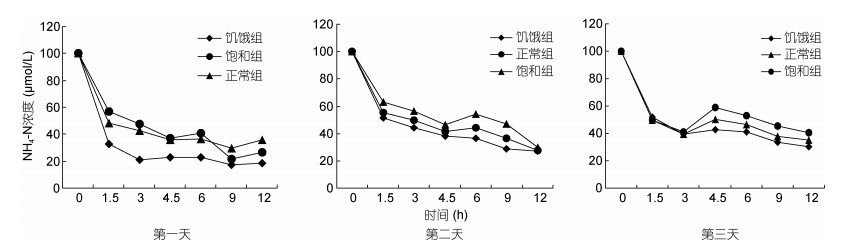


 饥饿处理
饥饿处理 正常对照
正常对照 饱和处理。
饱和处理。
铜藻(Sargassum horneri)在营养限制胁迫后对NH4-N的超补偿吸收研究

http://dx.doi.org/10.11693/hyhz20180300068
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 李大鹏, 马增岭, 李慧, 丁刚, 辛美丽, 吴海一, 郭文. 2018.
- LI Da-Peng, MA Zeng-Ling, LI Hui, DING Gang, XIN Mei-Li, WU Hai-Yi, GUO Wen. 2018.
- 铜藻(Sargassum horneri)在营养限制胁迫后对NH4-N的超补偿吸收研究
- NH4-N OVER-COMPENSATORY UPTAKE OF SARGASSUM HORNERI UNDER THE STRESS OF NUTRIENTS DEFICIENCY
- 海洋与湖沼, 49(4): 904-909
- Oceanologia et Limnologia Sinica, 49(4): 904-909.
- http://dx.doi.org/10.11693/hyhz20180300068
-
文章历史
- 收稿日期:2018-03-27
- 收修改稿日期:2018-04-25
引用本文
李大鹏, 马增岭, 李慧, 丁刚, 辛美丽, 吴海一, 郭文. 2018. 铜藻(Sargassum horneri)在营养限制胁迫后对NH4-N的超补偿吸收研究[J]. 海洋与湖沼, 49(4): 904-909

LI Da-Peng, MA Zeng-Ling, LI Hui, DING Gang, XIN Mei-Li, WU Hai-Yi, GUO Wen. 2018. NH4-N OVER-COMPENSATORY UPTAKE OF SARGASSUM HORNERI UNDER THE STRESS OF NUTRIENTS DEFICIENCY[J]. Oceanologia et Limnologia Sinica, 49(4): 904-909.
铜藻(Sargassum horneri)在营养限制胁迫后对NH4-N的超补偿吸收研究
1.
山东省海洋生物研究院 青岛 266104;
2. 温州大学生命与环境科学学院 温州 325035
2. 温州大学生命与环境科学学院 温州 325035
摘要:以经济马尾藻铜藻(Sargassum horneri)为研究材料,研究了其在营养限制胁迫后对NH4-N的超补偿吸收情况。实验分营养限制和恢复营养两个阶段进行,每个阶段均设置饥饿处理组、饱和处理组和正常对照组。铜藻在低营养限制胁迫(饥饿)下培养10天后,恢复营养盐培养3天,分别采用次溴酸盐氧化法和锌镉还原法测定培养液中的NH4-N和NO3-N的浓度。本文研究发现,饥饿处理组中铜藻吸收氨氮的速率远高于正常对照组和饱和处理组,在恢复培养第一天时,饥饿处理组对NH4-N的吸收速率最高为14.94μmol/(g·h),与正常对照组及饱和处理组间差异显著。在恢复培养的后两天,三个处理组中铜藻对NH4-N的吸收速率差异慢慢变小,直至最后几乎相同。三组对NO3-N没有表现出较高的吸收,最高仅为6.15μmol/(g·h),结果表明:氮源包括NH4-N和NO3-N时,铜藻优先选择吸收NH4-N。实验后称重测定生长速率:对照组、饥饿处理组和饱和组生长率(SGR)分别为8.48%、8.86%、8.01%,ANOVA方差分析表明,三者存在显著差异(P=0.032 < 0.05),从而证实了铜藻也存在超补偿生长的现象。
关键词:铜藻 营养限制 超补偿 NH4-N吸收 NO3-N吸收
NH4-N OVER-COMPENSATORY UPTAKE OF SARGASSUM HORNERI UNDER THE STRESS OF NUTRIENTS DEFICIENCY
1.
Marine Biology Institute of Shandong Province, Qingdao 266104, China;
2. College of Life and Environmental Science, Wenzhou University, Wenzhou 325035, China
2. College of Life and Environmental Science, Wenzhou University, Wenzhou 325035, China
Abstract:
In this paper, Sargassum horneri was used as experimental material for the study of NH4-N over-compensatory uptake under laboratory conditions. Sargassum horneri was thoroughly cleaned, epibiotics carefully removed, and cultured in beakers in an illuminating incubator under 15℃, 100μmol/(m2·s), L:D 12h/12h, 30 salinity and pH 7.5-8.0.
The experiments included two periods. Firstly, Sargassum horneri was cultured for 10 days under the condition of nutrients stress, i.e. nutrients deficiency and nutrients enrichment, with fresh sea water one as the control. Secondly, the nutrients stress was lifted; the treated and the control were simultaneously cultured for another 3 days under the same nutrients supply with daily medium change. The experiments were carried out in 2L beakers with 3g Sargassum horneri and 1L medium, and the concentration of NH4-N and NO3-N was 100 and 280μmol/L respectively. Triplicate were set for each group. During the experiment, the NH4-N, NO3-N uptake rate were measured periodically.
After subjecting to the nutrients deficiency, Sargassum horneri showed a strong ability of NH4-N uptake. The uptake rate of NH4-N of the starved group, the control group and the fed group, was 14.94, 11.54, and 9.63μmol/(g·h), respectively. The starved had the highest uptake rate of NH4-N but its highest uptake rate of NO3-N was only 6.15μmol/(g·h), much smaller than that of NH4-N, indicating that Sargassum horneri assimilated NH4-N first. As a result, the concentration of NO3-N in the beakers was 3 times higher of the NH4-N.
As the culture time proceeded, difference between the treated and the control decreased gradually and finally were equivalent. The special growth rate (SGR) in the group of the starved, the control and the fed was 8.86%, 8.48%, and 8.01% respectively. The difference of the growth rate between the treated and the control was significant (P=0.032 < 0.05).
After 10 days of nutrients deficiency, there was a short phase of rapid NH4-N uptake demonstrating over-compensatory growth. Thus, NH4-N over-compensatory uptake of Sargassum horneri could be used as a guidance for cultivation. Moreover, Sargassum horneri have very high productivity and ability of absorbing large quantities of N. It is believed that Sargassum horneri cultivation in large-scale is a good solution to eutrophication in coastal waters.
Key words:
Sargassum horneri
nutrients deficiency
over-compensatory growth
NH4-N uptake
NO3-N uptake
氮是大型海藻正常生理活动所必需的基本营养元素, 自然海水中的氮浓度相对较低, 且随着季节变化氮浓度波动明显, 常常影响海藻生长(徐智广等, 2016)。植物受胁迫后生长生理受限, 若胁迫解除, 植物会快速生长, 以恢复胁迫造成的影响, 此时的生长速率会远大于未受胁迫时的速率, 超补偿生长正是如此(李梅等, 2004; 郭羽丰等, 2005)。Belsky(1986)将植物遭受胁迫后的补偿作用分为超补偿(over compensation)、等补偿(exact compensation)和低补偿(under compensation)三类。超补偿现象体现了生物进化过程中对环境的适应, 在自然界中这是一种常见现象, 最早发现于高等动植物中(邹春静等, 1998)。目前, 对某些生物的补偿生长研究较多, 如海洋微藻(刘晓娟等, 2007)、高等植物(关义新等, 1997)和水产动物(王燕妮等, 2001)等。关于大藻的超补偿现象研究较少, 有研究发现龙须菜(李大鹏等, 2005)和江蓠(刘静雯等, 2001)在营养限制胁迫后恢复到正常营养状态, 二者均对NH4-N均具有超补偿吸收能力。
铜藻(Sargassum horneri)是生物修复海洋生态的首选物种, 也含有膳食纤维、褐藻多糖、褐藻胶、褐藻淀粉等有益生源要素, 具有较高的生态和经济价值(孙建璋等, 2008; 毛玉泽等, 2012)。关于铜藻超补偿生长现象的存在情况以及通过人工调控超补偿现象能否提高铜藻生物产量等问题, 尚未见相关文献报道。对海藻补偿生长和补偿吸收现象的研究既对揭示海藻适应饥饿等环境胁迫的生理生态学对策有重要意义, 同时也为铜藻的繁殖发育和幼苗培育研究及利用海藻缓解海区富营养化提供了理论依据(严国安等, 1995; 张小兵等, 2007)。因此, 本研究采用化学方法分析了饥饿胁迫后的铜藻在恢复到正常营养下对两种氮源的吸收特点和对氨氮的补偿效应, 试图为经济海藻规模化高效率培养利用寻找新的技术途径。
1 材料与方法 1.1 实验材料实验在山东省海洋生物研究院海藻养殖与修复实验室进行。实验所用铜藻(Sargassum horneri)取自山东荣成海区, 选择藻体完整、无损伤腐烂、规格均一的藻体, 将附着在表面的杂藻用镊子去除干净后, 再用灭菌海水冲洗数次。保持藻体密度为3g/L, 用5L三角瓶置于MGC-250P光照培养箱中, 在温度15±0.5℃、光照强度100μmol/(m2·s)、光暗周期12h : 12h、盐度30、pH 7.5—8.0的条件下培养, 海水经煮沸消毒后添加NH4Cl和NaNO3作为氮源。
1.2 培养方法分别设置正常对照组、饱和处理组和饥饿处理组三个预处理组, 以3g/L的海藻密度在3L的锥形瓶中培养, 消毒海水中不加氮源做为饥饿处理组的培养基, 先培养两天左右, 直至培养液中N耗尽, 之后继续培养10天, 期间不更换培养基, 饥饿状态下的藻体色泽呈淡黄色。将消毒海水中添加氮源作为饱和处理组的培养液, 培养期间每天换新的培养液, 目的是给铜藻营造氮加富的营养环境, 培养周期为10天。用消毒海水作为对照组的培养液, 每天更换。
1.3 NH4-N吸收实验将经过处理的藻选取相同部位分别称3g放于盛有1L培养液的烧杯中, 每个处理设置三个平行, 在水温15℃, 光照强度100μmol/(m2·s), 光周期12L : 12D, 盐度30, pH 8.0的条件下培养3天, 每天更换培养液, 调节NO3-N、NH4-N初始浓度分别为280μmol/L、100μmol/L。
分别用次溴酸盐氧化法、锌镉还原法每天检测培养液中NH4-N、NO3-N的含量, 于1.5、3、4.5、6、9、12h取5mL培养液测NH4-N浓度; 于3、6、9、12、24h取2.5mL培养液测NO3-N浓度; 连续测3天, 结束后称重。吸收速率计算公式为
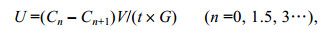
式中, U代表吸收速率, Cn为第n次测定时溶液中氨氮(硝氮)浓度, V为体积, t为培养时间, G为鲜重。
生长率计算公式如下:
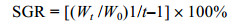
式中, W0表示铜藻开始培养时的重量, Wt为铜藻最后重量, t为培养天数。
2 结果 2.1 铜藻对NH4-N、NO3-N的吸收在恢复营养培养的三天中, 饥饿处理的铜藻对NH4-N均存在明显的快速吸收现象(图 1), 每次测量培养液中NH4-N的浓度由低到高顺序为饥饿处理组、正常对照组、饱和处理组, 而在第一天培养了9h后, 饱和处理组中NH4-N浓度低于正常对照组。第一天饥饿处理组与对照组和饱和处理组间培养液中NH4-N的浓度变化差异明显, 之后两天三者间差异逐渐减小。由图 2可以看出每天的前1.5h三组中的铜藻对NH4-N的吸收速率均高于其余时间对NH4-N的吸收速率, 之后每天三组对NH4-N的吸收速率呈锯齿状波动, 数值基本稳定, 但在培养至4.5—6h时, 一些组之间出现了负吸收速率的情况。饥饿组第一天对NH4-N的吸收速率可达14.94μmol/(g·h), 第二天和第三天也都比较高。孙宏超等(2007)通过铜藻对氨氮的吸收实验, 得出铜藻对海水中NH4-N的平均吸收速率为0.0339μmol/(g·h), 与本实验中饥饿组铜藻对NH4-N的吸收速率相比足以说明铜藻在营养限制胁迫后对NH4-N存在超补偿吸收现象。
 |
| 图 2 铜藻NH4-N的吸收速率随时间的变化 Fig. 2 Temporal change in NH4-N uptake rate of S. horneri |
 |
| 图 3 NO3-N浓度随时间的变化 Fig. 3 Temporal change in NO3-N concentration of medium |
恢复营养培养的第一、二天的前9h, 饥饿处理组培养液中NO3-N浓度明显变小, 与对照组和饱和处理组间差异明显, 第三天三组间没有明显差异。尽管培养液中浓度是NH4-N浓度的2.8倍之高, 但由图 4可以看出恢复营养培养的三天中, 每天铜藻对NO3-N的吸收速率远低于对NH4-N的吸收速率, 其中饥饿处理组对NO3-N的最大吸收速率为3.15μmol/(g·h)。
 |
| 图 4 铜藻NO3-N的吸收速率随时间的变化 Fig. 4 Temporal change in NO3-N uptake rate of S. horneri |
各实验组铜藻的特定生长率如表 1。分析结果得出, 三组中铜藻的特定生长率存在显著差异(P < 0.05), 并且饥饿处理组的特定生长率最大。低氮胁迫后的铜藻在恢复营养后具有较强的超补偿生长能力, 这减轻了不利环境带来的损伤。同时说明铜藻对营养盐胁迫有很强的耐受能力, 这使得它在种间竞争中占据优势。在人工养殖中也可以利用超补偿生长快速增加铜藻生物量。据研究表明植物只在恢复营养后的一段时间存在超补偿生长, 随着时间推移便会恢复到正常水平(金送笛等, 1994; 秦红杰等, 2010)。也有学者对超补偿生长的机制做了解释, 认为处于营养胁迫下的植物代谢水平大幅降低, 当恢复营养后代谢不会立即增强, 因此营养盐提供的能量会更多的用来恢复生长, 此时特定生长率就明显高于持续补给营养组(Bilton et al, 1973; 刘宁宁等, 2002)。
表 1 正常对照组及营养胁迫组铜藻的生长率(平均值±SD, n=3) Tab. 1 Different growth rates of S. horneri between treated and control group
| 组别 | 正常对照 | 饥饿处理 | 饱和处理 |
| SGR(%) | 8.48±0.03 | 8.86±0.02 | 8.01±0.02 |
大型海藻仅能吸收溶解在水中的营养物, 其中氮、磷是制约大型海藻生长的关键限制元素。氮的三种无机化合态分别为NO3-N、NH4-N和NO2-N, 三种形态之间在一定条件下实现互相转化。藻类通过酶将NO3-N先转化为NO2-N, 再变为氮营养的最后利用形态NH4-N。在缺氧水体中, 受反硝化菌的作用NO3-N和NO2-N易被还原为NH4-N。从铜藻对两种氮源的吸收实验结果来看, 实验初期铜藻对NH4-N的吸收速率显著高于对NO3-N的吸收速率, 这与海藻优先选择NH4-N作为N源有关, 胞内相关酶(NR)缺乏也是原因之一。McGlathery等(1996)研究绿藻对氮的吸收和反馈机制时得出, 海藻对NH4+表现出较快的吸收速率, 对NO3–的吸收率较慢, 且一般不会使用NO2–作为氮源。徐智广等研究了海带的氮吸收特性, 发现当NO3-N和NH4-N两种氮源同时存在时, 海藻会优先利用氨氮。同样, 有学者研究了江蓠对氮的吸收情况指出, 无论NH4+的起始浓度是否高于NO3–的浓度, 与NO3–相比两者对NH4+有很强吸收, 猜测这与NH4+是海藻的优先N源有关(钱鲁闽等, 2005)。从大型海藻对氨氮和硝氮两种氮源的吸收机制来分析, 原因可能是海藻对NO3-N的吸收为主动吸收, 需要耗能, 而对NH4-N为被动吸收, 不需耗能。另外, 本研究中铜藻对NH4+的吸收出现了负吸收速率, 随后逐渐恢复正常, 猜测原因为:在恢复营养培养初期, 铜藻吸收了培养中大量的NH4-N, 导致溶液中NH4-N含量急剧减少, 满足不了铜藻的需求, 进而选择吸收NO3-N。
据研究表明:海藻对氨氮吸收包括:短期快速吸收, 藻体内氮稳态调控的吸收(同化阶段), 培养液中营养盐控制的吸收(刘静雯等, 2001; 李大鹏等, 2005)。本实验也符合这一规律, 纵观铜藻对NH4-N吸收速率的变化趋势为:每天恢复营养培养的前期快速吸收, 中期吸收缓慢, 后期微弱吸收。在吸收实验起始的1h内, 吸收速率都达到最大值, 而在1—5h内呈现降低趋势, 5—9h吸收趋于稳定。海藻在营养限制下吸收NH4-N同时, 体内氮形式不断交换, 是阶段出现的原因(冯婧等, 2007)。藻类为了适应环境中营养盐的波动, 刚解除胁迫时会快速吸收氮, 吸收量远大于正常生长需求。李大鹏等(2005)研究龙须菜在营养限制胁迫后对氨氮的吸收时也发现龙须菜在营养限制后对NH4-N的吸收能力较强, 吸收速率远远高于正常培养组和饱和处理组。
海藻N吸收速率受许多因素的影响, 例如N浓度、温度、光照等培养条件以及培养时间等。通常大型海藻的氮吸收与营养供应关系可用米氏动力学方程: V=Vmax·C/(Km+C)来描述。但处于N饥饿状态以及半饥饿状态的海藻在高浓度介质下对NH4-N的吸收动力往往偏离米氏动力学方程, 而表现出起始阶段的快速吸收现象。温珊珊等(2008)在对半饥饿的藻体放于较高浓度NH4-N中测试吸收时, 发现藻体对NH4-N存在早期较短的快吸收, 吸收速率几乎随NH4-N浓度升高而增大, 与动力方程不对应。据分析这一现象的形成表明:藻体体内不同的N库影响着处于氮饥饿状态海藻对N的吸收, 开始的快吸收阶段用于补充由于氮饥饿而产生的N库亏空, 随着氮库不断被填充, N的吸收速率维持稳定, 之后吸收随介质中营养变化, 营养被利用, 导致吸收速率变小。
3.2 大型海藻对营养盐的超补偿吸收现象大型海藻的营养盐吸收能力主要取决于营养盐的含量, 一般情况下, 海水中营养盐含量越丰富, 藻体对其吸收就越快; 另外, 也会受生理情况和环境条件的影响。季节更替致使环境改变, 海藻常常不容易遇到合适的生存环境, 例如海区生物种类繁多, 营养盐供应不足, 从而不能满足海藻正常生长发育的营养盐需求, 遭受饥饿。藻类在胁迫解除后, 为了适应变化, 它们吸收营养生长方面都会高于正常藻体(吴立新等, 2000)。
大型海藻可以吸收大量的氮, 同时可以暂时存在体内, 即使生存条件苛刻, 它们也会吸收过量营养盐, 以满足条件适宜时快速生长的需求, 这作为积极反应, 在生产上具有很大意义(邹定辉等, 2011)。Fujita(1985)研究发现即使在氮限制状态下, 藻体依旧可以依靠体内氮库生长6—14d;研究者通过研究营养盐供应对氨氮吸收的影响表明, 在水体N充足下, 大型海藻也储备营养, 供随后存活所需。段舜山等(2003)报道, 一定的环境胁迫不影响藻类生长, 反而会产生促进作用。由于海藻场大面积消失, 导致铜藻生物量急剧下降, 通过本研究表明适当的环境胁迫可以激发藻体的生长和繁殖潜力, 从而弥补极端环境因子造成的损失。大型海藻在生长过程中会大量吸收养殖水体中的氮、磷等营养元素, 对富营养化水体有很好的净化效果, 是海区生态修复的首选物种之一(Pritchard et al, 2015)。对大型海藻氮吸收特征的研究, 对开发一种经济环保的富营养化去除方法意义重大。
4 结论 4.1饥饿处理后的铜藻对NH4-N存在明显的快速吸收现象, 与正常组差异明显; 铜藻在饥饿处理后对NH4-N吸收存在超补偿吸收现象, 但持续时间较短(2天左右)。
4.2铜藻在营养盐胁迫处理后, 特定生长率存在显著性差异(P < 0.05), 初步验证了大型海藻中除龙须菜外铜藻也有超补偿生长现象。
参考文献
| 王燕妮, 张志蓉, 郑曙明, 2001. 鲤鱼的补偿生长及饥饿对淀粉酶的影响. 水生态学杂志, 21(5): 6–7 |
| 毛玉泽, 蒋增杰, 方建光, 2012. 基于大型藻类的网箱养殖生物修复技术. 中国科技成果,(10): 21–24 |
| 冯婧, 骆其君, 徐志标, 等, 2007. 不同氮源及浓度对条斑紫菜叶状体生长的影响. 宁波大学学报(理工版), 20(1): 19–22 |
| 刘宁宁, 段舜山, 2002. 蛋白核小球藻在光胁迫下的超补偿现象. 生态科学, 21(1): 53–54 |
| 刘晓娟, 段舜山, 李爱芬, 等, 2007. 绿色巴夫藻受紫外(UV-B)胁迫后的超补偿生长效应. 应用生态学报, 18(1): 169–173 |
| 刘静雯, 董双林, 马甡, 2001. 温度和盐度对几种大型海藻生长率和NH4-N吸收的影响. 海洋学报, 23(2): 109–116 |
| 关义新, 戴俊英, 徐世昌, 等, 1997. 玉米花期干旱及复水对植株补偿生长及产量的影响. 作物学报, 23(6): 740–745 |
| 孙宏超, 章守宇, 2007. 铜藻对几种无机盐的吸收动力学的初步研究. 上海海洋大学学报, 16(6): 573–579 |
| 孙建璋, 陈万东, 庄定根, 等, 2008. 中国南麂列岛铜藻Sargassum horneri实地生态学的初步研究. 南方水产, 4(3): 58–63 |
| 严国安, 谭智群, 1995. 藻类净化污水的研究及其进展. 环境工程学报, 3(3): 45–54 |
| 李梅, 徐瑾, 刘志礼, 等, 2004. 锶诱导的氧化胁迫对叉鞭金藻(Dicrateria inornata)的影响. 海洋与湖沼, 35(5): 467–472 |
| 李大鹏, 林贞贤, 2005. 龙须菜(Gracilaria lemaneiformis)在营养限制胁迫后对NH4-N的超补偿吸收研究. 海洋与湖沼, 36(4): 307–312 |
| 吴立新, 董双林, 2000. 水产动物继饥饿或营养不足后的补偿生长研究进展. 应用生态学报, 11(6): 943–946 |
| 邹定辉, 夏建荣, 2011. 大型海藻的营养盐代谢及其与近岸海域富营养化的关系. 生态学杂志, 30(3): 589–595 |
| 邹春静, 徐文铎, 张远, 等, 1998. 沙地云杉幼树对食叶害虫危害的补偿与超补偿效应. 应用生态学报, 9(2): 119–122 |
| 张小兵, 胡章喜, 黄振华, 等, 2007. 三角褐指藻在磷营养限制胁迫下的补偿生长效应. 生态科学, 26(2): 111–114 |
| 金送笛, 李永函, 倪彩虹, 等, 1994. 菹草(Potamogeton crispus)对水中氮、磷的吸收及若干影响因素. 生态学报, 14(2): 168–173 |
| 段舜山, 郭羽丰, 刘振乾, 等, 2003. 四列藻在营养限制胁迫下的超补偿生长研究. 生态学报, 23(7): 1297–1304 |
| 秦红杰, 李敦海, 2010. 铜绿微囊藻高温胁迫后的超补偿生长. 环境科学, 31(7): 1504–1509 |
| 钱鲁闽, 徐永健, 王永胜, 2005. 营养盐因子对龙须菜和菊花江蓠氮磷吸收速率的影响. 应用海洋学学报, 24(4): 546–552 |
| 徐智广, 吴海一, 孙福新, 等, 2016. 不同氮源加富对海带生长、光合固碳和氮吸收特性的影响. 水产学报, 40(4): 577–584 |
| 郭羽丰, 段舜山, 李爱芬, 等, 2005. 四列藻继盐胁迫后的超补偿生长. 海洋科学, 29(5): 37–42 |
| 温珊珊, 张寒野, 何文辉, 等, 2008. 真江蓠对氨氮去除效率与吸收动力学研究. 水产学报, 32(5): 794–803 |
| Belsky A J, 1986. Does herbivory benefit plants? A review of the evidence. The American Naturalist, 127(6): 870–892 DOI:10.1086/284531 |
| Bilton H T, Robins G L, 1973. The effects of starvation and subsequent feeding on survival and growth. Journal of the Fisheries Board of Canada, 30(1): 1–5 DOI:10.1139/f73-001 |
| Fujita R M, 1985. The role of nitrogen status in regulating transient ammonium uptake and nitrogen storage by macroalgae. Journal of Experimental Marine Biology and Ecology, 92(2-3): 283–301 DOI:10.1016/0022-0981(85)90100-5 |
| Mcglathery K J, Pedersen M F, Borum J, 1996. Changes in intracellular nitrogen pools and feedback controls on nitrogen uptake in Chaetomorpha linum (chlorophyta) 1. Journal of Phycology, 32(3): 393–401 DOI:10.1111/j.0022-3646.1996.00393.x |
| Pritchard D W, Hurd C L, Beardall J, et al, 2015. Restricted use of nitrate and a strong preference for ammonium reflects the nitrogen ecophysiology of a light-limited red alga. Journal of Phycology, 51(2): 277–287 DOI:10.1111/jpy.12272 |