 2018, Vol. 49
2018, Vol. 49中国海洋湖沼学会主办。
文章信息
- 高亚平, 方建光, 房景辉, 赵永超, 吉红九, 李文豪, 王晓芹, 李凤雪, 蒋增杰. 2018.
- GAO Ya-Ping, FANG Jian-Guang, FANG Jing-Hui, ZHAO Yong-Chao, JI Hong-Jiu, LI Wen-Hao, WANG Xiao-Qin, LI Feng-Xue, JIANG Zeng-Jie. 2018.
- 长牡蛎和紫贻贝对浒苔微观繁殖体生物控制的可行性研究
- BIOLOGICAL CONTROL FEASIBILITY OF PACIFIC OYSTER CRASSOSTREA GIGAS AND BULE MUSSEL MYTILUS EDULIS ON ULVA PROLIFERA MICROSCOPIC PROPAGULES
- 海洋与湖沼, 49(5): 1116-1122
- Oceanologia et Limnologia Sinica, 49(5): 1116-1122.
- http://dx.doi.org/10.11693/hyhz20180400078
-
文章历史
- 收稿日期:2018-04-04
- 收修改稿日期:2018-05-21




2. 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071;
3. 江苏省海洋水产研究所 南通 226007;
4. 上海海洋大学 上海 201306
2. Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China;
3. Marine Fisheries Research Institute of Jiangsu, Nantong 226007, China;
4. Shanghai Ocean University, Shanghai 201306, China
近年来我国黄海海域连续多年暴发大规模的浒苔绿潮, 漂浮的浒苔在短短几个月之内生物量迅速增长, 海域覆盖面积呈倍数扩增(王宗灵等, 2018)。研究结果表明, 漂浮浒苔强大的繁殖能力是导致其生物量快速增长而引起绿潮暴发的主要原因之一(罗民波等, 2015)。据报道, 每平方厘米藻体能够产生大约5.35×106个游孢子或1.07×107个配子; 在繁殖高峰的5—8月间, 一株成熟藻体25%左右的部分能够放散生殖细胞, 产生约1.15×107个游孢子或2.31×107个配子(张华伟等, 2011)。浒苔繁殖方式多样, 包括有性生殖、无性生殖和营养繁殖等, 繁殖能力强, 在生活史周期中的任何一个中间形态都可以单独发育成为成熟藻体。其中, 海水中和潮间带底泥中含有大量的浒苔微观繁殖体, 包括浒苔孢子、配子、合子以及其发育不同程度的个体, 它们和浒苔藻体都具有较强的抗胁迫能力(Taylor et al, 2001)。
浒苔绿潮的连年发生, 给我国沿海环境、人居生活和渔业生产等带来极大影响(刘佳等, 2018), 寻求有效的浒苔防控途径显得极为迫切。对抗或抑制侵入性或有害物种的方法一般包括机械法、生物法、化学法、遗传法和环境控制(Allendorf et al, 2003)。近年来在我国对浒苔的治理上已规模化的使用了机械打捞法, 改性黏土治理也取得重要进展(李靖等, 2015;张悦等, 2016), 但治理效果依然有待进一步提高, 而对环境影响较小的生物防控技术是值得尝试的重要手段。
滤食性贝类是海洋生态系统中重要的生态类群, 利用鳃丝及其上着生的纤毛过滤、截留水体中的浮游植物、碎屑等颗粒物来获得营养来源。贝类发达的滤食系统使它具有强大的滤水能力, 据报道, 19—20.5℃条件下, 长牡蛎(Crassostrea gigas)的滤水率为1.97L/(ind·h) (组织干重Wd: 0.32g) (王俊等, 2000), 18℃下, 紫贻贝(Mytilus edulis)的滤水率分别为和1.3L/(ind·h) (Wd=0.028g) (Riisgård et al, 2003), 单个个体一天时间就分别可以过滤47.28L和31.2L的海水。对颗粒物摄食选择性的研究结果表明, 滤食性贝类能够有效地截留4μm以上的颗粒物(Barillé et al, 1993)。黄海浒苔绿潮从形成到消亡的全年过程可分为3个时期, 分别为微观繁殖体期(11—3月), 绿潮暴发和持续期(4—7月)、绿潮海藻消亡期(8—10月)(李曰嵩等, 2015)。研究表明, 浒苔的各种游动细胞大小分别为:雄配子长6.7±0.51μm, 宽2.5±0.34μm, 雌配子长7.4±0.48μm, 宽3.3±0.22μm, 游孢子11.7±0.73μm, 宽5.2±0.75μm (王晓坤等, 2007)。从浒苔的游动孢子和配子的粒径来看, 处于滤食性贝类有效截留的粒径范围内, 这为实施滤食性贝类对浒苔微观繁殖体阶段的生物防控策略提供了可能。长牡蛎和紫贻贝是我国重要的海水养殖经济贝类种类, 适应能力强, 分布广泛, 且生长速度快, 筏式吊养、滩涂平挂等养殖技术非常成熟, 可以作为实施浒苔微观繁殖体阶段生物防控的候选种类, 且研究表明, 海水水体中微观繁殖体的高值区主要集中在紫菜筏架养殖区附近(方松等, 2012), 因此我们开展了长牡蛎和紫贻贝对浒苔微观繁殖体的摄食实验, 并在江苏如东紫菜养殖区的滩涂进行了长牡蛎和紫贻贝的挂养实验, 旨在探究利用滤食性贝类进行浒苔防控的可行性, 为我国浒苔的治理提供新思路。
1 材料与方法 1.1 牡蛎和贻贝滤除浒苔微观繁殖体的室内受控实验 1.1.1 微观繁体的获得足量浒苔(取自江苏射阳河口)于室外帆布围隔内(3m×2m×1.0m)培养, 围隔设进水口和排水口, 水体约每1.5h更新一次, 浒苔培养4—7d, 挑选膨大藻体, 切成1—1.5cm的藻段, 用事先经0.45μm滤膜过滤过的海水冲洗多次, 最后置于0.45μm过滤的海水内, 待半数藻段孢子/配子放散, 制备为浒苔微繁体母液。为防止颗粒物浓度(微繁体)过高引起贝类的不充分摄食, 微繁体母液经0.45μm滤膜过滤过的海水稀释, 至终浓度约2.3—2.5×104 ind/mL。
1.1.2 贝类滤食的测定以事先清洗干净并驯化3—4d的长牡蛎和紫贻贝(取自青岛团岛湾)为实验对象, 并在实验前一天置于0.45μm过滤的海水内, 使其排空肠道。实验采用静水法, 每组1个长牡蛎或4个紫贻贝(n=4—6) (长牡蛎的壳长和干重分别为43.1±2.2mm, 146.7±8.5mg; 紫贻贝的壳长和干重分别为25.6±3.29mm, 67.7±5.3mg), 实验水体为稀释好的微繁体液, 每组水体体积7.5L, 对照组不放置贝类(n=3), 每间隔1h, 以便携式颗粒计数器PAMAS (型号S4031GO, 德国)测定水体2—30μm颗粒物的数量。实验持续2h, 期间充气。2h内长牡蛎和紫贻贝对不同大小微观繁殖体的截留率(RP)计算如下:
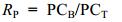 (1)
(1)式中, PCB为贝类在第1h或第2h对不同粒径的摄食数量, PCT为该时间段所摄食的颗粒总数。
1.1.3 贝类对微观繁殖体的有效滤除率的测定与计算贝类滤食前和滤食后的微繁体差值为被贝类滤除的微繁体, 有两种可能的存在形式, 一是不被贝类消化, 存在于假粪中; 二是被部分/全部消化, 存在于真粪中。而真粪和假粪中微繁体的萌发活性可能保留也可能丧失, 因此实验结束后, 将贝类置于0.45μm滤膜过滤的海水中, 在接下来的24h内, 以吸管虹吸真粪, 以200μm筛绢收集所有假粪, 并将假粪均分4份, 于三角锥形瓶内, 参考刘峰等(2010)的方法, 添加饱和二氧化锗(GeO2)水溶液和f/2培养液工作液于光照培养箱内培养2周, 计真粪和假粪中萌发的浒苔微观繁殖体数量。同时对照组水体与贝类摄食后的水体进行培养, 计数微繁体数量。实验以单位时间内被单位个体(重量)的贝类有效滤食的微繁体数量作为贝类对微繁体的有效滤除率RF(%)。
主要计算公式如下:
 (2)
(2)Ns为贝类滤食实验组微繁体萌发数量(ind), Nc为对照组微繁体萌发数量(ind); T为实验持续时间(h)。
 (3)
(3)Nw为贝类滤食后单位水体微繁体萌发数量, V为实验水体总体积(l), Nf为粪便和假粪中萌发的微繁体数量(ind)。
 (4)
(4)Nc为对照组微繁体萌发数量, Ncw为对照组单位水体微繁体萌发数量。
1.2 紫菜养殖海区牡蛎和贻贝的挂养实验2016年5—7月在南通如东紫菜养殖区进行平挂式养殖实验设施的布设, 平挂方式采用竹竿搭架式, 每排筏架总长约60m, 宽度2m, 每2m放置一横竹竿, 形成2m×2m的小区, 牡蛎和贻贝苗绳顺筏架方向, 系于横竹竿上。2016年11月, 运送长牡蛎、紫贻贝苗种各100绳进行养殖, 并针对次年2—5月份的浒苔形成期, 对养殖的长牡蛎和紫贻贝的生长情况进行跟踪调查。
1.3 挂养长牡蛎和紫贻贝对浒苔微观繁殖体的控制能力评估根据养殖期间长牡蛎和紫贻贝的大小, 参考牛亚丽(2014)对长牡蛎和Wang等(2015)对紫贻贝的滤水率的研究结果(具有相同规格与相近水温及尽可能一致的地理位置)、对微观繁殖体的有效滤除率及江苏如东海区水体微观繁殖体浓度(方松等, 2012), 计算挂养长牡蛎和紫贻贝对微观繁殖体的控制能力。
2 结果 2.1 长牡蛎和紫贻贝对浒苔微观繁殖体的滤除制备得到的实验用微观繁殖体水体中, 微观繁殖体粒径多为5—12μm(图 1a, 图 1b), 该范围个体约占总数的89.5%—89.8%, 其中以8—9μm的个体数量最多, 约占总颗粒数的28.8%—30.7%。长牡蛎和紫贻贝对水体2—30μm的颗粒均可截留, 但不同粒径颗粒的截留率不同, 2—3μm颗粒的截留率最小。2h的摄食实验中长牡蛎和紫贻贝分别滤除了总颗粒的56.4%和50.9%, 平均每小时每个个体的截留速率为6892ind/(ind·h)和1589ind/(ind·h)。
 |
| 图 1 长牡蛎(a)和紫贻贝(b)摄食前后微观繁殖体颗粒浓度的变化 Fig. 1 Changes in microscopic propagules particle concentration before and after feeding for oyster C. gigas (a) and mussel M. edulis (b) |
 |
| 图 2 长牡蛎(a)和紫贻贝(b)摄食前各颗粒所占比重及摄食各颗粒占总摄食颗粒的比重 Fig. 2 Percentages of particles before feeding and during feeding for oyster C. gigas (a) and mussel M. edulis (b) |
 |
| 图 3 长牡蛎和紫贻贝滤食后水体和粪便中萌发的微观繁殖体比重 Fig. 3 Percentages of the germinated microscopic propagules in the filtered water and faces of oyster C. gigas and mussel M. edulis |
 |
| 图 4 2—5月份挂养长牡蛎和紫贻贝的壳长(a)及湿重(b)的增长 Fig. 4 Growth of shell length (a) and wet weight (b) of oyster C. gigas and mussel M. edulis February to May |
对摄食前后的水体进行培养的结果表明, 牡蛎滤食后的单位水体中, 萌发的微观繁殖体数为对照的43.8%, 即对微繁体的有效滤除率为56.2%, 紫贻贝滤食后的水体萌发的微观繁殖体为对照组的51.6%, 对微繁体的滤除率为48.4%。牡蛎和贻贝的真粪中未见萌发的微繁体, 但产生的假粪中萌发的微繁体较多, 分别为对照组总萌发数量的23.6%和30.4%, 对总的萌发微繁体的统计表明, 2h的摄食实验中, 牡蛎和贻贝可有效滤除总的微观繁殖体的32.6%和18.0%, 平均每小时每个个体的有效滤除率为16.3%和2.3%, 有效滤除速率为3980 ind/(ind·h)和574 ind/(ind·h)。
2.2 紫菜养殖海区牡蛎和贻贝的挂养实验至2017年6月份, 牡蛎和贻贝的成活率近100%。在2—5月份浒苔暴发展期间, 长牡蛎平均壳长由34.5mm生长至40.3mm, 紫贻贝的壳长由18.0mm增至21.9mm; 至5月底, 两种类的平均湿重分别为7.96g和1.18g。
2.3 挂养长牡蛎和紫贻贝对浒苔微观繁殖体的控制能力评估根据挂养长牡蛎和紫贻贝的数量及滤水率, 计算得到每绳长牡蛎每天滤水量为139.6L, 紫贻贝为908.6L, 挂养的100绳牡蛎和紫贻贝总计每天滤水量为1.05×105L。参考方松等(2012)调查得到的3—5月份江苏沿岸水体微观繁殖体数量, 挂养的牡蛎和贻贝每天可有效滤除的微观繁殖体个数约6.37×105个。
| 每绳的个数(ind) | 3—5月个体 平均干重(g) |
滤水率 (L/(g h)) |
有效滤除率(%/(ind·h)) | 水体微观繁殖体数量(ind) | 每绳每小时 滤水量(L) |
每绳每天滤 水量(L) |
100绳总计滤 水量(L) |
滤除微繁体数(ind) | |
| 长牡蛎 | 102.6 | 0.135 | 0.42 | 16.3 | 146 | 5.8 | 139.6 | 13961.8 | 332263.1 |
| 紫贻贝 | 932.5 | 0.058 | 0.7 | 2.3 | 146 | 37.9 | 908.6 | 90862.8 | 305117.3 |
| 总计 | 1048.2 | 104824.6 | 637380.4 |
我们的滤食实验证实, 牡蛎和贻贝可有效截留微观繁殖体。在制备的微观繁殖体实验液中, 5— 12μm的颗粒约占总颗粒数的90%, 这与王晓坤等(2007)观察到的浒苔微观繁殖体粒径范围一致。实验还表明该范围内(5—12μm)的颗粒可被长牡蛎和紫贻贝有效截留, 这亦在Strohmeier等(2012)的研究中得到体现, 该研究观察到紫贻贝的最小有效截留粒径为4μm, 最大截留粒径约30—35μm; 在对维吉尼亚美洲牡蛎(C. virginica)的研究中, 该种类可以完全过滤大于5μm的颗粒。滤食性贝类摄食食物颗粒的第一步是有效过滤, 即通过其鳃丝和其上着生的纤毛的组合运动完成对适合颗粒的优先摄取, 由于不同种类前侧纤毛的复杂程度及栉鳃形式的不同, 对不同粒径颗粒的有效截留率也不同, 通常双壳贝类的摄食粒径有一定的分布范围, 因此过大或过小均不能被有效截留(Møhlenberg et al, 1978; Riisgård, 1988; Jørgensen, 1989)。同时, 对摄食后的微观繁殖体水体进行的培养亦表明, 萌发的微观繁殖体数为对照的43.8%—49.7%, 即进一步证实, 长牡蛎和紫贻贝对微繁体可以有效滤除, 滤除率为50.3%—56.2%。
除了通过第一步的有效过滤实现对某些颗粒截留, 贝类随后会继续通过栉鳃对颗粒的进行选择及对某些颗粒的优先输送, 将颗粒运送至唇瓣; 唇瓣亦具有对食物的选择性和摄取性, 优先性摄取某些颗粒, 将颗粒传送至胃, 胃部还会对食物进行进一步选择, 将某些颗粒优先输送至消化盲囊, 进行消化(Ward et al, 2004)。长牡蛎和紫贻贝对浒苔微观繁殖体颗粒的截留并不意味着可以经过随后的输送, 进行完全的消化吸收, 因此, 我们对假粪和粪便进行的培养表明, 部分微观繁殖体仍保留活性, 存在于假粪中, 培养后可以萌发, 但是部分已经过消化存在于真粪中, 丧失了萌发的能力。我们的研究中, 紫贻贝假粪中萌发的微观繁殖体比例高于长牡蛎, 这种种属差异可能与滤食后存在于唇瓣及胃的颗粒选择性有关。
虽然颗粒大小是影响贝类捕获效率的重要因素, 实验中长牡蛎和紫贻贝可有效滤除浒苔微观繁殖体, 但是颗粒本身的理化性质亦影响贝类对颗粒的捕获, 例如Shumway等人(1985)的研究表明, 与硅藻三角褐指藻和鞭毛虫Chroomonas salina相比, 欧洲牡蛎Ostrea edulis, 优先捕获腰鞭毛虫Prorocentrum minimum。而Rosa等(2017)的研究表明, 微藻表面的理化特性影响牡蛎和贻贝对9种不同微藻的摄取, 凝集素亲和力和润湿性及表面电荷是影响两种贝类摄食选择性的重要因素。因此, 我们的实验虽然证实长牡蛎和紫贻贝可以有效捕获并部分的消化吸收浒苔微观繁殖体, 但实验所用的微观繁殖体几乎为纯的微繁体液, 而在自然海域, 浒苔微观繁殖体与种类繁多的浮游动植物颗粒并存, 贝类是否会对浒苔微观繁殖体存在优先捕获或选择性不捕获, 需进一步研究。
生物法历来为对抗或抑制有害物种的重要方法。已有研究表明, 利用滤食性贝类、挠足类及纤毛类控制赤潮藻的发展是值得推广和发展的赤潮治理方法, 而对于有毒赤潮, 该方式则因毒素会在食物链富集而不建议采用。由于浒苔不含毒素, 因此利用滤食性贝类防控浒苔具有一定的可行性。本研究在江苏如东紫菜养殖区进行的挂养实验表明, 挂养的长牡蛎和紫贻贝可以存活并生长, 但7—9月份高温季节遭遇了藤壶在长牡蛎壳上的大量附着现象, 后续将尝试采用“11月份养殖大规格苗种, 翌年7月份之前收获”的策略避开藤壶附着高峰期, 达到生物防控效果的同时, 获得一定的经济效益。此外, 后续研究将继续探究具有丰富资源量的蛤、蛏、蚶等滩涂贝类对浒苔微观繁殖体的摄食和防控能力。
4 结论长牡蛎和紫贻贝可有效截留浒苔微观繁殖体, 并经消化吸收使得部分微观繁殖体失去萌发活力, 从而证实两种贝类对防控浒苔微观繁殖体的能力; 在江苏紫菜养殖区进行的平挂式养殖初步成功, 浒苔发生期, 挂养的长牡蛎和紫贻贝每天可滤食微观繁殖体约63.7万个。对养殖方式的进一步优化改进, 以及研究和发展江苏浅滩其他滩涂贝类对浒苔的防控, 亟需进一步的深入开展。
| 方松, 王宗灵, 李艳, 等, 2012. 南黄海及长江口附近海域绿潮暴发前期微观繁殖体的动态变化. 海洋学报, 34(4): 147–154 |
| 牛亚丽, 2014.桑沟湾滤食性贝类碳、氮、磷, 硅元素收支的季节变化研究.舟山: 浙江海洋学院硕士学位论文 http://cdmd.cnki.com.cn/article/cdmd-10340-1014219529.htm |
| 王俊, 姜祖辉, 张波, 等, 2000. 太平洋牡蛎同化率的研究. 应用生态学报, 11(3): 441–444 DOI:10.3321/j.issn:1001-9332.2000.03.028 |
| 王宗灵, 傅明珠, 肖洁, 等, 2018. 黄海浒苔绿潮研究进展. 海洋学报, 40(2): 1–13 |
| 王晓坤, 马家海, 叶道才, 等, 2007. 浒苔(Enteromorpha prolifera)生活史的初步研究. 海洋通报, 26(5): 112–116 DOI:10.3969/j.issn.1001-6392.2007.05.018 |
| 刘佳, 刘宁, 2018. 浒苔绿潮影响下滨海旅游环境价值损失及影响因素——以青岛市海水浴场为例. 资源科学, 40(2): 392–403 |
| 刘峰, 逄少军, 单体锋, 等, 2010. 一种新的海水中石莼属海藻显微阶段个体数定量方法及在黄海绿潮爆发过程中的应用. 科学通报, 55(6): 468–473 |
| 张悦, 宋秀贤, 李靖, 等, 2016. 不同体系改性粘土对浒苔(Ulva prolifera)微观繁殖体去除及萌发的影响. 海洋学报, 38(8): 93–102 DOI:10.3969/j.issn.0253-4193.2016.08.010 |
| 张华伟, 马家海, 胡翔, 2011. 绿潮漂浮浒苔繁殖特性的研究. 上海海洋大学学报, 20(4): 600–606 |
| 李靖, 孙雷, 宋秀贤, 等, 2015. 改性粘土对浒苔(Ulva prolifera)微观繁殖体去除效果及萌发的影响. 海洋与湖沼, 46(2): 345–350 |
| 李曰嵩, 肖文军, 杨红, 等, 2015. 2012年黄海绿潮藻早期发生和聚集动力学成因分析. 海洋环境科学, 34(2): 268–273 |
| 罗民波, 刘峰, 2015. 南黄海浒苔绿潮的发生过程及关键要素研究进展. 海洋渔业, 37(6): 570–574 DOI:10.3969/j.issn.1004-2490.2015.06.013 |
| Allendorf F W, Lundquist L L, 2003. Introduction:population biology, evolution, and control of invasive species. Conservation Biology, 17(1): 24–30 DOI:10.1046/j.1523-1739.2003.02365.x |
| Barillé L, Prou J, Héral M, et al, 1993. No influence of food quality, but ration-dependent retention efficiencies in the Japanese oyster Crassostrea gigas. Journal of Experimental Marine Biology and Ecology, 171(1): 91–106 DOI:10.1016/0022-0981(93)90142-B |
| Jørgensen C B, 1989. Water processing in ciliary feeders, with special reference to the bivalve filter pump. Comparative Biochemistry and Physiology Part A:Physiology, 94(2): 383–394 |
| Møhlenberg F, Riisgård H U, 1978. Efficiency of particle retention in 13 species of suspension feeding bivalves. Ophelia, 17(2): 239–246 DOI:10.1080/00785326.1978.10425487 |
| Riisgård H U, 1988. Efficiency of particle retention and filtration rate in 6 species of Northeast American bivalves. Marine Ecology Progress Series, 45: 217–223 DOI:10.3354/meps045217 |
| Riisgård H U, Kittner C, Seerup D F, 2003. Regulation of opening state and filtration rate in filter-feeding bivalves (Cardium edule, Mytilus edulis, Mya arenaria) in response to low algal concentration. Journal of Experimental Marine Biology and Ecology, 284(1-2): 105–127 DOI:10.1016/S0022-0981(02)00496-3 |
| Rosa M, Ward J E, Holohan B A, et al, 2017. Physicochemical surface properties of microalgae and their combined effects on particle selection by suspension-feeding bivalve molluscs. Journal of Experimental Marine Biology and Ecology, 486: 59–68 DOI:10.1016/j.jembe.2016.09.007 |
| Shumway S E, Cucci T L, Newell R C, et al, 1985. Particle selection, ingestion, and absorption in filter-feeding bivalves. Journal of Experimental Marine Biology and Ecology, 91(1-2): 77–92 DOI:10.1016/0022-0981(85)90222-9 |
| Strohmeier T, Strand Ø, Alunno-Bruscia M, et al, 2012. Variability in particle retention efficiency by the mussel Mytilus edulis. Journal of Experimental Marine Biology and Ecology, 412: 96–102 DOI:10.1016/j.jembe.2011.11.006 |
| Taylor R, Fletcher R L, Raven J A, 2001. Preliminary studies on the growth of selected 'green tide' algae in laboratory culture:effects of irradiance, temperature, salinity and nutrients on growth rate. Botanica Marina, 44(4): 327–336 |
| Wang Y J, Li L S, Hu M H, et al, 2015. Physiological energetics of the thick shell mussel Mytilus coruscus exposed to seawater acidification and thermal stress. Science of the Total Environment, 514: 261–272 DOI:10.1016/j.scitotenv.2015.01.092 |
| Ward J E, Shumway S E, 2004. Separating the grain from the chaff:particle selection in suspension-and deposit-feeding bivalves. Journal of Experimental Marine Biology and Ecology, 300(1-2): 83–130 DOI:10.1016/j.jembe.2004.03.002 |