 2018, Vol. 49
2018, Vol. 49中国海洋湖沼学会主办。
文章信息
- 孙鹏, 王咏雪, 田阔, 求锦津, 张苗苗, 聂振林, 田嘉琦, 水柏年. 2018.
- SUN Peng, WANG Yong-Xue, TIAN Kuo, QIU Jin-Jin, ZHANG Miao-Miao, NIE Zhen-Lin, TIAN Jia-Qi, SHUI Bo-Nian. 2018.
- 浙江5个海湾鱼类分类多样性研究
- TAXONOMIC DIVERSITY OF FISH SPECIES IN FIVE BAYS IN ZHEJIANG PROVINCE
- 海洋与湖沼, 49(6): 1325-1333
- Oceanologia et Limnologia Sinica, 49(6): 1325-1333.
- http://dx.doi.org/10.11693/hyhz20180100019
-
文章历史
- 收稿日期:2018-01-26
- 收修改稿日期:2018-06-28




2. 台州市海洋与渔业局 台州 318000
2. Taizhou Bureau of Oceanology and Fishery, Taizhou 318000, China
生物多样性的含义十分广泛, 不仅包括物种的多样性, 还包括物种间进化关系、进化历史以及生态系统等各层面的多样性(曲方圆等, 2010)。常用的生物多样性指数如Shannon Wiener多样性指数、Pielous均匀度指数及Margalef物种丰富度指数等在研究鱼类群落结构及其变化方面有着广泛的应用。但它们受取样时间、方法及样本大小的影响较大, 且仅能反映该生物群落多样性的总体水平, 具有一定的不确定性。为研究分类学层面的生物多样性, 国外有学者提出了两个生物分类多样性的概念, 即平均分类差异指数Δ+和分类差异变异指数Λ+, 用于分析物种在分类树上分布的均匀程度(Clarke et al, 1998a, b)。蒋志刚等(1999)提出了G-F指数的概念用于科级、属级水平上多样性的分析。这两种指数仅根据研究地域物种的有无便可对该地生物多样性进行分析, 对开展不同区域、不同生境间和不同历史时期的对比研究, 以及取样性质未知或不一致情况下的生物多样性研究具有重要意义。近年来, 分类多样性的概念被国内一些学者的应用引起了众多学者的关注, 对中国海域的鱼类分类多样性做过相关的研究(徐宾铎等, 2007; 李娜娜等, 2011; 林龙山等, 2012; 胡成业等, 2015)。
随着浙江沿海地区经济的迅速发展, 沿岸海域的渔业资源所受的人为扰动因素加大, 出现了渔业资源衰退、生物多样性下降等现象。目前, 有关浙江沿岸海域分类多样性的研究仅见于底栖生物(胡成业等, 2016; 求锦津等, 2017), 而鱼类多样性的研究主要集中在群落多样性层面上, 关于分类多样性的研究及报道较为罕见。本研究所选取的5个海湾(杭州湾、象山湾、三门湾、台州湾与乐清湾)同为半封闭型海湾, 底质均为泥质, 水体受台湾暖流和浙闽沿岸流影响, 自然环境相差不大。通过对浙江5个海湾鱼类分类多样性的分析对比, 揭示其多样性特征及差异, 以期为鱼类资源的开发、利用、保护和管理等应用研究提供科学依据。
1 材料与方法 1.1 研究区域浙江海岸线蜿蜒曲折, 海湾众多, 本研究以三门湾调查数据为基准, 又选取了相似的的杭州湾、象山湾、台州湾与乐清湾作为研究对象。其研究区域图如图 1所示。
 |
| 图 1 研究区域 Fig. 1 The study area |
本研究基于2014年11月(秋季)和2015年5月(春季)在三门湾开展的渔业资源调查数据, 结合杭州湾(沈新强等, 2011; 王淼等, 2013, 2015, 2016; 谢旭等, 2013)、象山湾(象山县海洋与渔业局等, 2008; 姜亚洲等, 2014; 郭小雨等, 2015)、三门湾(邵凡户, 1994; 蔡萌等, 2009; 李新等, 2014; 冀萌萌等, 2016)、台州湾(齐海明等, 2014)、乐清湾(乐清水产局, 1999; 闫丽娜等, 2013; 夏陆军等, 2016)近20年左右的相关资料, 另外台州湾的相关数据还引用了中国水产科学研究院东海水产研究所编制的《台州港临海港区头门作业区港池试挖及北洋涂涂面整理工程环境影响报告书》、浙江省海洋生态环境科学研究所编制的《临海市国有资产投资控股有限公司年产150万台铝热交换器项目填海工程建设项目环境影响报告表》及浙江大学编制的《临海市2016-2出让海域填海工程环境影响评价专题报告》、《台州中心港区(临海)疏港公路一期白沙至头门段工程(港区产业城段)环境影响报告》中的相关资料。依据《中国海洋生物名录》系统整理鱼类种类及其相应的分类阶元, 排除同种异名, 构建5个海湾鱼种名录。
1.3 分类多样性指数本研究采用平均分类差异指数Δ+与分类差异变异指数Λ+ (Warwick et al, 1995; Clarke et al, 1998a, b)两个指数体现5个海湾鱼类的分类多样性。
平均分类差异指数(average taxonomic distinctne ss index):
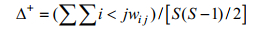 (1)
(1)分类差异变异指数(variation in taxonomic distinctness index):
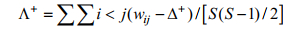 (2)
(2)式(1)、(2)中, ωij为第i和j个种类在分类系统树中的分支路径长度, S为种类数。平均分类差异指数Δ+表示群落中所有物种之间分类路径长度的平均值, 分类差异变异指数Λ+就是与平均分类差异指数Δ+偏离程度的理论平均值。由于所研究的鱼类均属于脊索动物门、脊椎动物亚门的软骨鱼纲与硬骨鱼纲, 本研究涉及的分类阶元定为种、属、科、目、纲5个级别, 5个分类阶元水平的多样性权重值见表 1。平均分类差异指数Δ+、分类差异变异指数Λ+及95%置信漏斗区间由PRIMER5.2软件包中的TAXDTEST求得。
| 分类等级 | 路径长度的权重ωij | ||||
| 杭州湾 | 象山湾 | 三门湾 | 台州湾 | 乐清湾 | |
| 种 | 5.84 | 4.32 | 9.16 | 7.09 | 9.17 |
| 属 | 23.88 | 17.19 | 22.7 | 23.7 | 22.01 |
| 科 | 36.25 | 36.42 | 42.73 | 42.26 | 42.22 |
| 目 | 64.93 | 66.34 | 68.18 | 67.52 | 67.9 |
| 纲 | 100 | 100 | 100 | 100 | 100 |
为表明不同分类阶元上鱼类组成的多样性, 本研究采用分类阶元包含指数TINCLi (李娜娜等, 2011)进行分析。
分类阶元包含指数:
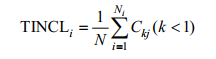 (3)
(3)式(3)中, Ni表示第i级分类阶元的数目, Ckj为第j个k级分类阶元的数目。TINCLi值越小, 则在上一级分类阶元所包含的下一级阶元的均值越小, 表示鱼类在该分类阶元的种类分布越不集中, 亲缘关系较远, 相应分类多样性也越高(李娜娜等, 2011)。
1.5 G-F多样性指数本研究采用G-F指数(蒋志刚等, 1999)来分析不同海湾在科、属水平上的分类多样性。
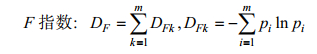 (4)
(4)式(4)中, pi=Ski/Sk, Ski为名录中k科i属中的物种数, Sk为名录中k科中的物种数, n为k科中的属数, m为鱼类的科数。
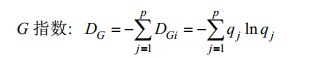 (5)
(5)式(5)中, qj=Sj/S, Sj为名录中j属中的物种数, S为名录中的物种数, p为名录中的属数。
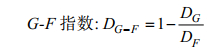 (6)
(6)一般而言, 研究区域单种科越多, G-F指数值越小, 反之则越大。当DF=0, 即所有的科均为单种时, 则该地区的G-F指数为0。G-F指数越接近0或为负数, 则说明F指数(科间多样性)下降, 或者说明G指数(属间多样性)上升, 反之则趋近于1。
1.6 相似性系数为比较5个海湾之间鱼类种类的差异, 本研究采用相似性系数来计算5个海湾之间鱼类目级和科级分类阶元的相似性系数。
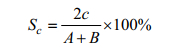 (7)
(7)式中, A为甲所拥有的鱼类目(科)数, B为乙所拥有的鱼类目(科)数, c为两者共有目(科)数, Sc为相似性指数, 其值为0≤Sc≤1。Sc越大, 表明两个海域间鱼类种类相似度越高。
2 结果与分析 2.1 种类组成在5个海湾中, 象山湾鱼种数最多(78种), 其后依次为三门湾(66种), 乐清湾(63种), 台州湾(55种), 杭州湾(42种), 其具体分类阶元数见图 2。在目级分类阶元, 5个海湾均为鲈形目(Perciformes)鱼种数最多, 即依次为杭州湾21种、象山湾38种、三门湾27种、台州湾29种及乐清湾32种。在科级分类阶元, 5个海湾石首鱼科(Sciaenidae)、
 |
| 图 2 5个海湾鱼类不同分类阶元数 Fig. 2 The number of different taxonomic categories of fish in the five bays |
根据本文构建的鱼类物种名录及多样性权重值, 求得5个海湾的Δ+和Λ+(表 3, 图 3, 图 4)。其中, 平均分类差异指数Δ+值大小排序为象山湾(56.01) > 乐清湾(55.17) > 台州湾(50.08) > 杭州湾(49.62) > 三门湾(48.79);分类差异变异指数Λ+值大小排序为象山湾(553.83) > 乐清湾(394.64) > 杭州湾(363.87) > 台州湾(327.21) > 三门湾(313.63)。
| 区域 | 平均分类差异指数Δ+ | 分类变异差异指数Λ+ |
| 杭州湾 | 49.62 | 363.87 |
| 象山湾 | 56.01 | 553.83 |
| 三门湾 | 48.79 | 313.63 |
| 台州湾 | 50.08 | 327.21 |
| 乐清湾 | 55.17 | 394.64 |
 |
| 图 3 平均分类差异指数 Fig. 3 The average taxonomic distinctness |
 |
| 图 4 分类变异差异指数 Fig. 4 Variation in taxonomic distinctness |
应用分类阶元包含指数TINCLi分析5个海湾鱼类组成, 结果如表 4显示。三门湾鱼类的科、属、种的分类阶元包含指数值均较高, 其鱼类组成目、科、属所分别拥有的(科、属、种)、(属、种)和(种)的平均值依次为(2.7, 4.7, 6.6)、(1.74, 2.44)和(1.41)。在目级水平, 杭州湾鱼类分类阶元包含指数最低, 目所拥有的(科、属、种)的平均值为(1.55, 3.18, 3.82), 在科、属级水平, 象山湾鱼类分类阶元包含指数最低, 其科、属所分别拥有的(属、种)和(种)的平均值分别为(1.62, 1.86)和(1.15)。
| 纲 | 目 | 科 | 物种数 | ||||
| HZB | XSB | SMB | TZB | YQB | |||
| 软骨鱼纲Chondrichthyes | 鲼形目Myliobatiformes | 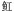 科Dasyatidae 科Dasyatidae |
1 | 1 | 1 | 3 | |
| 鳐形目Rajiformes | 团扇鳐科Platyrhinidae | 1 | |||||
| 鳐科Rajidae | 2 | ||||||
| 电鳐目Torpediniformes | 电鳐科Torpedinidae | 1 | |||||
| 真鲨目Carcharhiniformes | 真鲨科Carcharhinidae | 1 | |||||
| 角鲨目Squalusformes | 角鲨科Squalidae | 1 | |||||
| 六鳃鲨目exanchiformes | 六鳃鲨科Hexanchiformes | 1 | |||||
| 硬骨鱼纲Osteichthyes | 鲈形目Perciformes | 鲷科Sparidae | 2 | 1 | 2 | ||
| 发光鲷科Acropomatidae | 1 | ||||||
| 长鲳科Centrolophidae | 1 | ||||||
| 塘鳢鱼科Eleotridae | 1 | 1 | |||||
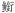 科Engraulidae 科Engraulidae |
1 | 1 | |||||
| 虎鱼科Gobiidae |
10 | 9 | 7 | 7 | 5 | ||
| 石首鱼科Sciaenidae | 6 | 8 | 12 | 10 | 8 | ||
| 羊鱼科Mullidae | 1 | 1 | |||||
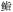 科Serranidae 科Serranidae |
1 | 2 | 3 | 2 | 4 | ||
| 鲭科Scombrida | 1 | 2 | 2 | 1 | |||
| 鲳科Stromateidae | 2 | 1 | 4 | 4 | 3 | ||
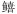 科Sillaginidae 科Sillaginidae |
1 | 1 | 1 | ||||
| 鳢科Channidae | 1 | ||||||
| 带鱼科Trichiuridae | 1 | 1 | 2 | 1 | 1 | ||
| 鲹科Carangidae | 2 | 1 | |||||
| 鳄齿鱼科hampsodontidae | 1 | ||||||
| 石鲈科Pomadasyidae | 1 | 2 | 2 | ||||
| 天竺鲷科Apogonidae | 1 | ||||||
| 鲱形目Clupeitormes | 鲱科Clupeidae | 4 | 3 | 2 | 4 | ||
| 鳀科Engraulidae | 6 | 7 | 5 | 6 | 6 | ||
| 鲻形目Mugiliformes | 鲻科Mugilidae | 1 | 1 | 2 | 2 | 4 | |
| 马鲅科Polynemidae | 1 | 1 | 2 | 2 | 1 | ||
| 鲽形目Pleuronectiformes | 舌鳎科Cynoglossidae | 4 | 4 | 5 | 4 | 6 | |
| 鳎科Soleidae | 1 | ||||||
| 鲆科Bothidae | 1 | ||||||
| 牙鲆科Paralichthyidae | 1 | ||||||
| 鲽科Pleuronectidae | 1 | ||||||
| 海鲢目Elopiformes | 海鲢科Elopidae | 1 | |||||
| 鲀形目Tetraodontiformes | 单角鲀科Monacanthidae | 2 | 1 | ||||
| 鲀科Tetraodontidae | 2 | 3 | 4 | 2 | |||
| 翻车鲀科Molidae | 1 | ||||||
| 刺鲀科Diodontidae | 1 | ||||||
| 鲉形目Scorpaeniformes | 鲂 科Triglidae 科Triglidae |
1 | 2 | 1 | |||
| 鲉科Scorpaenidae | 1 | ||||||
| 毒鲉科Synanceiidae | 1 | ||||||
| 鲬科Platycephalidae | 1 | 1 | |||||
| 鳕形目Gadiformes | 犀鳕科Bregmacerotidae | 1 | |||||
| 鲇形目Siluriformes | 海鲶科Ariidae | 1 | 1 | 1 | |||
| 灯笼鱼目Myctophiformes | 灯笼鱼科Myctophidae | 1 | 1 | ||||
| 狗母鱼科Synodontidae | 2 | 2 | 1 | 2 | 1 | ||
| 海龙目Syngnathiformes | 海龙科Syngnathidae | 1 | 1 | 1 | |||
| 鳗鲡目Anguilliformus | 海鳗科Muraenesocidae | 1 | 1 | 1 | 1 | 1 | |
| 蛇鳗科Ophichthyidae | 1 | ||||||
| 康吉鳗科Congridae | 1 | 1 | |||||
| 海鳝科Muraenidae | 1 | ||||||
| 鼠目onorhynchiformes |
遮目鱼科Chanoidae | 1 | |||||
 目Lophiiformes 目Lophiiformes |
科Lophiidae |
1 | 1 | ||||
| 鲑形目Salmoniformes | 银鱼科Salangidae | 1 | 1 | ||||
| 总计 | 42 | 78 | 66 | 55 | 63 | ||
| 注:表中HZW为杭州湾, XSW为象山湾, SMW为三门湾, TZW为台州湾, YQW为乐清湾 | |||||||
| 区域 | 物种数 | 种/属(S/G) | 种/科(S/F) | 种/目(S/O) | 属/科(G/F) | 属/目(G/O) | 科/目(F/O) |
| 杭州湾 | 42 | 1.2 | 2.47 | 3.82 | 2.06 | 3.18 | 1.55 |
| 象山湾 | 78 | 1.15 | 1.86 | 4.33 | 1.62 | 3.78 | 2.33 |
| 三门湾 | 66 | 1.41 | 2.44 | 6.6 | 1.74 | 4.7 | 2.7 |
| 台州湾 | 55 | 1.28 | 2.62 | 6.11 | 2.05 | 4.78 | 2.33 |
| 乐清湾 | 63 | 1.4 | 2.33 | 6.3 | 1.67 | 4.5 | 2.7 |
| 注:表中S、G、F和O分别代表种、属、科和目。下同 | |||||||
G-F多样性指数分析结果(表 5)表明: G指数大小排序为象山湾(4.12) > 台州湾(3.84) > 三门湾(3.72) > 乐清湾(3.63) > 杭州湾(3.47);F指数大小排序为象山湾(10.85) > 台州湾(9.23) > 三门湾(8.06) > 乐清湾(7.56) > 杭州湾(6.12); G-F指数大小排序为象山湾(0.62) > 台州湾(0.58) > 三门湾(0.54) > 乐清湾(0.52) > 杭州湾(0.43)。三个指数均为象山湾最高, 杭州湾最低。
| 区域 | 物种数 | G指数 | F指数 | G-F指数 |
| 杭州湾 | 42 | 3.47 | 6.12 | 0.43 |
| 象山湾 | 78 | 4.12 | 10.85 | 0.62 |
| 三门湾 | 66 | 3.72 | 8.06 | 0.54 |
| 台州湾 | 55 | 3.84 | 9.23 | 0.58 |
| 乐清湾 | 63 | 3.63 | 7.56 | 0.52 |
将5个海湾两两对比, 求得它们之间的鱼类相似性系数(表 6)。在目级水平台州湾与乐清湾之间相似性最高(0.84), 三门湾与象山湾相似性最低(0.5)。在科级水平台州湾与乐清湾之间相似性最高(0.71), 象山湾与台州湾相似性最低(0.54)。
| 区域 | 杭州湾(目/科) | 象山湾(目/科) | 三门湾(目/科) | 台州湾(目/科) |
| 象山湾 | 0.69/0.54 | — | — | — |
| 三门湾 | 0.67/0.64 | 0.5/0.58 | — | — |
| 台州湾 | 0.70/0.68 | 0.67/0.54 | 0.53/0.63 | — |
| 乐清湾 | 0.8/0.64 | 0.64/0.55 | 0.70/0.56 | 0.84/0.71 |
平均分类差异指数和分类差异变异指数是基于分类阶元间权重, 根据种类间分类等级的路径长度, 将群落分类多样性特征量化的指标体现(Rogers et al, 1999; Ceschia et al, 2007)。三门湾鱼类种间的亲缘关系最近, 分类多样性最低, 而象山湾鱼类种间的亲缘关系最远, 分类多样性最高。这可能是多种原因所致:首先, 鱼类的地理分布受诸多环境因子的影响, 象山湾岸线曲折, 海底地形复杂, 能够形成复杂的水文环境, 为分类多样性较高的鱼类分布提供了条件。同时, 象山湾内水体交换方式主要是湾外海水由下层入侵, 再涌升随落潮流由上层外泄(李家芳等, 1985), 可以形成上升流, 将海底丰富的营养盐带到表层, 且湾内水体较其他四个海湾清澈, 有利于浮游生物的繁衍, 为鱼类的生息繁衍提供丰富的饵料, 这些复杂的地形及水文等因素可能导致象山湾的鱼类的多样性较高。其次, 5个海湾附近地区产业结构存在一定差异, 导致人类活动对海洋环境的扰动不同而产生环境异质性。最后, 象山湾比邻渔业资源丰富的舟山渔场, 湾内与舟山渔场鱼类群体相互交流较方便, 这也可能是象山湾内鱼类多样性最高的原因之一。另外, 5个海湾Δ+值均低于东海大陆架区域(65.69) (李圣法, 2005), 即鱼类分类多样性均低于东海大陆架区, 这可能与研究区域范围窄, 生态环境性质较陆架区域相对单一有关。
象山湾分类差异变异指数(Λ+)值最高(553.83), 三门湾湾最低(313.63)。这说明象山湾鱼类在各分类阶元亲缘关系分布最不均匀, 三门湾鱼类分布最均匀。这可能是象山湾内鱼类种类较多, 在各级分类阶元种数也较多且不均匀所致。
分类阶元包含指数对比结果(表 4)显示, 在5个海湾中, 三门湾的科、属、种的分类阶元包含指数值均较高; 杭州湾鱼类目级水平分类阶元包含指数最低, 象山湾科、属分类阶元最低。该结果说明, 三门湾鱼类组成中每一目、科、属所分别拥有的(科、属、种), (属、种)和(种)的平均数目均较多, 亲缘关系近, 分类多样性较低; 而象山湾鱼类在各分类阶元的种类分布不集中, 亲缘关系较远, 分类多样性较高, 此结果与也与平均分类差异指数和分类差异变异指数的分析结果一致。
3.2 G-F多样性传统的生物多样性测度方法只考虑了种水平上的多样性, 无法反映一个生物群落中科级、属级的多样性状况。蒋志刚等(1999)提出的G-F指数是一种将属间的多样性(G指数)和科间的多样性(F指数)进行分析处理后得到的标准化指数, 可以对不同地区科级和属级层面的生物多样性进行比较。目前, G-F指数多用于鸟兽方面的研究(李永民等, 2006; 李忠秋等, 2006; 陈小荣等, 2013; 蒋科毅等, 2013), 近年来有学者尝试将其应用于底栖生物、大型藻类及鱼类多样性方面的分析(胡成业等, 2016; 隋晓云等, 2014; 求锦津等, 2017)。结果表明, 象山湾G-F指数(0.62)最高, 这与象山湾在科级、属级分类阶元数及物种数均较多的结果一致。G-F指数高, 表明象山湾的鱼类在科、属水平上多样性相对较高。象山湾内鱼类物种数最多, 在进化过程中, 生态位相近的物种为了饵料、栖息空间等资源出现激烈的竞争, 可能导致某一物种占有的实际生态位越来越窄, 促使生态位相近的物种向着不同的摄食方式、栖息及习性分化, 以降低种间的竞争关系, 进而使不同物种间达到平衡而共存。G-F指数的大小与物种数的多寡有关。研究发现, 杭州湾物种数最少, 台州湾次之, 但台州湾G-F指数高于物种数较多的三门湾与乐清湾。由分类阶元包含指数分析结果可知, 台州湾鱼类每个目级分类阶元包含的科、属级分类阶元数相对较多, 这可能是导致其G-F指数相对较高的原因。另一方面也说明了台州湾内处于相近生态位的鱼类物种数相对较少, 群落结构比较脆弱, 容易受到干扰破坏。
3.3 相似性在目级水平, 台州湾与乐清湾之间相似性最高, 三门湾与象山湾的相似性最低。在科级水平, 台州湾与乐清湾之间鱼类相似性最高, 象山湾与台州湾的相似性最低。这可能是因为台州湾与乐清湾的海洋生态坏境象山湾而言具有更多的相似性, 导致两海区分布的鱼种比较接近。
综上所述, 象山湾鱼种数最多, 分类多样性也较高, 群落结构比较稳定。乐清湾次之, 其多样性虽不及象山湾, 但仍优于其他三个海湾。台州湾虽鱼种数位列第四, 分类多样性较低, 但G-F指数表明其科、属间的多样性均较高, 这体现了台州湾鱼类群落结构仍然较完整, 但群落结构比较脆弱, 容易受干扰破坏。杭州湾鱼种数位列第五, 三门湾鱼种数位列第二, 但两个海湾的4个指数表明, 多样性均较低, 鱼类群落结构不稳定, 亟待进行生态修复。
4 结论本研究得出, 5个海湾中象山湾鱼类分类多样性较高, 乐清湾次之, 再次为台州湾, 杭州湾与三门湾均较低。这可能与5个海湾自然环境、产业导向、开发程度等有关。开展5个海湾鱼类群落及生态修复与保护工作十分迫切, 且具有重要意义。建议科学合理地优化5个海湾沿海工业、农业、养殖业等产业结构, 尽量减轻人为开发对5个海湾生态环境的负面影响, 实现海洋渔业的可持续发展。
| 王淼, 张丹, 张玉平, 等, 2013. 杭州湾北部张网渔业资源种类组成及群落多样性初步分析. 水产科技情报, 40(4): 188–192, 198 DOI:10.3969/j.issn.1001-1994.2013.04.005 |
| 王淼, 洪波, 张玉平, 等, 2015. 夏、冬季杭州湾北部游泳动物群落结构. 广东海洋大学学报, 35(3): 56–62 DOI:10.3969/j.issn.1673-9159.2015.03.009 |
| 王淼, 洪波, 张玉平, 等, 2016. 春季和夏季杭州湾北部海域鱼类种群结构分析. 水生态学杂志, 37(5): 75–81 |
| 乐清水产局, 1999. 乐清县水产志. 杭州: 浙江人民出版社, |
| 曲方圆, 于子山, 2010. 分类多样性在大型底栖动物生态学方面的应用:以黄海底栖动物为例. 生物多样性, 18(2): 150–155 |
| 齐海明, 徐兆礼, 陈佳杰, 2014. 春秋季台州湾海域鱼类资源的时空分布特征研究. 水产学报, 38(9): 1351–1359 |
| 闫丽娜, 叶深, 李德伟, 等, 2013. 乐清湾口海域春、秋季鱼类种类组成和数量分布. 海洋与湖沼, 44(4): 1062–1067 |
| 李新, 周青松, 俞存根, 等, 2014. 浙江三门湾春季鱼类种类组成及多样性研究. 浙江海洋学院学报(自然科学版), 33(6): 488–494 DOI:10.3969/j.issn.1008-830X.2014.06.002 |
| 李永民, 吴孝兵, 2006. 芜湖市冬夏季鸟类多样性分析. 应用生态学报, 17(2): 269–274 DOI:10.3321/j.issn:1001-9332.2006.02.022 |
| 李圣法, 2005.东海大陆架鱼类群落生态学研究--空间格局及其多样性.上海: 华东师范大学博士学位论文 http://med.wanfangdata.com.cn/Paper/Detail?id=DegreePaper_Y739480 |
| 李忠秋, 蒋志刚, 李春旺, 等, 2006. 陕西老县城自然保护区的鸟类多样性及G-F指数分析. 动物学杂志, 41(1): 32–42 DOI:10.3969/j.issn.0250-3263.2006.01.006 |
| 李娜娜, 董丽娜, 李永振, 等, 2011. 大亚湾海域鱼类分类多样性研究. 水产学报, 35(6): 863–870 |
| 李家芳, 冯应俊, 蔡伟章, 等, 1985. 象山港海洋自然环境分析及其开发利用前景. 海洋环境科学, 4(1): 18–25 |
| 求锦津, 水柏年, 水玉跃, 等, 2017. 浙江6个列岛大型底栖藻类分类多样性. 海洋与湖沼, 48(5): 1004–1013 |
| 沈新强, 袁骐, 2011. 杭州湾增殖放流海域游泳动物群落结构特征分析. 海洋渔业, 33(3): 251–257 DOI:10.3969/j.issn.1004-2490.2011.03.002 |
| 陈小荣, 许大明, 鲍毅新, 等, 2013. G-F指数测度百山祖兽类物种多样性. 生态学杂志, 32(6): 1421–1427 |
| 邵凡户, 1994. 三门湾志. 合肥: 黄山书社, |
| 林龙山, 王燕平, 李渊, 等, 2012. 东山湾及其邻近海域鱼类物种分类多样性. 中国水产科学, 19(6): 1060–1067 |
| 胡成业, 杜肖, 水玉跃, 等, 2016. 浙江6个列岛潮间带大型底栖动物分类多样性. 中国水产科学, 23(2): 458–468 |
| 胡成业, 徐衡, 水柏年, 等, 2015. 唐山湾海域鱼类分类多样性研究. 水生生物学报, 39(3): 608–614 |
| 姜亚洲, 林楠, 袁兴伟, 等, 2014. 象山港游泳动物群落功能群组成与功能群多样性. 海洋与湖沼, 45(1): 108–114 |
| 夏陆军, 周青松, 俞存根, 等, 2016. 乐清湾口海域春秋季鱼类群落多样性研究. 渔业现代化, 43(2): 68–75 DOI:10.3969/j.issn.1007-9580.2016.02.013 |
| 徐宾铎, 任一平, 叶振江, 等, 2007. 青岛近岸水域鱼类群落分类学多样性的研究. 中国海洋大学学报, 37(6): 907–910, 878 |
| 郭小雨, 周青松, 俞存根, 等, 2015. 象山港东北部海域春秋季鱼类生物多样性研究. 浙江海洋学院学报(自然科学版), 34(6): 505–513 DOI:10.3969/j.issn.1008-830X.2015.06.002 |
| 象山县海洋与渔业局, 渔业志编纂办公室, 2008. 象山县渔业志. 北京: 方志出版社, |
| 隋晓云, 陈咏霞, 卫玉龙, 2014. 潭江鱼类群落组成及资源评价. 四川师范大学学报(自然科学版), 37(4): 589–596 DOI:10.3969/j.issn.1001-8395.2014.04.028 |
| 蒋志刚, 纪力强, 1999. 鸟兽物种多样性测度的G-F指数方法. 生物多样性, 7(3): 220–225 DOI:10.3321/j.issn:1005-0094.1999.03.010 |
| 蒋科毅, 吴明, 邵学新, 等, 2013. 杭州湾及钱塘江河口南岸滨海湿地鸟类群落多样性及其对滩涂围垦的响应. 生物多样性, 21(2): 214–223 |
| 谢旭, 俞存根, 周青松, 等, 2013. 杭州湾海域春、秋季鱼类种类组成和数量分布. 海洋与湖沼, 44(3): 656–663 |
| 蔡萌, 徐兆礼, 2009. 浙江三门湾冬夏季鱼类种类组成和数量变化. 上海海洋大学学报, 18(2): 198–205 |
| 冀萌萌, 田阔, 胡成业, 等, 2016. 三门湾春、秋季游泳动物群落组成及物种多样性研究. 水生态学杂志, 37(2): 42–48 |
| Ceschia C, Falace A, Warwick R, 2007. Biodiversity evaluation of the macroalgal flora of the Gulf of Trieste (Northern Adriatic Sea) using taxonomic distinctness indices. Hydrobiologia, 580(1): 43–56 DOI:10.1007/s10750-006-0466-8 |
| Clarke K R, Warwick R M, 1998a. A taxonomic distinctness index and its statistical properties. Journal of Applied Ecology, 35(4): 523–531 DOI:10.1046/j.1365-2664.1998.3540523.x |
| Clarke K R, Warwick R M, 1998b. The taxonomic distinctness measure of biodiversity: weighting of step lengths between hierarchical levels. Marine Ecology Progress Series, 184: 21–29 |
| Rogers S I, Clarke K R, Reynolds J D, 1999. The taxonomic distinctness of coastal bottom-dwelling fish communities of the North-east Atlantic. Journal of Animal Ecology, 68(4): 769–782 DOI:10.1046/j.1365-2656.1999.00327.x |
| Warwick R M, Clarke K R, 1995. New 'biodiversity' measures reveal a decrease in taxonomic distinctness with increasing stress. Marine Ecology Progress Series, 129(6): 301–305 |