 2018, Vol. 49
2018, Vol. 49

长江口水域四种鱼类的耳石微化学研究

http://dx.doi.org/10.11693/hyhz20180300069
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 刘洪波, 姜涛, 邱晨, 杨健. 2018.
- LIU Hong-Bo, JIANG Tao, QIU Chen, YANG Jian. 2018.
- 长江口水域四种鱼类的耳石微化学研究
- OTOLITH MICROCHEMISTRY OF FOUR FISH SPECIES FROM THE CHANGJIANG RIVER ESTUARY, CHINA
- 海洋与湖沼, 49(6): 1358-1364
- Oceanologia et Limnologia Sinica, 49(6): 1358-1364.
- http://dx.doi.org/10.11693/hyhz20180300069
-
文章历史
- 收稿日期:2018-03-28
- 收修改稿日期:2018-07-01
引用本文
刘洪波, 姜涛, 邱晨, 杨健. 2018. 长江口水域四种鱼类的耳石微化学研究[J]. 海洋与湖沼, 49(6): 1358-1364

LIU Hong-Bo, JIANG Tao, QIU Chen, YANG Jian. 2018. OTOLITH MICROCHEMISTRY OF FOUR FISH SPECIES FROM THE CHANGJIANG RIVER ESTUARY, CHINA[J]. Oceanologia et Limnologia Sinica, 49(6): 1358-1364.
长江口水域四种鱼类的耳石微化学研究
1.
中国水产科学研究院淡水渔业研究中心 中国水产科学研究院长江中下游渔业生态环境评价与资源养护重点实验室 无锡 214081;
2. 南京农业大学无锡渔业学院 无锡 214081
2. 南京农业大学无锡渔业学院 无锡 214081
摘要:利用X-射线电子探针微区技术(electron probe microanalysis,EPMA),对长江口不同水域中捕获的焦氏舌鳎、皮氏叫姑鱼、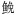 鱼和光泽黄颡鱼耳石中的Sr含量进行耳石微化学研究,结果发现,焦氏舌鳎是典型的海水鱼类,不仅Sr:Ca比(按惯例标准化为Sr:Ca×103)的移动平均值高(>7),而且Sr含量面分析图亦呈现为对应高盐度海水的黄色或红色图谱。虽然皮氏叫姑鱼和鱼从出生直至被捕Sr:Ca比的移动平均值都在3—7间波动,总体上属河口半咸水栖息鱼类,但从Sr含量面分析图来看,皮氏叫姑鱼生活史履历更为复杂,可包括淡水(对应蓝色图谱)和半咸水生境(对应绿色图谱);而鱼仅在长江口较高盐度半咸水或海水生境中活动,未见进入过淡水的履历。光泽黄颡鱼表现出仅利用淡水生境的履历,其Sr:Ca比的移动平均值仅在1.5—3间窄幅波动,整个生活史均在淡水区域里活动,表现为长江口典型的淡水栖息鱼类。本研究从新的角度提供了较为客观、直观和最新的信息,用以较为准确地重建和掌握长江口这些不同鱼类的生境利用特征。
鱼和光泽黄颡鱼耳石中的Sr含量进行耳石微化学研究,结果发现,焦氏舌鳎是典型的海水鱼类,不仅Sr:Ca比(按惯例标准化为Sr:Ca×103)的移动平均值高(>7),而且Sr含量面分析图亦呈现为对应高盐度海水的黄色或红色图谱。虽然皮氏叫姑鱼和鱼从出生直至被捕Sr:Ca比的移动平均值都在3—7间波动,总体上属河口半咸水栖息鱼类,但从Sr含量面分析图来看,皮氏叫姑鱼生活史履历更为复杂,可包括淡水(对应蓝色图谱)和半咸水生境(对应绿色图谱);而鱼仅在长江口较高盐度半咸水或海水生境中活动,未见进入过淡水的履历。光泽黄颡鱼表现出仅利用淡水生境的履历,其Sr:Ca比的移动平均值仅在1.5—3间窄幅波动,整个生活史均在淡水区域里活动,表现为长江口典型的淡水栖息鱼类。本研究从新的角度提供了较为客观、直观和最新的信息,用以较为准确地重建和掌握长江口这些不同鱼类的生境利用特征。
鱼和光泽黄颡鱼耳石中的Sr含量进行耳石微化学研究,结果发现,焦氏舌鳎是典型的海水鱼类,不仅Sr:Ca比(按惯例标准化为Sr:Ca×103)的移动平均值高(>7),而且Sr含量面分析图亦呈现为对应高盐度海水的黄色或红色图谱。虽然皮氏叫姑鱼和鱼从出生直至被捕Sr:Ca比的移动平均值都在3—7间波动,总体上属河口半咸水栖息鱼类,但从Sr含量面分析图来看,皮氏叫姑鱼生活史履历更为复杂,可包括淡水(对应蓝色图谱)和半咸水生境(对应绿色图谱);而鱼仅在长江口较高盐度半咸水或海水生境中活动,未见进入过淡水的履历。光泽黄颡鱼表现出仅利用淡水生境的履历,其Sr:Ca比的移动平均值仅在1.5—3间窄幅波动,整个生活史均在淡水区域里活动,表现为长江口典型的淡水栖息鱼类。本研究从新的角度提供了较为客观、直观和最新的信息,用以较为准确地重建和掌握长江口这些不同鱼类的生境利用特征。
关键词:耳石 微化学 焦氏舌鳎 皮氏叫姑鱼 鱼 光泽黄颡鱼 长江口
鱼 光泽黄颡鱼 长江口
OTOLITH MICROCHEMISTRY OF FOUR FISH SPECIES FROM THE CHANGJIANG RIVER ESTUARY, CHINA
1.
Key Laboratory of Fishery Eco-environment Assessment and Resource Conservation in Middle and Lower Reaches of the Yangtze River, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, China;
2. Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China
2. Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China
Abstract:
An electron probe X-ray micro-analysis (EPMA) was performed to determine Sr and Ca accumulation in otolith of Cynoglossus joyneri, Johnius belengerii, Miichthys miiuy, and Pseudobagrus nitidus collected from different water areas in the Changjiang (Yangtze) River estuary, China. X-ray intensity mapping in the otoliths was analyzed in which the level of Sr content (positively related to salinity) is reflected in different colors. The results show that C. joyneri was a typical brackish and sea water resident fish species, the mean of Sr:Ca ratio (standardized to Sr:Ca×103) of the otolith was high (>7), displaying higher salinity patterns. J. belengerii and M. miiuy were estuarine residents as they had moderate Sr:Ca ratios (3-7), whereas J. belengerii seemed to live in different salinity habitats. Pseudobagrus nitidus lived in freshwater only and the average Sr:Ca ratio was low (1.5-3); this species could be used as a freshwater habitat indicator in the estuary. This study provides an objective and intuitive tool for discriminating the habitat patterns and migration history of the fish life in the estuary.
Key words:
otolith
microchemistry
Cynoglossus joyneri
Johnius belengeri
Miichthys miiuy
Pseudobagrus nitidus
Changjiang River estuary
长江口是太平洋西岸的第一大河口, 其上游可延伸到海洋潮汐所至的安徽大通, 下游经江苏启东、上海南汇等地汇入大海(庄平等, 2006)。受惠于长江干流淡水径流和海洋咸水潮汐的交互作用, 使得这段水域具有多样的生境和丰富的营养物质, 其盐度范围可达0.14—32.31 (史赟荣等, 2012), 适合300余种鱼类依各自的生态习性和盐度需求在此产卵、繁育、索饵、停留调整体液渗透压等等(庄平等, 2006)。这些鱼类或具有极高的经济价值, 或具有独特的科研价值, 或具有良好的水产品开发潜力, 都是生物多样性的宝贵资源。
不同的鱼类有不同的生活履历, 其不同的发育阶段生活环境亦可能截然不同。研究发现, 不同水环境之间的元素成分有很大差异, 比如海水中锶(Sr)的浓度为8mg/L, 比淡水中的浓度要高出100多倍(王夔, 1991)。水环境中元素的变化信息可以在鱼类的硬组织(如耳石)中记录并保存; 因此, 分析耳石中元素的信息可以有效揭示不同鱼类各自的生活史及所经历的环境变化。利用先进的X-射线电子探针微区分析(Electron probe microanalysis, EPMA)、激光烧蚀等离子质谱仪(Laser ablation inductively coupled plasma mass spectrometry, LA-ICPMS)分析和稳定同位素比质谱分析(Isotope ratio mass spectrometry, IRMS)等技术, 对保存于耳石中的环境元素进行深入的分析, 可以反演、重建和掌握鱼类整个生活史过程中大量的生态学、生理学等方面的信息及其生境的时空变化过程。这无疑对鱼类种群资源的管理和生物多样性的保护具有十分重要的意义。
已有研究表明, 积累在长江口鱼类耳石内的Sr含量存在显著的种间差异(窦硕增等, 2011)。利用该元素所记录的履历, 可重建鱼类生活史和进行种群识别(窦硕增等, 2011; Jiang et al, 2014)。作为传统的捕捞调查方法的重要补充, 本研究将选取长江口不同水域中渔获的习见种焦氏舌鳎(Cynoglossus joyneri)、皮氏叫姑鱼(Johnius belengerii)、
本研究上述四种鱼的样本均于2010年夏季采自长江口水域。样品编号、基本生物学数据见表 1。焦氏舌鳎的体长为15.8—18.7cm, 1—2龄; 皮氏叫姑鱼体长为12.2—16.8cm, 1—3龄;
表 1 焦氏舌鳎(JS)、皮氏叫姑鱼(PJ)、
| 种类 | 编号 | 采样时间 | 采样地 | 体长(cm) | 年龄 |
| 焦氏舌鳎 | JS-1 | 2010 | 168海区水域 | 15.8 | 1+ |
| 焦氏舌鳎 | JS-3 | 2010 | 168海区水域 | 18.7 | 1+ |
| 焦氏舌鳎 | JS-4 | 2010 | 168海区水域 | 16.8 | 1+ |
| 焦氏舌鳎 | JS-5 | 2010 | 168海区水域 | 18 | 2 |
| 皮氏叫姑鱼 | PJ-1 | 2010 | 168—169海区水域 | 12.2 | 1 |
| 皮氏叫姑鱼 | PJ-3 | 2010 | 168—169海区水域 | 16.8 | 2 |
| 皮氏叫姑鱼 | PJ-4 | 2010 | 168—169海区水域 | 15.1 | 2 |
| 皮氏叫姑鱼 | PJ-5 | 2010 | 168—169海区水域 | 14.9 | 3 |
| 鱼 |
m-1 | 2010 | 168—169海区水域 | 12.6 | 2 |
| 鱼 |
m-4 | 2010 | 168—169海区水域 | 17.2 | 2 |
| 鱼 |
m-5 | 2010 | 168—169海区水域 | 16 | 2 |
| 鱼 |
m-6 | 2010 | 168—169海区水域 | 12.1 | 2 |
| 光泽黄颡鱼 | GH-2 | 2010 | 167海区水域 | 9.4 | 1 |
| 光泽黄颡鱼 | GH-3 | 2010 | 167海区水域 | 10.9 | 1 |
| 光泽黄颡鱼 | GH-5 | 2010 | 167海区水域 | 8.5 | 2 |
用于EPMA分析的矢耳石样品制备方法参照Jiang等(2017)和Chen等(2017)。即先取出矢耳石, 用去离子水洗净, 干燥并拍照(图 1之A1、B1、C1、D1)后将其平整面朝下包埋于环氧树脂(EpoFix, 丹麦Struers公司)中, 待树脂完全固化后, 切去不含耳石部分多余的树脂, 用备有金刚石磨轮的碾磨机(Discoplan-TS型, 丹麦Struers公司)磨耳石至其核心暴露, 然后配合抛光液在装有机织布抛光盘的磨抛机(LaboPol-35, 丹麦Struers公司)上抛光至耳石表面光滑。接着放于Milli-Q水中用超声波水浴5min, 晾干拍照(图 1之A2、B2、C2、D2)。
 |
|
图 1 长江口水域焦氏舌鳎、皮氏叫姑鱼、 |
为进行EPMA分析, 耳石标本在真空镀膜机(JEE-420, 日本电子株式会社)中镀上碳(carbon)膜(36A, 25s)。然后, 从耳石核心顺着最长轴依次向边缘用EPMA分析仪(JXA-8100型, 日本电子株式会社)测定Sr和Ca的含量。利用SrTiO3和CaCO3作为测定结果准确度检验的标准物质。加速电压和电子束电流分别为15kV和2×10−8 A。根据耳石的不同规格将电子束电流分别聚焦于3μm(光泽黄颡鱼)或5μm(焦氏舌鳎、皮氏叫姑鱼、
为便于研究, Sr:Ca浓度比值(即Sr:Ca比, 按惯例标准化为Sr:Ca×103)根据SrO和CaO的分子量用Sr:Ca×103的方式表达。运用Excel 2013导出数据和绘制线分布图, 并参考Rodionov(2004)、Rodionov等(2005)和卢明杰等(2015)的STARS (Sequential t-test analysis of regime shifts)方法绘制Sr/Ca平均值发生显著变化的趋势转换图, 用移动平均值表示Sr:Ca高低波动特征比值(李孟孟等, 2017), 以客观分析和判定本研究各鱼种生活环境所经历的变化。稳态转换的参数是截断长度为10、置信度水平为0.1、权重为1。Sr:Ca比对应的淡水(< 3)、半咸水(3—7)、海水(> 7)等生境履历的划分方法见Yang等(2011)。其在Sr元素耳石面分布图谱上分别对应为蓝色、绿黄色、红色等区域。
2 结果四种鱼耳石Sr含量面分布分析图见图 1之A3、A4、A5、B3、B4、B5、C3、C4、C5、D3、D4、D5。从耳石核心(0μm)至边缘Sr:Ca比定量线分析图见图 2。由图 1及图 2可清晰地知道四种鱼各自不同的耳石形态和生活史过程中的生境履历。
 |
|
图 2 长江口焦氏舌鳎(JS-1, -3, -4, -5)、皮氏叫姑鱼(PJ-1, -3, -4, -5)、 |
4尾焦氏舌鳎(图 1之JS-1、JS-3、JS-4、JS-5)的耳石表面和边缘光滑, 薄而透明(图 1之A1), 耳石上的年轮在1—2之间(图 1之A2)。耳石的Sr元素含量面分布分析图为统一的黄色和红色(图 1之A3、A4、A5), 从核心到边缘的Sr:Ca比大于7, 最大值达16.3±1.4 (图 2之JS-3)。
4尾皮氏叫姑鱼(图 1之PJ-1、PJ-3、PJ-4、PJ-5)的耳石大而厚, 边缘一侧有脚趾状突起(图 1之B1), 从耳石上推断年龄为2或3(图 1之B2)。Sr含量强度分析图有兰色、绿色甚至红色的环带(图 1之B3、B4、B5), 耳石定量线分析的Sr:Ca比绝大部分在3—7之间波动(图 2之PJ-1、PJ-3、PJ-4、PJ-5)。
4尾
3尾光泽黄颡鱼(图 1之GH-2、GH-3、GH-5)的耳石中间较厚, 边缘呈半透明状(图 1之D1), 从打磨至耳石核心的剖面图上估测鱼龄为1—2之间(图 1之D2)。耳石Sr含量面分析图统一为兰色(图 1之D3、D4、D5), 线定量分析图显示Sr:Ca比较为平稳地在3以下水平波动(图 2之GH-2、GH-3、GH-5)。
3 讨论由于基于耳石中生境元素分析所获得的耳石微化学研究结果具有“有”或“无”的客观特性, 即使仅分析数个样本(Arai et al, 2002; 姜涛等, 2013)仍可准确呈现各种鱼类的栖息地类型和生境变化的过程; 因此成为了解析和反演鱼类生境履历的有力工具(Tsukamoto et al, 1998; Secor et al, 2000; Liu et al, 2015), 为渔业资源管理和人工繁养所需生境重建技术的发展提供了新手段。
本研究结果显示焦氏舌鳎是典型的海水鱼类, 不仅耳石核心Sr:Ca比 > 7, 表现出高Sr的黄色和红色, 其边缘部分也均为高Sr的红色图谱(Sr:Ca比最高达到16.3)(图 2之JS-3), 同时出生后的发育和生长阶段亦都需要生活于较高盐度的海水环境, 无生活于淡水环境的履历, 仅JS-2似短暂出入过半咸水生境。该经济鱼种对咸水环境高盐度的生境有很好的适应能力。
皮氏叫姑鱼和
 |
| 图 3 本研究中基于耳石微化学的长江口水域四种鱼类生境履历模式 Fig. 3 Life history patterns of four fish species from the Changjiang River estuary revealed by otolith microchemical analysis |
生活在相近水域的鱼类, 为避免剧烈的食物竞争, 往往采取分化栖息地(生态位)的策略来缓和生存压力(李忠义等, 2009)。本研究的结果也体现出相关的趋势。皮氏叫姑鱼和
光泽黄颡鱼是生活在长江口内低盐淡水区的优势种(张衡等, 2009), 其Sr:Ca比的移动平均值仅在1.5—3间窄幅波动(图 2), 耳石体现的整个生活史Sr含量均呈现最低的对应典型淡水生境的蓝色(图 1之D3、D4、D5), 即其全生活史过程均生活于长江口淡水生境中, 且该区域一年中环境因素主要是盐度的变化较小, 主要受长江径流控制。适应了此水域环境的光泽黄颡鱼已成为该水域丰度最高的淡水鱼类, 占据了绝对优势(史赟荣等, 2012), 可作为研究长江口淡水环境的典型淡水经济鱼种。
虽然四种鱼的栖息地各不相同, 我们仍可按照Sr:Ca×103的比值< 3为淡水、3—7为半咸水、> 7为海水的标准将四种鱼的生活史型划分为淡水型、河口半咸水型和海水型(图 3)。值得特别注意的是, 本文中对应于不同生态类型的四种鱼, 其孵化和早期生活史所需的水体盐度是相对稳定的, 而不是像刀鲚(Coilia nasus)那样, 早期发育阶段存在淡水孵化和海水孵化两种生态类型(窦硕增等, 2011)。另外, 所研究的4种鱼类虽然各自生活于长江口不同的生境(从纯淡水的光泽黄颡鱼, 半咸水的皮氏叫姑鱼和
对盐度生境的不同耐受范围决定了不同生态类型的鱼类只能较为局限性地生活在河口的不同水域, 但传统的捕捞调查法很难准确了解这些信息。应用耳石微化学研究法所重建的焦氏舌鳎、皮氏叫姑鱼、
参考文献
| 王凯, 章守宇, 汪振华, 等, 2012. 马鞍列岛海域皮氏叫姑鱼渔业生物学初步研究. 水产学报, 36(2): 228–237 |
| 王夔, 1991. 生命科学中的微量元素. 北京: 中国计量出版社, 5 |
卢明杰, 刘洪波, 姜涛, 等, 2015. 大辽河口红狼牙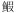 虎鱼耳石微化学的初步研究. 海洋渔业, 37(4): 310–317 DOI:10.3969/j.issn.1004-2490.2015.04.003 虎鱼耳石微化学的初步研究. 海洋渔业, 37(4): 310–317 DOI:10.3969/j.issn.1004-2490.2015.04.003 |
| 史赟荣, 晁敏, 全为民, 等, 2012. 长江口鱼类群落的多样性分析. 中国水产科学, 19(6): 1051–1059 |
| 庄平, 王幼槐, 李圣法, 等, 2006. 长江口鱼类. 上海: 上海科学技术出版社, |
| 李忠义, 戴芳群, 左涛, 等, 2009. 长江口及南黄海水域秋季小黄鱼与皮氏叫姑鱼的食物竞争. 水生态学杂志, 2(2): 67–72 |
| 李孟孟, 姜涛, 陈婷婷, 等, 2017. 长江安庆江段刀鲚耳石微化学及洄游生态学意义. 生态学报, 37(8): 2788–2795 |
| 何小燕, 胡挺, 汪亚平, 等, 2010. 江苏近岸海域水文气象要素的时空分布特征. 海洋科学, 34(9): 44–54 |
| 张衡, 朱国平, 2009. 长江河口潮间带鱼类群落的时空变化. 应用生态学报, 20(10): 2519–2526 |
|
罗海忠, 傅荣兵, 陈波, 等, 2006. 舟山近海鱼胚胎和早期仔鱼的发生及与盐度的关系. 浙江海洋学院学报(自然科学版), 25(1): 15–22 DOI:10.3969/j.issn.1008-830X.2006.01.004 |
|
钟俊生, 楼宝, 袁锦丰, 2005. 鱼仔稚鱼早期发育的研究. 上海水产大学学报, 14(3): 3231–3237 |
| 姜涛, 周昕期, 刘洪波, 等, 2013. 鄱阳湖刀鲚耳石的两种微化学特征. 水产学报, 37(2): 239–244 |
| 窦硕增, 横内一树, 于鑫, 等, 2011. 基于EPMA的耳石Sr:Ca比分析及其在鱼类生活履历反演中的应用实例研究. 海洋与湖沼, 42(4): 512–520 |
|
熊瑛, 刘洪波, 姜涛, 等, 2015. 黄海南部野生银鲳和鱼的耳石元素微化学研究. 海洋学报, 37(2): 36–43 DOI:10.3969/j.issn.0253-4193.2015.02.004 |
| Arai T, Kotake A, Aoyama T, et al, 2002. Identifying sea-run brown trout, Salmo trutta, using Sr:Ca ratios of otolith. Ichthyological Research, 49(4): 380–383 DOI:10.1007/s102280200057 |
| Chen T T, Jiang T, Liu H B, et al, 2017. Do all long supermaxilla-type estuarine tapertail anchovies (Coilia nasus Temminck et Schlegel.1846) migrate anadromously?. Journal of Applied Ichthyology, 33(2): 270–273 DOI:10.1111/jai.13309 |
| Jiang T, Liu H B, Shen X Q, et al, 2014. Life history variations among different populations of Coilia nasus along the Chinese coast inferred from otolith microchemistry. Journal of the Faculty of Agriculture, Kyushu University, 59(2): 383–389 |
| Jiang T, Yang J, Lu M J, et al, 2017. Discovery of a spawning area for anadromous Coilia nasus Temminck et Schlegel.1846 in Poyang Lake, China. Journal of Applied Ichthyology, 33(2): 189–192 DOI:10.1111/jai.13293 |
| Liu H B, Jiang T, Huang H H, et al, 2015. Estuarine dependency in Collichthys lucidus of the Yangtze River Estuary as revealed by the environmental signature of otolith strontium and calcium. Environmental Biology of Fishes, 98(1): 165–172 DOI:10.1007/s10641-014-0246-7 |
| Rodionov S N, 2004. A sequential algorithm for testing climate regime shifts. Geophysical Research Letters, 31(9): L09204 |
| Rodionov S N, Overland J E, 2005. Application of a sequential regime shift detection method to the Bering Sea ecosystem. ICES Journal of Marine Science, 62(3): 328–332 DOI:10.1016/j.icesjms.2005.01.013 |
| Secor D H, Rooker J R, 2000. Is otolith strontium a useful scalar of life cycles in estuarine fishes?. Fisheries Research, 46(1-3): 359–371 DOI:10.1016/S0165-7836(00)00159-4 |
| Tsukamoto K, Nakai I, Tesch W V, 1998. Do all freshwater eels migrate?. Nature, 396(6712): 635–636 DOI:10.1038/25264 |
| Yang J, Jiang T, Liu H B, 2011. Are there habitat salinity markers of the Sr:Ca ratio in the otolith of wild diadromous fishes? A literature survey. Ichthyological Research, 58(2): 291–294 |