 2019, Vol. 50
2019, Vol. 50

南海大眼金枪鱼(Thunnus obesus)摄食生态研究

http://dx.doi.org/10.11693/hyhz20180900215
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 李波, 阳秀芬, 王锦溪, 易木荣, 何雄波, 陶雅晋, 冯波, 颜云榕. 2019.
- LI Bo, YANG Xiu-Fen, WANG Jin-Xi, YI Mu-Rong, HE Xiong-Bo, TAO Ya-Jin, FENG Bo, YAN Yun-Rong. 2019.
- 南海大眼金枪鱼(Thunnus obesus)摄食生态研究
- FEEDING ECOLOGY OF BIGEYE TUNA (THUNNUS OBESUS) IN THE SOUTH CHINA SEA
- 海洋与湖沼, 50(2): 336-346
- Oceanologia et Limnologia Sinica, 50(2): 336-346.
- http://dx.doi.org/10.11693/hyhz20180900215
-
文章历史
- 收稿日期:2018-09-11
- 收修改稿日期:2018-10-20
引用本文
李波, 阳秀芬, 王锦溪, 易木荣, 何雄波, 陶雅晋, 冯波, 颜云榕. 2019. 南海大眼金枪鱼(Thunnus obesus)摄食生态研究[J]. 海洋与湖沼, 50(2): 336-346

LI Bo, YANG Xiu-Fen, WANG Jin-Xi, YI Mu-Rong, HE Xiong-Bo, TAO Ya-Jin, FENG Bo, YAN Yun-Rong. 2019. FEEDING ECOLOGY OF BIGEYE TUNA (THUNNUS OBESUS) IN THE SOUTH CHINA SEA[J]. Oceanologia et Limnologia Sinica, 50(2): 336-346.
南海大眼金枪鱼(Thunnus obesus)摄食生态研究
1.
广东海洋大学水产学院 湛江 524088;
2. 集美大学水产学院 厦门 361021;
3. 广东海洋大学南海渔业资源监测与评估中心 湛江 524088;
4. 广东海洋大学深圳研究院海洋渔业信息化技术中心 深圳 518000
2. 集美大学水产学院 厦门 361021;
3. 广东海洋大学南海渔业资源监测与评估中心 湛江 524088;
4. 广东海洋大学深圳研究院海洋渔业信息化技术中心 深圳 518000
摘要:根据2011年12月—2013年2月于我国南海海域采用金枪鱼延绳钓以及灯光罩网调查捕获的大眼金枪鱼(Thunnus obesus)的各项基础生物学以及胃含物的数据,对其摄食生态进行了分析研究。结果表明:(1)南海海域大眼金枪鱼的食物组成包括鸢乌贼、帆蜥鱼、金色小沙丁鱼、飞鱼、竹荚鱼、鲐鱼、小公鱼属、圆鲹属以及不可辨别的鱼类与虾类,其主要饵料生物为鸢乌贼(%IRI=45.21),其次为金色小沙丁鱼(%IRI=21.36)和帆蜥鱼(%IRI=13.72);(2)空胃率与平均饱满指数随季节变化明显,空胃率在春季时会达到顶峰(37.9%),秋季时最低(16.7%),呈先下降后上升趋势,同时平均饱满指数也在春季达到最高值(0.33),随季节下降并稳定于0.1,空胃率随性腺成熟度的提高有明显上升趋势,平均饱满度指数在性成熟度Ⅰ期与Ⅵ期均呈现高值(1.18、1.04);(3)Shannon-Weiner多样性指数H'和Pielou均匀度指数J随季节变化呈现出明显的变化,Shannon-Weiner多样性指数H'随叉长组基本在1.52—1.72之间变化,Pielou均匀度指数J随性腺成熟度的提高有逐步下降的趋势;(4)营养长度(NR)较高,说明其摄食的饵料成分多、氮来源较复杂,基础食物来源(CR)和生态位总空间(TA)的冬季指标值近乎夏季指标的两倍;通过δ15N稳定同位素法计算大眼金枪鱼的平均营养级为4.73。总体上,南海海域大眼金枪鱼主要以头足类和鱼类为食,其摄食习性与季节和个体发育相关,在食物链中处于较高营养位置。
关键词:南海海域 大眼金枪鱼 摄食习性 碳氮稳定同位素 营养级
FEEDING ECOLOGY OF BIGEYE TUNA (THUNNUS OBESUS) IN THE SOUTH CHINA SEA
1.
Fisheries College, Guangdong Ocean University, Zhanjiang 524088, China;
2. Fisheries College, Jimei University, Xiamen 361021, China;
3. Center of South China Sea Fisheries Resources Monitoring and Assessment, Guangdong Ocean University, Zhanjiang 524088, China;
4. Center of Marine Fisheries Information Technology, Shenzhen Institute of Guangdong Ocean University, Shenzhen 518000, China
2. Fisheries College, Jimei University, Xiamen 361021, China;
3. Center of South China Sea Fisheries Resources Monitoring and Assessment, Guangdong Ocean University, Zhanjiang 524088, China;
4. Center of Marine Fisheries Information Technology, Shenzhen Institute of Guangdong Ocean University, Shenzhen 518000, China
Abstract:
According to the data of individual biology and stomach contents of bigeye tuna (Thunnus obesus) captured in tuna long-line fishing and lighting net surveys in the South China Sea from December 2011 to February 2013; feeding ecology was analyzed and studied. The results show that (1) the food composition of bigeye tuna in the South China Sea area includes Sthenoteuthis oualaniensis, Alepisaurus ferox, Sardinella aurita, Cheilopogon agoo, Trachurus japonicus, Scomber japonicus, Anchoviella sp, Decapterus sp, and indiscriminable fish and shrimps, the main food organism is Sthenoteuthis oualaniensis (%IRI=45.21), Sardinella aurita (%IRI=21.36) and Alepisaurus ferox (%IRI=13.72); (2) the percentage of empty stomach and mean stomach fullness index changed significantly with the season. The percentage of empty stomach reached the highest (37.9%) in spring and the lowest (16.7%) in autumn, which decreased first and then increased, and the mean stomach fullness index reached the highest in spring. Value (0.33), falling with the season and stabilizing at 0.1. The percentage of the empty stomach was associated with gonad maturation. There was a significant increase in the maturity of fish gonad, and the mean stomach fullness index showed high values (1.18, 1.04) in the gonad maturity Ⅰ and Ⅵ stages; (3) the Shannon-Weiner diversity index H' and the Pielou evenness index J showed significant changes with the season (P < 0.01). The Shannon-Weiner diversity index H' ranged between 1.52 and 1.72 with the fork length group, the Pielou evenness index J has gradually decreased with the increase of maturity of the fish gonad. (4) The higher δ15N difference (NR), indicating that it has more bait ingredients and more complicated nitrogen sources; the winter indicator value of the δ13C difference (CR) and Total area (TA) is nearly twice the summer indicator. According to the δ15N stable isotope method, the average trophic level of bigeye tuna was 4.73. In general, bigeye tuna of the South China Sea mainly feed on cephalopods and fish, and their feeding habits are related to season and individual development, and are in a higher trophic level in the food chain.
Key words:
the South China Sea
bigeye tuna Thunnus obesus
feeding habits
stable carbon and nitrogen isotopes
trophic level
我国南海海域是太平洋的五大金枪鱼渔场之一, 同时也是我国大眼金枪鱼渔业灯光罩网与延绳钓作业的主要渔获区域(刘世刚, 2015; 杨胜龙等, 2015)。大眼金枪鱼(Thunnus obesus)隶属于硬骨鱼纲(Osteichthyes)、鲈形目(Perciformes)、鲭科(Scombrida)、金枪鱼属(Thunnus) (伍汉霖等, 1999), 由于其肉质鲜美, 经济价值高, 资源量相对丰富, 一直都是各国远洋渔船主要的捕捞目标之一。在世界金枪鱼的捕捞总产量中, 大眼金枪鱼的捕捞产量在近几年一直稳居第三位(FAO, 2018)。
在海洋生态系统之中, 鱼类的摄食生态研究是促进其结构和功能发展的基础, 同时也是现代鱼类生态学基础理论之一, 通过鱼类的摄食生态的研究可以加深对海洋生态系统中的捕食行为、食物选择方式、食物竞争机制以及捕食者与环境中饵料生物的关系的了解(窦硕增, 1996)。大眼金枪鱼拥有昼伏夜出的运动模式, 并且其视觉系统包含视网膜放光层, 即使光线减弱甚至黑夜也能保持很高的视觉灵敏度, 这使得大眼金枪鱼无论昼夜均可高效摄食, 所以大眼金枪鱼的摄食研究对海洋生态食物网有着重要意义(李军等, 2005)。国外学者对大眼金枪鱼摄食生态中对捕食者与猎物尺寸关系、主要饵料生物的垂直分布以及大眼金枪鱼本身垂直运动习性、垂直分布做了相关研究(Ohshimo et al, 2018; Brill et al, 2005)。国内学者对大眼金枪鱼摄食习性的时空变化以及生态位宽月变化、摄食习性与体长关系以及当前大眼金枪鱼资源的渔业现状等方面作了相关研究(李军等, 2005; 朱国平等, 2007b, 2011; 许柳雄等, 2008); 关于南海金枪鱼的研究, 有冯波等(2014)对南海海域大眼金枪鱼的基础生物学特性做了相关研究; 张衡等(2016)对南沙群岛海域大眼金枪鱼的渔获率和渔获高峰期的研究, 对南海海域大眼金枪鱼摄食生态方面的相关研究则未见报道。
本研究以南海海域的大眼金枪鱼为研究对象, 利用2011年12月份至2013年2月份以延绳钓与灯光罩网作业方式所采集的大眼金枪鱼为样本, 应用胃含物分析法研究大眼金枪鱼的饵料生物组成, 分析大眼金枪鱼摄食习性及其随月份和个体发育的变化, 并且结合碳、氮稳定同位素技术来分析大眼金枪鱼的摄食生态与营养级。本文旨在补充南海海域大眼金枪鱼的食性等基础数据, 为构建南海海洋生物食物链和渔业资源的可持续开发与管理提供参考依据。
1 材料与方法 1.1 样品采集本研究所使用的大眼金枪鱼(Thunnus obesus)样品是2011年12月—2013年2月于南海海域捕获, 捕捞作业方式为延绳钓和灯光罩网, 采样站点分布如图 1。调查渔船为: 2011年12月—2012年8月广西北海的“桂北渔60011”, 2012年2—3月采样调查渔船为海南的“琼儋州15029”, 2012年11月—2013年1月采样调查渔船为广州远洋渔业公司的“穗远渔29”, 2012年12月—2013年2月采样调查渔船为中国水产总公司的“昌荣2”, 除“琼儋州15029”的作业方式为灯光围网外, 其余渔船作业方式均为金枪鱼延绳钓。
 |
| 图 1 采样站点分布图 Fig. 1 Deployment of sampling stations |
在渔船现场按照《海洋调查规范》第六部分海洋生物调查(GB/T 12763.6-2007)(中华人民共和国国家质量监督检验检疫总局等, 2008)进行测定体长、叉长、体重、纯体重以及胃重等生物学数据, 测量长度的精度为1cm, 测量体重精确到0.1g, 测量胃重精确到0.01g, 摄食等级分为0—4级。现场解剖取出大眼金枪鱼的胃和适量干净的背部肌肉分别封装, 保存在–20℃环境条件下, 以作后续的胃含物分析和同位素测定。本研究中采集到大眼金枪鱼样品共387尾, 其中雌性样品共181尾, 叉长范围46—169cm; 雄性206尾, 叉长范围45—230cm。
表 1 南海海域大眼金枪鱼叉长组成 Tab. 1 The statistics of fork-length of T. obesus in the South China Sea
| 性别 | 尾数 | 叉长范围(cm) | 平均值(cm) | 标准误 | 标准差 |
| 雌性 | 181 | 46—169 | 117.92 | 1.88 | 25.35 |
| 雄性 | 206 | 45—230 | 114.04 | 2.07 | 29.76 |
| 雌性+雄性 | 387 | 45—230 | 115.85 | 1.41 | 27.82 |
在实验室, 用传统胃含物分析法对大眼金枪鱼的食物组成进行研究, 使用双筒解剖镜(Leica Zoom 2000 Z45V)并参考海洋生物分类资料对胃含物内的饵料生物进行分类鉴定(中国科学院动物研究所, 1962; 成庆泰等, 1987), 精确到最低分类阶元, 使用精度为0.01g的电子天平(Shimadzu Auy220)对饵料生物分别进行称重。
用于同位素测定的肌肉样本41尾, 叉长范围68—157cm, 采自2012年11月—2013年1月。将取得的大眼金枪鱼肌肉剪碎后, 在冷冻干燥机(ALPHA 1-4LDplus) –50℃恒温条件下干燥48h, 最后用玛瑙研钵充分磨至粉末状, 进行稳定同位素测定。实验样品碳、氮稳定同位素是在中国科学院水生生物研究所东湖生态试验站同位素分析实验室采用美国Thermo公司的Carlo Erba EA-1110元素分析仪与Delta Plus Finnigan稳定同位素质谱仪测定所得, 样品的碳、氮含量用‰表示。为保证实验结果的准确性和仪器的稳定性, 先进行仪器校正后, 再对同一样品的碳、氮稳定同位素分别进行测定。
1.4 数据处理与统计本研究采用的个数百分比(N%)、饵料质量百分比(W%)、出现频率(F%)和相对重要性指数百分比(IRI)等都是用于评价饵料重要性的重要指标, 在目前的鱼类食性类型研究中, 鱼类的食性类型通常采用饵料生物的生态类群出现频次百分比来划分, 但是并没有一个明确标准的划分标准。张波等(2003)在东黄海六种鳗的食性研究中, 采用了多数原则定位主要摄食对象, 在本研究中也采用同样方法分析各种饵料的重要性水平(Hyslop, 1980), 其计算公式如下:

鱼类的食物重量与鱼体体长之间有较好的相关性, 因此可以利用饱满指数这个指标反映鱼类的摄食情况(薛莹等, 2003; 王学昉等, 2013), 但是单以饱满指数这一指标来反应大眼金枪鱼的摄食强度变化并不完全准确, 所以在本实验中采用了饱满指数和空胃率两个指标来共同描述大眼金枪鱼的摄食强度变化状况, 使其描述更加准确, 其计算公式如下:

鱼类摄食生态位宽度, 即鱼类食物组成的多样性用Shannon-Wiener多样性指数H'和Pielou均匀度指数J(孙军等, 2004)表示, 计算公式如下:
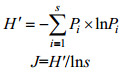
式中, s为饵料生物种数, Pi为饵料生物i在食物中所占的个数百分比。Shannon-Wiener多样性指数H'和Pielou均匀度指数J对于优势饵料和其他饵料的丰度相差悬殊的食物组成能较好的反应, 同时Shannon-Wiener指数对于稀有饵料很敏感, 因此, 适合描述鱼类总的食物生态位宽度(Marshall et al, 1997), 鱼类的Shannon-Wiener多样性指数H'和Pielou均匀度指数J的变化趋势一致, 两指数可以更加直观的反应鱼类总的食物的摄食生态位宽度。
鱼类营养生态位通过R语言分析稳定同位素参数计算并用营养结构框架图表示, 营养级(Trophic level, TL)按以下公式所得:

公式中的δX为δ13C或δ15N, Rsample为所测样品的同位素比值, 碳同位素是12C/13C, 氮同位素是14N/15N; Rstandard为标准物质的同位素比值, 碳氮稳定同位素测定的标准物质为国际通用标准物质箭石(Peedee belemnile limestone)。在水生生态系统中, 碳稳定同位素比值δ13C、可用于指示食物来源, 而氮稳定同位素比值δ15N用于确定研究对象的营养级(Mariotti et al, 1983; Zanden et al, 2001; 李云凯, 2014)。在进行稳定同位素检测时, 需要通过在测试样品中加入标准样品来保证研究结果的准确性和仪器的稳定性, 碳、氮稳定同位素比值精密度为±0.02×10–3; δ15Nsample为样品所测得氮稳定同位素值; δ15N0为基线生物氮稳定同位素值, δ15Nc为一个营养等级的氮同位素富集度, 取值为3.4‰ (Post, 2002); TLbasic为基线生物营养级, 在生态系统中, 选取捕食者主要摄食的饵料生物中, 常年存在并且食性较为简单的一种饵料生物, 作为计算营养级的基线生物(彭士明等, 2011)。本研究基线生物选择南海鸢乌贼(Sthenoteuthis oualaniensis), 该物种属于南海海域大眼金枪鱼的主要摄食对象, 并且其在所研究的南海海域中分布十分广泛、食性较简单, 完全符合作为计算大眼金枪鱼营养级基线生物的条件, 营养级为TLbasic=3.3 (张宇美, 2014)。
本研究使用Excel 2013和SPSS 22统计软件对大眼金枪鱼的饵料组成、摄食习性和营养级随生长发育的变化和摄食强度的变化等研究数据进行统计分析。
2 结果 2.1 大眼金枪鱼的食物组成胃含物分析表明, 南海海域大眼金枪鱼摄食种类包括鱼类、头足类、甲壳类等, 其中可辨别鱼类有8种(表 2)。该海域大眼金枪鱼主要摄食鱼类(%IRI=48.17)、头足类(%IRI=45.63), 优势饵料生物为鸢乌贼(%IRI=45.21), 其次为金色小沙丁鱼(Sardinella aurita) (%IRI=21.36)和帆蜥鱼(Alepisaurus ferox) (%IRI=13.72)等。
表 2 南海海域大眼金枪鱼的食物组成 Tab. 2 The diet composition of T. obesus in the South China Sea
| 饵料种类 | 饵料质量 百分比(%) |
个数 百分比(%) |
出现频次 (%) |
相对重要性指数 (%IRI) |
出现频次 百分比组成 |
| 未辨认头足类 Unidentified Cephalopoda |
1.52 | 1.94 | 6.56 | 0.42 | 4.47 |
| 鸢乌贼 Sthenoteuthis oualaniensis |
48.53 | 27.59 | 31.97 | 45.21 | 21.79 |
| 未辨认虾类 Unidentified shrimps |
0.36 | 15.30 | 21.31 | 6.20 | 14.53 |
| 未辨认鱼类 Unidentified fishes |
10.99 | 16.16 | 25.41 | 12.82 | 17.32 |
| 帆蜥鱼 Alepisaurus ferox |
12.41 | 15.73 | 26.23 | 13.72 | 17.88 |
| 金色小沙丁鱼 Sardinella aurita |
21.38 | 21.12 | 27.05 | 21.36 | 18.44 |
| 小公鱼属 Thryssa |
0.72 | 0.43 | 1.64 | 0.03 | 1.12 |
| 飞鱼 Cheilopogon agoo |
0.11 | 0.22 | 0.82 | — | 0.56 |
| 竹荚鱼 Trachurus japonicus |
0.25 | 0.22 | 0.82 | — | 0.56 |
| 鲐鱼 Scanber japonicus |
1.88 | 1.08 | 4.10 | 0.22 | 2.79 |
| 圆鲹属 Decapterus |
0.12 | 0.22 | 0.82 | — | 0.56 |
| 注:“—”表示所占比例 < 0.01%或无 | |||||
大眼金枪鱼的摄食习性具有明显的月变化(图 2), 根据饵料种类重量百分比可以看出, 鱼类与头足类为大眼金枪鱼的主要饵料种类, 2012年8、11、12月, 不可辨别鱼类为大眼金枪鱼的主要摄食对象。2012年3月份大眼金枪鱼食物中鸢乌贼所占比例最大, 达到了61.96%, 平均饵料种类重量百分比为12.82%, 除不可辨别鱼类外, 鸢乌贼所占比例最大, 其次是金色小沙丁鱼、帆蜥鱼。
 |
| 图 2 南海海域大眼金枪鱼主要饵料类群重量百分比的月变化 Fig. 2 Monthly variation in weight percentage of major prey groups to the diets of T. obesus in the South China Sea |
大眼金枪鱼的空胃率和平均饱满指数都有明显的季节变化(图 3), 空胃率在春季达到了最高值(37.9%), 秋季降为最低(16.7%), 随季节变化总体呈先下降后上升的趋势; 平均饱满指数在春季达到了峰值(0.326), 在夏季达到最低值(0.101), 随季节变化总体呈先下降后平稳的趋势。
 |
| 图 3 南海海域大眼金枪鱼各季节的空胃率和平均饱满指数(误差线表示标准误差) Fig. 3 Seasonal variation in percentage of empty stomach and mean stomach fullness index of T. obesus in the South China Sea (standard error) |
单因素方差分析表明, 各叉长组之间空胃率和平均饱满指数均表现为差异性极显著(P= < 0.01)。其中, 空胃率的变化规律不明显, 平均饱满指数总体呈现出先下降后上升再回落的趋势(图 4)。
 |
| 图 4 南海海域大眼金枪鱼各叉长组的空胃率和平均饱满指数 Fig. 4 The percentage of empty stomach and mean stomach fullness index for each fork length class of T. obesus in the South China Sea |
随着性腺成熟度的提升, 大眼金枪鱼的空胃率随之提升, Ⅵ期时空胃率最大达到100%, Ⅰ期时空胃率最小为16%, 各性腺成熟度之间空胃率变化极显著(P= < 0.01)。平均饱满指数随性腺成熟度变化极显著(P= < 0.01), 呈现出先下降后上升的趋势, 平均饱满指数在Ⅰ期时最大为1.18, 在Ⅲ期时, 平均饱满指数最小为0.09。
2.4 摄食生态位宽度随季节、叉长组及性腺成熟度变化根据南海海域大眼金枪鱼Shannon-Weiner多样性指数H′和Pielou均匀度指数J与季节变化差异(图 6), 可以看出在夏季时H′最低为0.815, 秋季时最高为1.609;对于Pielou均匀度指数J, 夏季时达到最低值为0.603, 秋季时达到最大值1.000; H′和J总体上呈现为先下降后上升的趋势, 两者数值变化呈正相关关系。
 |
| 图 6 南海海域大眼金枪鱼各季节Shannon-Weiner多样性指数H'和Pielou均匀度指数J Fig. 6 The Shannon-Weiner diversity index H′ and the Pielou evenness index J for each season of T. obesus in the South China Sea 注: H′: Shannon-Weiner多样性指数H′; J: Pielou均匀度指数J |
Shannon-Weiner多样性指数H′和Pielou均匀度指数J随体长组变化时, Pielou均匀度指数J最低为0.62(100—110cm), Shannon-Weiner多样性指数H’除100—110cm叉长组外, 其余各叉长组基本在1.52—1.72之间变化。
其Shannon-Weiner多样性指数H′和Pielou均匀度指数J随性腺成熟度变化时, Pielou均匀度指数J随着性腺成熟度的提高而缓慢降低, Ⅰ期时最高为0.98, 在Ⅵ期时带到最低点, 其值为0.56; Shannon-Weiner多样性指数H′在Ⅲ期时最高为1.74, 在Ⅵ期时最低为0.60。
2.5 碳、氮同位素特性、营养生态位与营养等级南海海域大眼金枪鱼δ13C值范围为–23.130‰–18.502‰, 平均值为–20.607‰, 最大差值为4.628‰; δ15N值范围为6.706‰—19.455‰, 平均值为15.290‰, 最大差值为12.749‰。
通过构建营养结构框架图分析大目金枪鱼稳定同位素特征值(图 9, 表 3)。由图可得秋冬季大眼金枪鱼生态位总空间(TA, total area)和核心生态位空间(SEAc, standard ellipse area)的重叠率较高, δ15N和δ13C的宽幅在冬季比秋季更广泛; 在大眼金枪鱼秋冬季的同位素生态位指标中, 营养长度(NR, δ15N difference)、营养多样性(CD, mean distance to centroid)、核心生态位空间(SEAc)、鱼类群落的整体密度(MNND, mean nearest neighbor distance)和鱼类群落营养生态位分布范围(SDNND, standard deviation of nearest neighbor distance)指标值均为秋季高于冬季, 而基础食物来源(CR)和生态位总空间(TA)的冬季指标值近乎夏季指标的两倍。
 |
| 图 9 基于碳氮稳定同位素南海海域大眼金枪鱼营养框架图 Fig. 9 The bio-plots based on stable C and N isotopes of T. obesus in the South China Sea |
表 3 南海海域大眼金枪鱼的营养生态位变量 Tab. 3 Trophic niche variables for T. obesus in the South China Sea
| 生态位指标 | 秋季 | 冬季 | 平均 |
| CR | 2.27 | 4.63 | 3.45 |
| NR | 11.30 | 10.90 | 11.10 |
| CD | 2.69 | 2.26 | 2.48 |
| TA | 14.20 | 28.00 | 21.10 |
| SEAc | 7.85 | 7.71 | 7.78 |
| MNND | 1.04 | 0.75 | 0.90 |
| SDNND | 1.19 | 0.80 | 1.00 |
| 注: CR: δ13C差值; NR: δ15N差值; CD:平均离心距离; TA:凸多边形总面积; SEAc:标准椭圆面积; MNND:平均最邻近距离; SDNND:平均最邻近距离标准差 | |||
根据δ15N值计算出不同叉长组大眼金枪鱼的营养级范围是4.28—5.31, 平均营养级为4.73(表 4), 各叉长组之间营养级TL差异性不显著(P > 0.05)。
表 4 南海海域大眼金枪鱼δ15N、δ13C和营养级随生长的变化 Tab. 4 δ15N, δ13C and trophic level variation of different T. obesus wolf-herring fork length groups in the South China Sea
| 叉长组(mm) | 频数 | 平均δ13C (‰) | 平均δ15N (‰) | 营养级 |
| < 110 | 2 | –20.765 | 14.759 | 4.73 |
| 110-120 | 15 | –20.687 | 16.725 | 5.31 |
| 120-130 | 2 | –20.434 | 13.215 | 4.28 |
| 130-140 | 11 | –20.877 | 14.950 | 4.79 |
| 140-150 | 7 | –20.227 | 13.377 | 4.33 |
| 150-160 | 4 | –20.243 | 15.417 | 4.93 |
从表 2可知, 南海海域大眼金枪鱼的摄食鱼类主要为金色小沙丁鱼、帆蜥鱼, 头足类主要为鸢乌贼, 从大眼金枪鱼饵料种类所栖息的水层可知, 大眼金枪鱼主要摄食中上层游泳动物,同时兼食个别深水鱼类(Post, 2012; Potier et al, 2007),例如饵料食物中帆蜥鱼为栖息在0—1000m水层(FishBase)。Borodulina (1981)认为, 大眼金枪鱼主要摄食上层及中上层的鱼类和头足类, 许柳雄等(2008)对印度洋中西部海域大眼金枪鱼的研究得出, 大眼金枪鱼的最适合摄食水层约为100—300m, 即表层与中层之间水层。朱国平等(2011)对太平洋中东部大眼金枪鱼的分析得出, 200—350m水层中大眼金枪鱼的摄食活动非常活跃, 本研究中的大眼金枪鱼捕食的饵料生物所栖息水层与其研究结果相一致, 因此在本研究中判断南海海域大眼金枪鱼摄食习性主要为捕食中上层鱼类及头足类。
根据各个海域大眼金枪鱼饵料组成对比可以发现(表 5), 大眼金枪鱼是广食性鱼类, 食物来源比较广泛, 结合胃含物分析的结果, 其食物来源主要为鱼类、头足类和甲壳类。许柳雄等(2008)对印度洋中西部海域大眼金枪鱼的食物组成分析得出大眼金枪鱼的饵料重要性依次为头足类(%IRI=51.76)、鱼类(%IRI=29.52)和甲壳类(%IRI=12.07)。朱国平等(2007a)对大西洋西部大眼金枪鱼的食物饵料分析得出大眼金枪鱼的主要食物为鱼类(%IRI=84.9)、头足类(%IRI=11.6)和甲壳类(%IRI=1.4)。本研究所得结论为头足类(%IRI=48.17)、鱼类(%IRI=45.63)和虾类(%IRI=6.2), 造成各类食物重要性指数些许差异的原因可能是各海域之间物种资源量差异。
表 5 各海域大眼金枪鱼食物组成对比 Tab. 5 Comparison of food composition of T. obesus in different sea areas
| 海域 | 样品尾数/实胃数 | 叉长范围(cm) | 主要摄食种类 |
| 大西洋西部海域 (朱国平等, 2007a) |
780/607 | 85—206 | 沙丁鱼Sardina pilchardus, 鱿鱼Loligo pealei, 虾类Shrimp, 乌鲂Brama brama, 帆蜥鱼Alepisaurus ferox, 鲯鳅Coryphaena hippurus, 鲐鱼Scomber japonicus, 秋刀鱼Cololabis saira, 乌贼Sepia officinalis, 飞鱼Exocoetus volitans, 海蜘蛛Pycnogonida, 七星鱼Scomberomorus sinensis及杂鱼Other fishes |
| 大平洋中东部海域 (郑晓春等, 2015) |
1535/1535 | — | 帆蜥鱼Alepisaurus ferox, 竹荚鱼Trachurus japonicus, 秋刀鱼Cololabis saira, 蛇鲭Gemplus serpens, 宽尾鳞鲀Abalistes stellatu, 虱目鱼Chanos chanos, 白腹鲭Scomber japonicus, 低褶胸鱼Sternoptyx diaphana, 柔鱼科Ommastrephidae, 乌贼科Sepiidae, 刚毛对虾Peneus setiferus, 海蜘蛛Pycnogonida |
| 印度洋中南部海域 (李攀等, 2010) |
435/163 | 57—182 | 柔鱼科鱼类Ommastrephida, 帆蜥鱼Alepisaurus ferox, 虾类Shrimp, 刺鲅Acanthocybium solandr, 鲐鱼Scomber japonicus, 秋刀鱼Cololabis saira, 乌鲂Brama brama, 蜞鳅Coryphaena hippurus, 沙丁鱼Sardina pilchardus, 银鲳Ampus argenteus和带鱼Trichiutus lepturus |
| 印度洋中西部海域 (许柳雄等, 2008) |
308/154 | 95.4—179.2 | 鱿Loligo, 鲐Scomber, 沙丁鱼Sardina, 乌贼Brama, 蟹Portunus trituberculatus, 竹筴鱼Trachurus, 蛇鲭Gempylus, 乌鲂Brama, 刺鲷Sparus, 对虾Penaeus, 帆蜥Alepisaurus, 鳞鲀Balistes, 水母Physalia和杂鱼Other fishes |
随着季节的更替, 海洋中的温度、盐度、含氧量、潮汐、潮流等环境因子会发生一定的变化, 鱼类摄食强度就会随着季节变化, 以季节性变化反映了鱼体代谢强度、摄食行为以及繁殖行为的变化(Battaglia et al, 2013; 李军, 1994; 陶雅晋等, 2017)。在本研究中, 对南海海域大眼金枪鱼各季节摄食强度进行分析, 得出了春季时摄食强度最大, 随季节推移先下降后回升的结论, 这与朱国平等(2007a)对大眼金枪鱼的季节性摄食月变化作初步研究所得结论基本一致, 但在秋季时与其开始回升的结果不同的是, 本研究中秋季时大眼金枪鱼摄食强度仍处于最低值, 猜测其原因, 本研究海域不同于地处赤道的印度洋中西部海域, 南海海域金枪鱼类由于高度洄游的特性, 鱼群每年8—10月间洄游进入南海(陈炎等, 2000; 张鹏, 2010; 王中铎等, 2012), 因此秋季时人类捕捞作业也正值高峰期, 使该海域金枪鱼食物有所减少, 所以会造成研究中秋季摄食强度低的结果。
鱼类在自身的生长发育过程中, 其个体体长体重增加, 运动能力也随之加强, 生殖系统也随之发育成熟, 并伴随着性成熟度的提高, 需要消耗的能量随之提高, 通过加大捕食饵料量来加大能量的摄入, 其摄食能力与消化饵料能力也会发生一定的变化(崔奕波等, 2001)。通过研究显示, 由于成鱼需要维持的生命活动以及繁殖行为, 能量消耗量变大, 更容易出现空胃率高的现象。根据图 4、图 5可知, 大眼金枪鱼在个体叉长较小时, 由于生长需要较大营养供给, 空胃率会偏高, 随着叉长的增长, 空胃率呈现出逐步上升的趋势, 同时随着性成熟度的提高, 空胃率也随之提升; 对于研究中出现春季样品空胃率偏高的问题, 分析其原因, 该季节采样的捕捞方式为延绳钓, 由于大眼金枪鱼从海中被捕捞到船上, 压强变化较大, 会导致一部分鱼发生吐胃的现象, 其胃含物均无法采集到, 所以造成空胃率偏高(郑晓春等, 2015), 不能准确做出鉴定, 影响到大眼金枪鱼摄食分析的准确性, 故建议对钓获时已经死亡的大眼金枪鱼做胃含分析。
 |
| 图 5 南海海域大眼金枪鱼各性腺成熟度的空胃率和平均饱满指数 Fig. 5 The percentage of empty stomach and mean stomach fullness index for each maturity of fish gonad of T. obesus in the South China Sea |
生态位宽度是一种用以描述在特定生态环境中某一物种进行所有的生态学过程时, 所具有的功能地位, 其生态学过程包括生物入侵、种群定居、发育繁衍以及消退、消亡等过程(张光明等, 1997)。牛克昌等(2009)在群落构建的中性理论和生态位理论研究中表明, 生态位理论在对决定群落构建和生物多样性维持中起到了重要的作用。从本研究的图 6—图 8中可以看出, Shannon-Weiner多样性指数H’在随季节变化时出现较大变动, 其原因可能是由于大眼金枪鱼具有很强的洄游性, 会随季节变化在外洋与南海海域随海流进出, 造成其捕食对象发生变化, 进而引起生态位宽的变化, 这与李军等(2005)对大眼金枪鱼的洄游性综述相对应。朱国平等(2007b)对大西洋西部大眼金枪鱼摄食生态的初步研究结果表示在叉长为90—160cm时, 其生态位宽度H’稳定在1.5—1.7范围内, 叉长大于160cm时, 随着个体生长, 大眼金枪鱼的生态位宽呈现下降的趋势。本研究中南海海域大眼金枪鱼在叉长组在110—150cm时生态位宽H’同样稳定在1.5—1.7之间, 但未对高于150cm的叉长组进行均匀划分, 是由于在本研究中由于大于150cm叉长的金枪鱼较少, 故为避免误差并未对高于150cm的叉长组进行均匀划分。
 |
| 图 7 南海海域大眼金枪鱼各叉长组Shannon-Weiner多样性指数H′和Pielou均匀度指数J Fig. 7 The Shannon-Weiner diversity index H′ and the Pielou evenness index J for each fork length group of T. obesus in the South China Sea 注: H′: Shannon-Weiner多样性指数H′; J: Pielou均匀度指数J |
 |
| 图 8 南海海域大眼金枪鱼各性腺成熟期Shannon-Weiner多样性指数H′和Pielou均匀度指数J Fig. 8 The Shannon-Weiner diversity index H′ and the Pielou evenness index J for each maturity of fish gonad of T. obesus in the South China Sea 注: H′: Shannon-Weiner多样性指数H′; J: Pielou均匀度指数J |
通过胃含物分析计算H'和J仅表示大眼金枪鱼被捕获近期的摄食生态位宽, 采用碳氮稳定同位素计算大眼金枪鱼营养结构可以表示其整个生态学过程的营养生态位状况(颜云榕等, 2011; 张欢等, 2013)。从营养框架图(图 9)可知, 南海海域大眼金枪鱼在整个生态学过程中基础食物来源(CR)与营养长度(NR)来源广泛, δ15N差值高达12.749‰, 说明其摄食的饵料成分多, 氮来源较复杂, 可印证本研究中大眼金枪鱼食物来源广泛, 是广食性鱼类这一结论; 其核心生态位空间(SEAc)在秋冬季重叠率较高, 说明其不同季节主要摄食对象基本一致, 未发生变化。
营养级是一种用以反映在食物网中, 捕食者所处位置的生态系统营养动力学的概念, 在海洋鱼类的食物网中营养级同样适用, 目前的研究中海洋生态系统中海洋生物营养级的研究越来越重要(纪炜炜等, 2010)。在本研究中, 通过应用碳、氮同位素分析法来研究南海海域大眼金枪鱼的营养级, 能够弥补传统胃含物分析法计算营养级的不足, 可更加真实地反映南海海域大眼金枪鱼的摄食生态状况。由表 4显示, δ15N平均值为15.290‰, 除叉长组110—120mm的营养级偏高外, 其余叉长组均稳定在4.20—4.90之间, 各叉长组营养级以δ15N为指标所计算的南海海域大眼金枪鱼平均营养级为4.73, 高于肖悦悦等(2015)计算的热带太平洋中部海域大眼金枪鱼营养级3.42, 造成原因可能是本研究不同于其使用食物网中各营养级相对度量的划分使用摄食营养级计算(Hasler, 1975); 在FishBase中大眼金枪鱼营养级为4.50, 这与本研究所得营养级相近, 造成差异的原因一方面是由于基线生物选取的不同, 另一方面是由于采样海域不同, 其大眼金枪鱼的摄食习性会有所不同。
4 结论南海海域大眼金枪鱼主要以头足类和鱼类为食, 其摄食习性随季节变化和个体生长发育变动明显, 其营养级较高, 在食物链中处于较高级营养位置。本研究补充了南海大眼金枪鱼的摄食习性和营养结构资料, 在构建南海海洋生物食物网、渔业资源的可持续开发和管理方面有重要参考意义。
参考文献
王中铎, 郭昱嵩, 颜云榕, 等. 2012. 南海大眼金枪鱼和黄鳍金枪鱼的群体遗传结构. 水产学报, 36(2): 191-201 |
王学昉, 许柳雄, 周成, 等. 2013. 金枪鱼围网鲣的捕获率与其摄食状态的关系. 大连海洋大学学报, 28(6): 622-626 DOI:10.3969/j.issn.2095-1388.2013.06.020 |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.6-2007海洋调查规范第6部分: 海洋生物调查.北京: 中国标准出版社, 1-157
|
中国科学院动物研究所.. 1962. 南海鱼类志. 北京: 科学出版社
|
牛克昌, 刘怿宁, 沈泽昊, 等. 2009. 群落构建的中性理论和生态位理论. 生物多样性, 17(6): 579-593 |
冯波, 李忠炉, 侯刚. 2014. 南海大眼金枪鱼和黄鳍金枪鱼生物学特性及其分布. 海洋与湖沼, 45(4): 886-894 |
成庆泰, 郑葆珊. 1987. 中国鱼类系统检索. 北京: 科学出版社
|
朱国平, 刘维, 戴小杰, 等. 2011. 热带太平洋中东部大眼金枪鱼摄食强度的时空变化. 大连海洋大学学报, 26(1): 68-73 DOI:10.3969/j.issn.1000-9957.2011.01.013 |
朱国平, 许柳雄, 周应祺, 等. 2007a. 印度洋中西部和大西洋西部水域大眼金枪鱼的食性比较. 生态学报, 27(1): 135-141 |
朱国平, 周应祺, 许柳雄, 等. 2007b. 大西洋西部大眼金枪鱼摄食生态的初步研究. 水产学报, 31(1): 23-30 |
伍汉霖, 邵广昭, 赖春福. 1999. 拉汉世界鱼类名典. 基隆: 水产出版社
|
刘世刚, 2015.基于灯光罩网法的南海黄鳍金枪鱼声学评估技术研究.大连: 大连海洋大学硕士学位论文, 1-3
|
许柳雄, 朱国平, 宋利明. 2008. 印度洋中西部水域大眼金枪鱼的食性. 水产学报, 32(3): 387-394 |
孙军, 刘东艳. 2004. 多样性指数在海洋浮游植物研究中的应用. 海洋学报, 26(1): 62-75 DOI:10.3321/j.issn:0253-4193.2004.01.007 |
纪炜炜, 李圣法, 陈雪忠. 2010. 鱼类营养级在海洋生态系统研究中的应用. 中国水产科学, 17(4): 878-887 |
李军. 1994. 渤海鲈鱼食物组成与摄食习性的研究. 海洋科学, (3): 39-44 |
李军, 李志凌, 叶振江. 2005. 大眼金枪鱼渔业现状和生物学研究进展. 齐鲁渔业, 22(12): 35-38 |
李攀, 陈锦淘, 朱国平, 等. 2010. 印度洋中南部大眼金枪鱼生物学特性研究. 海洋渔业, 32(3): 283-289 DOI:10.3969/j.issn.1004-2490.2010.03.008 |
李云凯. 2014. 稳定同位素技术在鲨鱼摄食和洄游行为研究中的应用. 应用生态学报, 25(9): 2756-2764 |
杨胜龙, 靳少非, 吴祖立, 等. 2015. 太平洋金枪鱼渔场关键次表层环境变量的季节变化. 海洋科学, 39(5): 36-46 |
肖悦悦, 朱江峰, 戴小杰. 2015. 热带太平洋中上层鱼类群落结构研究. 海洋湖沼通报, (4): 96-104 |
张欢, 谢平, 吴功果, 等. 2013. 日本沼虾与秀丽白虾的营养生态位. 环境科学研究, 26(1): 22-26 |
张波, 唐启升. 2003. 东、黄海六种鳗的食性. 水产学报, 27(4): 307-314 |
张鹏, 杨吝, 张旭丰, 等. 2010. 南海金枪鱼和鸢乌贼资源开发现状及前景. 南方水产科学, 6(1): 68-74 DOI:10.3969/j.issn.1673-2227.2010.01.012 |
张衡, 吴祖立, 周为峰, 等. 2016. 南海南沙群岛灯光罩网渔场金枪鱼科渔获种类、渔获率及其峰值期. 海洋渔业, 38(2): 140-148 DOI:10.3969/j.issn.1004-2490.2016.02.004 |
张光明, 谢寿昌. 1997. 生态位概念演变与展望. 生态学杂志, 16(6): 46-51 DOI:10.3321/j.issn:1000-4890.1997.06.010 |
张宇美, 2014.基于碳氮稳定同位素的南海鸢乌贼摄食生态与营养级研究.湛江: 广东海洋大学硕士学位论文, 38-39
|
陈炎, 陈丕茂. 2000. 南沙群岛金枪鱼资源初探. 远洋渔业, (2): 7-10 |
郑晓春, 戴小杰, 朱江峰, 等. 2015. 太平洋中东部海域大眼金枪鱼胃含物分析. 南方水产科学, 11(1): 75-80 DOI:10.3969/j.issn.2095-0780.2015.01.011 |
陶雅晋, 莫檬, 何雄波, 等. 2017. 南海黄鳍金枪鱼(Thunnus albacores)摄食习性及其随生长发育的变化. 渔业科学进展, 38(4): 1-10 |
崔奕波, 解绶启, 朱晓鸣, 等. 2001. 鱼类生长与生物能量学研究新进展. 浙江海洋学院学报(自然科学版), 20(S1): 11-15 |
彭士明, 施兆鸿, 尹飞. 2011. 利用碳氮稳定同位素技术分析东海银鲳食性. 生态学杂志, 30(7): 1565-1569 |
窦硕增. 1996. 鱼类摄食生态研究的理论及方法. 海洋与湖沼, 27(5): 556-561 DOI:10.3321/j.issn:0029-814X.1996.05.016 |
颜云榕, 卢伙胜, 金显仕. 2011. 海洋鱼类摄食生态与食物网研究进展. 水产学报, 35(1): 145-153 |
薛莹, 金显仕. 2003. 鱼类食性和食物网研究评述. 海洋水产研究, 24(2): 76-87 |
Battaglia P, France A, Pierpaolo C et al, 2013. Feeding habits of the Atlantic bluefin tuna, Thunnus thynnus (L. 1758), in the central Mediterranean Sea (Strait of Messina). Helgoland Marine Research, 67(1): 97-107 DOI:10.1007/s10152-012-0307-2 |
Borodulina O D, 1981. Food composition of the yellowfin tuna Thunnus albacares (Bonnaterre) (Scombridae) in some habitats. Vopr Ikhtiol, 21: 1006-1015 |
Brill R W, Bigelow K A, Musyl M K et al, 2005. Bigeye tuna (Thunnus obesus) behavior and physiology and their relevance to stock assessments and fishery biology. Collective Volume of Scientific Papers ICCAT, 57(2): 142-161 |
FAO, 2018. FAO yearbook. Fishery and Aquaculture Statistics 2016/FAO annuaire. Rome, 1-104
|
Hasler A D, 1975. Coupling of Land and Water Systems. Berlin Heidelberg: Springer-Verlag, 129-136
|
Hyslop E J, 1980. Stomach contents analysis——a review of methods and their application. Journal of Fish Biology, 17(4): 411-429 DOI:10.1111/jfb.1980.17.issue-4 |
Mariotti A, 1983. Atmospheric nitrogen is a reliable standard for natural 15N abundance measurements. Nature, 303(5919): 685-687 DOI:10.1038/303685a0 |
Marshall S, Elliott M, 1997. A comparison of univariate and multivariate numerical and graphical techniques for determining inter- and intraspecific feeding relationships in estuarine fish. Journal of Fish Biology, 51(3): 526-545 DOI:10.1111/j.1095-8649.1997.tb01510.x |
Ohshimo S, Hiraoka Y, Sato T et al, 2018. Feeding habits of bigeye tuna (Thunnus obesus) in the North Pacific from 2011 to 2013. Marine and Freshwater Research, 69(4): 585-606 DOI:10.1071/MF17058 |
Post D M, 2012. Using stable isotopes to estimate trophic position:models, methods, and assumptions. Ecology, 83(3): 703-718 |
Potier M, Marsac F, Cherel Y et al, 2007. Forage fauna in the diet of three large pelagic fishes (lancetfish, swordfish and yellowfin tuna) in the western equatorial Indian Ocean. Fisheries Research, 83(1): 60-72 DOI:10.1016/j.fishres.2006.08.020 |
Vaske Júnior T, Travassos P E, Hazin F H V et al, 2012. Forage fauna in the diet of bigeye tuna (Thunnus obesus) in the western tropical Atlantic Ocean. Brazilian Journal of Oceanography, 60(1): 89-97 DOI:10.1590/S1679-87592012000100009 |
Zanden M J V, Rasmussen J B, 2001. Variation in δ15N and δ13C trophic fractionation:implications for aquatic food web studies. Limnology and Oceanography, 46(8): 2061-2066 DOI:10.4319/lo.2001.46.8.2061 |