 2019, Vol. 50
2019, Vol. 50

黑鲷、真鲷及其杂交子代基因编码区微卫星序列及密码子偏好性分析

http://dx.doi.org/10.11693/hyhz20190200038
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 曹广勇, 张志勇, 张志伟, 陈淑吟, 祝斐, 贾超峰, 陈自强, 曾海峰, 汤晓建. 2019.
- CAO Guang-Yong, ZHANG Zhi-Yong, ZHANG Zhi-Wei, CHEN Shu-Yin, ZHU Fei, JIA Chao-Feng, CHEN Zi-Qiang, ZENG Hai-Feng, TANG Xiao-Jian. 2019.
- 黑鲷、真鲷及其杂交子代基因编码区微卫星序列及密码子偏好性分析
- ANALYSIS OF THE MICROSATELLITE SEQUENCES AND CODON BIAS OF THE CODING SEQUENCE IN ACANTHOPAGRUS SCHLEGELII, PAGRUS MAJOR AND THEIR HYBRID PROGENIES
- 海洋与湖沼, 50(5): 1108-1115
- Oceanologia et Limnologia Sinica, 50(5): 1108-1115.
- http://dx.doi.org/10.11693/hyhz20190200038
文章历史
-
收稿日期:2019-02-22
收修改稿日期:2019-05-05
引用本文
曹广勇, 张志勇, 张志伟, 陈淑吟, 祝斐, 贾超峰, 陈自强, 曾海峰, 汤晓建. 2019. 黑鲷、真鲷及其杂交子代基因编码区微卫星序列及密码子偏好性分析[J]. 海洋与湖沼, 50(5): 1108-1115

CAO Guang-Yong, ZHANG Zhi-Yong, ZHANG Zhi-Wei, CHEN Shu-Yin, ZHU Fei, JIA Chao-Feng, CHEN Zi-Qiang, ZENG Hai-Feng, TANG Xiao-Jian. 2019. ANALYSIS OF THE MICROSATELLITE SEQUENCES AND CODON BIAS OF THE CODING SEQUENCE IN ACANTHOPAGRUS SCHLEGELII, PAGRUS MAJOR AND THEIR HYBRID PROGENIES[J]. Oceanologia et Limnologia Sinica, 50(5): 1108-1115.
黑鲷、真鲷及其杂交子代基因编码区微卫星序列及密码子偏好性分析
1. 上海海洋大学水产与生命学院 上海 201306;
2. 江苏省海洋水产研究所 江苏省海水鱼类遗传育种重点实验室 南通 226007
2. 江苏省海洋水产研究所 江苏省海水鱼类遗传育种重点实验室 南通 226007
摘要:本研究采用生物信息学方法分析比较了黑鲷、真鲷及其两种杂交子代基因编码区(Coding sequence,CDS)中微卫星序列(简单重复序列,SSR)的分布规律及密码子偏好性。结果表明:(1)黑鲷、真鲷及其两种杂交子代微卫星序列均以三碱基重复类型为主,其次为二碱基重复,片段长度大多数在12-20bp之间。(2)黑鲷与反交(黑鲷♀×真鲷♂)在三、五碱基中优势基元类型相同,真鲷与正交(真鲷♀×黑鲷♂)在三碱基中优势基元类型相同,而黑鲷与正交在四碱基中优势基元类型相同。(3)UUC、UAC等28个密码子为黑鲷、真鲷及其两种杂交子代基因编码区的偏好性密码子,偏爱使用以C或G结尾的密码子,且第三位密码子碱基的GC含量(GC3s)均高于基因编码区平均GC含量。(4)真鲷与正交的密码子使用的偏好性相似,一定程度上说明黑鲷、真鲷杂交子代偏母系遗传。分析结果将为研究鲷科鱼类微卫星标记、研究其群体遗传多样性、构建转基因表达系统和开展分子遗传育种提供理论基础。
关键词:黑鲷 真鲷 CDS序列 微卫星 密码子偏好性
ANALYSIS OF THE MICROSATELLITE SEQUENCES AND CODON BIAS OF THE CODING SEQUENCE IN ACANTHOPAGRUS SCHLEGELII, PAGRUS MAJOR AND THEIR HYBRID PROGENIES
1. Fishery and Life Science College, Shanghai Ocean University, Shanghai 201306, China;
2. Jiangsu Marine Fisheries Research Institute, Nantong 226007, China
2. Jiangsu Marine Fisheries Research Institute, Nantong 226007, China
Abstract: The microsatellite sequences (simple sequence repeats, SSR) and codon bias in the coding regions (CDS) of black porgy (Acanthopagrus schlegelii), red porgy (Pagrus major), and their offspring (orthogonal and reciprocal ones) were analyzed and compared using the method of bio-informatics. The results are followed. The microsatellite sequence is mainly composed of the three-base repeat type, followed by the two-base repeat, and the most fragment length of repeat sequences is between 12bp and 20bp. The black porgy and the reciprocal cross (black porgy♀×red porgy ♂) have the same dominant primitive type in the three and five bases, and the red porgy and the orthogonal (red porgy♀×black porgy ♂) dominant base types in the three bases as well, while the black porgy and the orthogonal are the same in the four bases. Twenty-eight pieces of genetic codons such as UUC and UAC are the preferred codons in gene coding regions of the four sea bream species, preferring to use the codons ended in C or G, and the GC content of the third codon base (GC3s). Both are higher than the average GC content of the coding region of the gene. The preference of codon use of the red porgy and the orthogonal is similar, which to some degree shows that black porgy and red porgy hybrids are of maternal hereditary characteristics. This research may provide a theoretical basis for the study of microsatellite markers of sea bream, their population genetic diversity, the construction of a transgenic expression system, and molecular genetic breeding.
Key words:
Acanthopagrus schlegelii Pagrus major coding sequence microsatellite codon usage bias
黑鲷(Acanthopagrus schlegelii)和真鲷(Pagrus major)均隶属鲈形目(Percoiformes)、鲷科(Sparidae), 是我国沿海重要的海水养殖经济鱼类。真鲷个体大且生长快, 而黑鲷营养价值高、抗逆性强。杂交选育可以使杂种后代增加变异性和异质性, 综合双亲的优良性状, 进而获得优良性状稳定的新品种, 是鱼类育种的有效途径之一。目前, 黑鲷、真鲷及其杂交子代对比分析已有相关报道, 例如:黑鲷和真鲷杂交子代脂肪含量明显高于亲本黑鲷, 具有较好的温度、盐度及低氧抗逆性(邱新志等, 2008; 蒋宏雷等, 2009; 姜景腾等, 2010; 王思婷等, 2016); 真鲷(♀)×黑鲷(♂)杂交子一代偏母系遗传, 且与真鲷的Calmodulin功能基因的组织表达有所差异(林勉等, 2014; 陈淑吟等, 2017), 而在基因编码序列(Coding sequence, CDS)生物信息分析方面尚未见报道。
微卫星特征及密码子偏好性分析是在基因组水平进行生物信息分析的两种常见方法。在真核生物基因组中, 高等生物基因组倾向于单碱基微卫星类型占主导地位(黄杰等, 2015), 将单碱基类型除外, 则以二碱基微卫星序列最为丰富(Ma et al, 1996); 而在水生生物中, 出现以二碱基重复和三碱基重复为主的两种现象(崔建洲等, 2006; 李偲等, 2011; 刘博等, 2012; 袁文成等, 2015)。此外, 生物体内编码某种氨基酸会出现倾向特定密码子的现象, 即密码子偏好性(codon bias), 一般相近物种的密码子使用模式相似(Lynn et al, 2002), 而不同物种的密码子偏好性也有所差异。密码子的这种偏好性是物种进化过程中基因突变和选择的结果, 基因中的GC含量以及密码子第3位上GC含量(GC3S)可能为影响密码子偏性的主要因素(Adams et al, 2003; Romero et al, 2003)。本文采用生物信息学工具在基因编码序列水平上研究黑鲷、真鲷及其两种杂交子代微卫星序列的分布规律及密码子偏好性, 为鲷科鱼类构建转基因表达系统、提高特定基因的表达效率、分子遗传育种、种质资源保护和开发利用提供参考。
1 材料与方法 1.1 数据来源本课题组于2016年对黑鲷、真鲷及其两种杂交子代(正交:真鲷♀×黑鲷♂; 反交:黑鲷♀×真鲷♂)进行转录组测序, 实验用鱼来源于吕四养殖基地, 取样均为1月龄仔鱼, 以全鱼进行测序比对并获得该4种鱼CDS序列数据。所有序列数据均以FASTA文件格式保存。
1.2 SSR位点搜索利用Krait (Du et al, 2018)搜索和统计黑鲷、真鲷及其两种杂交子代CDS序列中的SSR位点。搜索参数分别设置为:单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸、六核苷酸的最少重复次数分别为10、6、5、4、4、4。Perl语言操作系统下MISA软件(MISA, MIcroSAtellite identification tool) (Lu et al, 2013)检验。
1.3 密码子分析指标和方法相对密码子使用度RSCU (Relative Synonymous Codon Usage), 是指对于某一特定的密码子在编码对应氨基酸的同义密码子间的相对概率, 可直观地反映密码子使用的偏好性。当某一密码子RSCU=1, 代表该密码子的使用没有偏好性; RSCU > 1时, 表明该密码子为使用相对较多的密码子, 反之亦然。第i个氨基酸的第j个密码子的RSCU的计算公式(吴宪明等, 2007)如下:
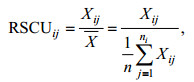
式中, Xij是编码第i个氨基酸的第j个密码子的出现次数, ni是编码第i个氨基酸的同义密码子的数量(值为1—6)。
用CodonW对黑鲷、真鲷及其两种杂交子代CDS序列分别进行分析, 得出每条序列中各密码子的RSCU以及密码子的第3位碱基中A、U、G、C的含量在第3位碱基总量中所占的比率(分别为A3s、U3s、G3s、C3s); 并使用Microsoft Excel 2003和SPSS16.0对黑鲷、真鲷及其两种杂交子代密码子偏好性数据进行统计及聚类分析。
2 结果与分析 2.1 微卫星特征分析 2.1.1 CDS序列中SSR的数量和分布规律黑鲷、真鲷及其两种杂交子代中共检索到185532条CDS序列, GC含量约为53%, 14225个完整型SSR位点(黑鲷: 3613, 真鲷: 3597, 正交: 3315, 反交: 3700), 分布在12709条CDS序列(黑鲷: 3215, 真鲷: 3172, 正交: 2996, 反交: 3326), SSR发生频率为6.9%(黑鲷: 8.5%, 真鲷: 7.8%, 正交: 6.8%, 反交: 5.3%)。经对黑鲷、真鲷及其两种杂交子代CDS序列中不同碱基完整型SSR进行分析, 每种鱼的具体SSR信息见表 1。
表 1 黑鲷、真鲷及其两种杂交子代CDS序列中SSR位点的搜索结果 Tab. 1 Results of SSR search in coding sequence of four sea-bream
| 序号 | 项目 | 黑鲷 | 真鲷 | 正交 | 反交 |
| 1 | 序列总数量 | 37887 | 40641 | 44152 | 62852 |
| 2 | 序列总长度(bp) | 36848430 | 37735593 | 37773645 | 48081141 |
| 3 | GC含量(%) | 53.92 | 53.49 | 53.7 | 52.53 |
| 4 | 检测到的完美型SSR数量 | 3613 | 3597 | 3315 | 3700 |
| 5 | 完美型SSR总长度(bp) | 57689 | 58025 | 52856 | 58509 |
| 6 | SSRs的平均长度(bp) | 15.97 | 16.14 | 15.95 | 15.82 |
| 7 | SSR占比(%) | 0.16 | 0.16 | 0.14 | 0.13 |
| 8 | 相对丰度(loci/Mb) | 98.05 | 95.32 | 87.76 | 76.95 |
| 9 | 相对密度(bp/Mb) | 1565.58 | 1537.67 | 1399.28 | 1216.88 |
| 10 | 单碱基重复SSR数目(%) | 257(7.11%) | 202(5.62%) | 142(4.28%) | 226(6.11%) |
| 11 | 二碱基重复SSR数目(%) | 550(15.22%) | 514(14.29%) | 515(15.54%) | 572(15.46%) |
| 12 | 三碱基重复SSR数目(%) | 2666(73.79%) | 2724(75.73%) | 2529(76.29%) | 2748(74.27%) |
| 13 | 四碱基重复SSR数目(%) | 69(1.91%) | 66(1.83%) | 62(1.87%) | 86(2.32%) |
| 14 | 五碱基重复SSR数目(%) | 18(0.5%) | 19(0.53%) | 15(0.45%) | 23(0.62%) |
| 15 | 六碱基重复SSR数目(%) | 53(1.47%) | 72(2.0%) | 52(1.57%) | 45(1.22%) |
在SSR位点各碱基重复数量中, 三碱基重复在黑鲷、真鲷及其两种杂交子代CDS序列中占比均为最大(约75%), 二碱基重复其次(约15%), 五碱基重复占比最小, 约为0.5% (表 1)。
黑鲷微卫星81种基元(motif)类型, 真鲷微卫星94种基元类型, 正交80种基元类型, 反交79种基元类型。黑鲷、真鲷及其两种杂交子代CDS序列中二、三碱基重复数量约占总SSR位点90%, 其中二、三碱基重复类型分别以AC、AG、AAC、ACC、ACG、AAG重复基元分布最多, 这表明CDS序列中微卫星的分布对A/T碱基不具有偏好性。
黑鲷、真鲷及其两种杂交子代在单碱基和二碱基重复的优势基元类型上完全一致(单碱基为A/T, 二碱基为AC/GT)。黑鲷与反交在三、五碱基中优势基元类型相同, 真鲷与正交在三碱基中优势基元类型相同, 而黑鲷与正交在四碱基中优势基元类型相同。该4种鱼优势基元类型在单碱基和二碱基重复中的占比较为接近, 黑鲷与反交在三碱基重复类型中的优势基元占比较为接近, 而黑鲷与正交在四碱基中优势基元类型相同。黑鲷、真鲷及其两种杂交子代在四碱基重复的优势基元类型及占比差异较大。该4种鱼CDS序列中优势重复碱基具体信息见表 2。
表 2 黑鲷、真鲷及其两种杂交子代CDS序列中优势重复碱基的组成和频率统计(考虑到碱基互补作用) Tab. 2 Composition and frequency of the dominant repeat motifs in the coding sequence of four sea-bream (in consideration of sequence complementarity)
| 重复类型 | 黑鲷 | 真鲷 | 正交 | 反交 |
| 单碱基重复 | 225 (87.5%) [A/T] |
173 (85.6%) [A/T] |
119 (83.8%) [A/T] |
192 (85.0%) [A/T] |
| 二碱基重复 | 309(56.2%) [AC/GT] |
304(59.1%) [AC/GT] |
297(57.7%) [AC/GT] |
316(55.2%) [AC/GT] |
| 三碱基重复 | 760(28.5%) [ACG/CGT] |
280(10.3%) [AAG/CTT] |
295(11.7%) [AAG/CTT] |
767(27.9%) [ACG/CGT] |
| 四碱基重复 | 20(29.0%) [ACAG/CTGT] |
4(6.1%) [AAAG/CTTT] |
14(22.6%) [ACAG/CTGT] |
12(14.0%) [AAAC/GTTT] |
| 五碱基重复 | 4(22.2%) [AAAAC/GTTTT] |
3(15.8%) [ACCGC/GCGGT] |
/ / |
3(13.0%) [AAAAC/GTTTT] |
| 六碱基重复 | / / |
2(2.8%) [AACCAG/CTGGTT] |
3(5.8%) [AACAGG/CCTGTT] |
/ / |
黑鲷、真鲷及其两种杂交子代CDS序列中微卫星长度分布和数量与片段长度的分布关系结果相似。以黑鲷为例, 微卫星长度分布规律和数量与片段长度的分布关系如图 1、图 2所示。黑鲷CDS序列中所发现的3613个完整型微卫星中, 微卫星长度差异不大, 片段长度从10—30bp不等, 平均长度为15.97bp。黑鲷微卫星序列中, 主要是重复长度大于15bp的重复序列, 其中长度为15bp的三碱基重复数量最多(1585条), 占微卫星(完整型)总数的43.9%;长度为18bp的三碱基重复次之(807条), 占微卫星(完整型)总数的22.3%(图 1)。
 |
| 图 1 黑鲷CDS序列中微卫星的长度分布 Fig. 1 Length distribution of microsatellites in the coding sequence of A. schlegelii |
 |
| 图 2 黑鲷CDS序列中SSR数量随重复次数的变化 Fig. 2 Variation of repeat times with the number of microsatellites in the coding sequence of A. schlegelii |
从图 2可以看出, 黑鲷SSR数量随着重复次数的增加呈明显下降趋势。三碱基重复随重复次数的增加, SSR数量呈直线下降, 当重复次数从5次增加到8次时, 黑鲷SSR数量下降至0, 之后稳定。三碱基重复SSR数量则随重复次数增加而减少。当二碱基重复次数达到10次、单碱基重复次数达到12次、其他碱基重复曲线均较为平缓。
2.2 密码子偏好性 2.2.1 黑鲷、真鲷及其两种杂交子代基因编码区碱基组成黑鲷基因编码区第三位密码子的碱基GC3s含量最高(60.88%), 反交最低(58.32%), 可见黑鲷、真鲷及其两种杂交子代的编码蛋白基因序列偏爱使用碱基以GC结尾的密码子。从第三位密码子碱基组成看, 黑鲷等4种鱼基因编码区A、U、G和C四种碱基结尾的密码子以C结尾的密码子最多, 其他依次为G3s、U3s和A3s(表 3)。由表 1与表 3对比还可以发现, 黑鲷、真鲷及其两种杂交子代第三位密码子碱基的GC含量(GC3s)均高于基因编码区平均GC含量, 而第三位密码子也是导致密码子使用发生偏向性的重要因素之一。
表 3 黑鲷、真鲷及其两种杂交子代CDS序列密码子的A3s、U3s、G3s、C3s Tab. 3 Values of A3s, U3s, G3s, and C3s in the coding sequences of four sea-bream
| 物种 | A3s | U3s | G3s | C3s |
| 黑鲷 | 18.75 | 20.37 | 27.66 | 33.22 |
| 真鲷 | 19.19 | 20.97 | 27.12 | 32.72 |
| 正交 | 19.01 | 20.65 | 27.41 | 32.92 |
| 反交 | 19.86 | 21.82 | 26.88 | 31.44 |
| 注: A3s, U3s, G3s和C3s分别为A、U、G和C四种碱基结尾的密码子含量(百分比%) | ||||
UUC、UAC、CAC、CAG等28个密码子的RSCU均大于1, 为黑鲷编码蛋白质基因的偏好性密码子(表 4), 黑鲷与真鲷、正交、反交的编码蛋白基因的偏好性密码子完全相同, 其中尤其偏爱使用以C结尾的密码子(除CGC外, 其余的都为偏好性密码子), 其次为G结尾的密码子, 特别是CUG, 它们的RSCU均达到2.5以上, 而以A或U碱基结尾的仅有7个。虽然在黑鲷等4种鱼基因编码区偏爱以C或G结尾的同义密码子, 但NCG形式的四种密码子(N代表A, U, G, C四种碱基中的任一种), 没有一个为偏好性密码子, 且均在其同义密码子中为最小。此外, 以密码子编码氨基酸的原理计算, 黑鲷、真鲷及其两种杂交子代的编码20种氨基酸中丝氨酸(Ser)和亮氨酸(Leu)含量最高(如图 3所示), 该两种氨基酸的偏好密码子均以G/C结尾。
表 4 黑鲷、真鲷及其两种杂交子代CDS序列偏好密码子的RSCU Tab. 4 RSCU of preferable codons of coding sequences in four sea-bream
| 氨基酸 | 密码子 | 黑鲷 | 真鲷 | 正交 | 反交 |
| 苯丙氨酸 | UUC | 1.25 | 1.22 | 1.23 | 1.18 |
| 酪氨酸 | UAC | 1.39 | 1.37 | 1.38 | 1.31 |
| 组氨酸 | CAC | 1.31 | 1.29 | 1.31 | 1.25 |
| 谷氨酰胺 | CAG | 1.52 | 1.51 | 1.52 | 1.49 |
| 天冬酰胺 | AAC | 1.39 | 1.37 | 1.38 | 1.30 |
| 赖氨酸 | AAG | 1.17 | 1.15 | 1.16 | 1.16 |
| 天冬氨酸 | GAC | 1.27 | 1.25 | 1.26 | 1.20 |
| 谷氨酸 | GAG | 1.41 | 1.40 | 1.40 | 1.35 |
| 半胱氨酸 | UGC | 1.06 | 1.03 | 1.04 | 1.01 |
| 亮氨酸 | CUC | 1.33 | 1.32 | 1.32 | 1.26 |
| CUG | 2.70 | 2.65 | 2.67 | 2.50 | |
| 丝氨酸 | UCU | 1.01 | 1.04 | 1.03 | 1.08 |
| UCC | 1.32 | 1.30 | 1.31 | 1.31 | |
| AGC | 1.46 | 1.43 | 1.44 | 1.36 | |
| 精氨酸 | AGA | 1.46 | 1.50 | 1.48 | 1.61 |
| AGG | 1.57 | 1.56 | 1.57 | 1.60 | |
| 缬氨酸 | GUC | 1.17 | 1.15 | 1.15 | 1.10 |
| GUG | 1.77 | 1.76 | 1.76 | 1.70 | |
| 脯氨酸 | CCU | 1.14 | 1.18 | 1.16 | 1.18 |
| CCC | 1.13 | 1.12 | 1.12 | 1.11 | |
| CCA | 1.09 | 1.11 | 1.10 | 1.10 | |
| 苏氨酸 | ACC | 1.36 | 1.34 | 1.35 | 1.32 |
| ACA | 1.15 | 1.19 | 1.17 | 1.18 | |
| 丙氨酸 | GCU | 1.14 | 1.18 | 1.16 | 1.17 |
| GCC | 1.40 | 1.38 | 1.39 | 1.36 | |
| 甘氨酸 | GGC | 1.20 | 1.18 | 1.19 | 1.10 |
| GGA | 1.22 | 1.23 | 1.23 | 1.29 | |
| 异亮氨酸 | AUC | 1.76 | 1.73 | 1.74 | 1.58 |
| 注:色氨酸和甲硫氨酸的RSCU为1, 终止密码子未列入表中 | |||||
 |
| 图 3 黑鲷、真鲷及其两种杂交子代各种氨基酸含量 Fig. 3 The content of a variety of amino acid residues of coding sequences in four sea-bream 注:氨基酸理论含量以密码子编码氨基酸的原理计算 |
在对基因密码子使用概率进行聚类分析的过程中, 将每种鱼的基因编码区序列作为一个对象, 将偏好性密码子的RSCU作为变量。除去RSCU始终为1的编码色氨酸(Trp)和甲硫氨酸(Met)的密码子UGG和AUG, 再除去3个不编码氨基酸的终止密码子, 取余下的59个密码子的RSCU对基因的密码子使用偏好性进行分析。聚类结果见图 4:真鲷与正交首先聚为一类, 然后与黑鲷聚为一类。说明真鲷与正交的密码子使用的偏好性相似, 也一定程度表明黑鲷和真鲷杂交子代偏母系遗传。
 |
| 图 4 黑鲷、真鲷及其两种杂交子代的CDS序列的基因密码子偏好性聚类 Fig. 4 Cluster tree based on the RSCU values of coding sequences in four sea-bream |
黑鲷、真鲷、正交和反交CDS序列SSR的发生频率(含SSR的Unigene数/Unigene总数)分别为8.5%、7.8%、6.8%、5.3%, 平均间隔分别10.2kb、10.5kb、11.4kb、13.0kb出现1个SSR序列。这4种鱼微卫星序列的重复类型中, 均为三碱基重复类型的数量最高(约75%); 二碱基重复类型的数量次之(约15%)。这与翘嘴鳜(Siniperca chuatsi)、缢蛏(Sinonovacula constricta)等以三碱基重复为主的水生生物相同(刘博等, 2012; 袁文成等, 2015), 与草鱼(Ctenopharyngodon idellus)、红鳍东方鲀(Takifugu rubripes)等水生生物以二碱基重复为主不同(崔建洲等, 2006; 李偲等, 2011)。
在所有的微卫星重复类型中, 二核苷酸重复类型最为活跃, 变异水平最高, 拷贝数分布范围最广(白翠翠等, 2016)。两碱基重复类型中, 黑鲷、真鲷及其两种杂交子代均以AC重复拷贝类别最为丰富, 这与真菌类生物的内含子和红鳍东方鲀的研究结果一致(Tóth et al, 2000; 崔建洲等, 2006), 但与中国明对虾(Fenneropenaeus chinensis)的AT、三疣梭子蟹(Portunus trituberculatus)的AG重复拷贝多不同(高焕等, 2004; 宋来鹏等, 2008), 与禾本科植物也不同(郑燕等, 2011)。三碱基重复类型中, 黑鲷和反交以ACG最多, 其次是AAG和ACC, 真鲷和正交以AAG最多, 其次是ACC和ACG。有研究表明, 三碱基重复类型在水生生物中分布特征差异较大, 例如:三疣梭子蟹基因组其中以ACT多, 其次是AGG和AAT(宋来鹏等, 2008); 中国明对虾以AAT多, 其次是AAG和ATC(高焕等, 2004); 红鳍东方鲀中以AAT多, 其次是AGG和ATC, GCC少(崔建洲等, 2006)。同时有研究发现, 二碱基重复引物的多态性较高, 多应用于种群间遗传多样性分析(房祖业等, 2018); 一些三碱基重复与某些遗传性病变密切相关(Pearson et al, 1998)。这4种鱼与上述几种物种在二碱基重复类型中均缺少GC重复, 可能由于DNA CpG的甲基化, 胞苷酸(C)很容易经过脱氨基作用转变成胸腺嘧啶(T), 导致GC重复减少, 剩余少量GC维持DNA热力学稳定性所必需, 而突变后的序列TG(即AC类型)相应增加(Schorderet et al, 1992)。
SSR分子标记多态性的高低取决于其长度大小, 可以判断其潜在利用价值(李珊等, 2010)。当SSR长度为12—20bp时, 呈中度多态性; 长度为20bp以上时, 表示多态性较高; 长度为12bp以下时, 则呈低多态性(Temnykh et al, 2001)。本研究中在SSR位点搜索时已将长度在10bp以下的SSR序列筛除, 发现黑鲷、真鲷及其两种杂交子代CDS序列中完整型SSR长度均在10—85个碱基之间, 片段长度在12—20bp之间的SSR序列最多(约占各SSR总数的85%), 表明该4种鱼CDS序列来源的SSR具有中等的多态性潜能, 在开发黑鲷等微卫星分子标记研究方面将具有一定程度的利用价值。
3.2 密码子偏好性分析黑鲷、真鲷、正交及反交4种鱼的基因编码区在密码子使用上呈现高度一致性, 均偏爱C/G结尾的密码子, 确定了28种偏好密码子(RSCU > 1), 其中21种以G/C结尾, 偏好性较强的有CUG (RSCU > 2), 表明该4种鱼具有较近的亲缘关系。而在聚类分析中, 发现真鲷与正交的密码子使用的偏好性相似, 一定程度上说明黑鲷和真鲷杂交子代偏母系遗传, 这与染色体构成和AFLP分析结果一致(林勉等, 2014)。此外, 黑鲷、真鲷及其两种杂交子代的编码20种氨基酸中丝氨酸(Ser)和亮氨酸(Leu)含量最高, 且两种氨基酸的偏好密码子均以G/C结尾。该两种氨基酸分别在维持免疫系统和控制血糖具有重要作用, 间接表明鲷科鱼类具有较高营养价值。密码子偏好性分析也一定程度上反映了黑鲷等4种鱼具有较高的营养价值。
在进化过程中物种的密码子使用特点具有保守性, 不同的物种对密码子的使用偏好性有所差异, 相近物种的密码子使用模式相似(Lynn et al, 2002)。本研究发现, 黑鲷、真鲷、正交及反交4种鱼的基因编码区的GC含量, 尤其是第3位密码子的GC含量都较高, 这一特点与金鱼(Carassius auratus)的基因组编码区密码子组成相似(张欣等, 2017), 与鲤(Cyprinus carpio)和斑马鱼(Danio rerio)HOX基因家族密码子组成存在差异(周丹等, 2013)。通过优化与改造密码子, 可促进翻译效率及提高生物中外源基因的表达, 例:黑鲷偏好密码子的基因作为外源基因, 而在真鲷、金鱼等物种中获得高效表达; 若依据黑鲷的密码子偏好性信息进行密码子改造, 可提高以黑鲷偏好密码子的外源基因在其他模式生物中的表达效率。对于今后鲷科鱼类转基因过程中对构建合适的转基因表达系统具有重要的指导意义, 从而提高特定蛋白的表达量, 增强转基因育种的效果。然而, 关于黑鲷、真鲷及其两种杂交子代4种鱼基因编码区密码子偏好使用对功能基因保守结构域的影响及其蛋白所携带信息的影响还有待进一步研究。
4 结论本研究利用生物信息学软件分析比较了黑鲷、真鲷及其两种杂交子代4种鱼基因编码区(CDS序列)中微卫星序列的分布特征及密码子偏好性, 结果显示:该4种鱼基因编码区均为以三碱基重复为主的微卫星序列并使用相同的偏好性密码子, 各碱基重复中优势基元类型及偏好性密码子的相对密码子使用度有所差异, 进而推断黑鲷和真鲷杂交子代一定程度偏母系遗传。本研究结果将为研究鲷科鱼类微卫星标记、研究其群体遗传多样性、构建转基因表达系统和开展分子遗传育种提供理论基础。
参考文献
王思婷, 李鹏, 陈淑吟, 等. 2016. 黑鲷与真鲷及其杂交子一代脂肪含量测定与脂肪酸成分分析. 食品科学, 37(20): 81-85 DOI:10.7506/spkx1002-6630-201620014 |
白翠翠, 柳淑芳, 庄志猛. 2016. 虾虎鱼科(Gobiidae)基因组微卫星DNA的分布特征. 渔业科学进展, 37(5): 9-15 |
刘博, 邵艳卿, 滕爽爽, 等. 2012. 缢蛏(Sinonovacula constricta)EST-SSR分布特征及引物开发利用. 海洋与湖沼, 43(1): 132-137 |
李珊, 周天华, 赵桂仿, 等. 2010. 马蹄香表达序列标签资源的SSR信息分析. 中草药, 41(3): 464-468 |
李偲, 刘航, 黄容, 等. 2011. 草鱼Ⅰ型微卫星标记的发掘及其多态性检测. 水生生物学报, 35(4): 681-687 |
吴宪明, 吴松锋, 朱云平, 等. 2007. 密码子偏性的分析方法及相关研究进展. 遗传, 29(4): 420-426 DOI:10.3321/j.issn:0253-9772.2007.04.006 |
邱新志, 吴雄飞. 2008. 盐度对真鲷(♀)×黑鲷(♂)杂交子一代仔鱼活力的影响. 渔业现代化, 35(4): 41-43 DOI:10.3969/j.issn.1007-9580.2008.04.010 |
宋来鹏, 刘萍, 李健, 等. 2008. 三疣梭子蟹基因组微卫星特征分析. 中国水产科学, 15(5): 738-744 DOI:10.3321/j.issn:1005-8737.2008.05.004 |
张欣, 尤宏争, 李荣妮, 等. 2017. 金鱼密码子用法特点分析. 华北农学报, 32(S1): 30-36 DOI:10.7668/hbnxb.2017.S1.006 |
陈淑吟, 李鹏, 张志勇, 等. 2017. 黑鲷♀×真鲷♂反交子代与黑鲷的CaM基因克隆与mRNA表达分析. 中国水产科学, 24(6): 1193-1202 |
林勉, 苗亮, 李明云, 等. 2014. 真鲷、黑鲷及其杂交子代的染色体构成与AFLP分析. 海洋学报, 36(8): 117-121 DOI:10.3969/j.issn.0253-4193.2014.08.013 |
周丹, 薛仁余, 张晓峰, 等. 2013. 鲤和斑马鱼HOX基因家族同义密码子使用偏性的分析. 水产学杂志, 26(2): 19-25 DOI:10.3969/j.issn.1005-3832.2013.02.004 |
郑燕, 张耿, 吴为人. 2011. 禾本科植物微卫星序列的特征分析和比较. 基因组学与应用生物学, 30(5): 513-520 DOI:10.3969/gab.030.000513 |
房祖业, 陈晓东, 吴咏诗, 等. 2018. 大刺鳅(Mastacembelus armatus)二、三、四碱基重复微卫星标记的筛选和特征分析. 海洋与湖沼, 49(1): 174-182 |
姜景腾, 吴雄飞, 蒋宏雷. 2010. 低氧胁迫对真鲷(♀)与黑鲷(♂)杂交子一代体内酶活力的影响. 宁波大学学报(理工版), 23(4): 10-14 DOI:10.3969/j.issn.1001-5132.2010.04.003 |
袁文成, 黄鹤忠, 李文龙, 等. 2015. 翘嘴鳜(Siniperca chuatsi)转录组EST-SSR位点的信息分析及其多态性检测. 海洋与湖沼, 46(2): 403-409 |
高焕, 刘萍, 孟宪红, 等. 2004. 中国对虾(Fenneropenaeus chinensis)基因组微卫星特征分析. 海洋与湖沼, 35(5): 424-431 DOI:10.3321/j.issn:0029-814X.2004.05.005 |
黄杰, 周瑜, 刘与之, 等. 2015. 基于454 GS FLX高通量测序的四川山鹧鸪基因组微卫星特征分析. 四川动物, 34(1): 8-14 |
崔建洲, 申雪艳, 杨官品, 等. 2006. 红鳍东方鲀基因组微卫星特征分析. 中国海洋大学学报, 36(2): 249-254 |
蒋宏雷, 吴雄飞. 2009. 真鲷与黑鲷及杂交F1耐受温度、生长比较. 现代渔业信息, 24(2): 26-28 DOI:10.3969/j.issn.1004-8340.2009.02.008 |
Adams M J, Antoniw J F, 2003. Codon usage bias amongst plant viruses. Archives of Virology, 149(1): 113-135 DOI:10.1007/s00705-003-0186-6 |
Du L M, Zhang C, Liu Q et al, 2018. Krait:an ultrafast tool for genome-wide survey of microsatellites and primer design. Bioinformatics, 34(4): 681-683 DOI:10.1093/bioinformatics/btx665 |
Lu X, Wang H X, Liu B Z et al, 2013. Three EST-SSR markers associated with QTL for the growth of the clam Meretrix meretrix revealed by selective genotyping. Marine Biotechnology, 15(1): 16-25 DOI:10.1007/s10126-012-9453-4 |
Lynn D J, Singer G A C, Hickey D A, 2002. Synonymous codon usage is subject to selection in thermophilic bacteria. Nucleic Acids Research, 30(19): 4272-4277 DOI:10.1093/nar/gkf546 |
Ma Z Q, Röder M, Sorrells M E, 1996. Frequencies and sequence characteristics of di-, tri-, and tetra-nucleotide microsatellites in wheat. Genome, 39(1): 123-130 DOI:10.1139/g96-017 |
Pearson C E, Sinden R, 1998. Trinucleotide repeat DNA structures:dynamic mutations from dynamic DNA. Current Opinion in Structural Biology, 8(3): 321-330 DOI:10.1016/S0959-440X(98)80065-1 |
Romero H, Zavala A, Musto H et al, 2003. The influence of translational selection on codon usage in fishes from the family Cyprinidae. Gene, 317: 141-147 DOI:10.1016/S0378-1119(03)00701-7 |
Schorderet D F, Gartler S M, 1992. Analysis of CpG suppression in methylated and nonmethylated species. Proceedings of the National Academy of Sciences of the United States of America, 89(3): 957-961 DOI:10.1073/pnas.89.3.957 |
Tó th, Gáspári Z, Jurka J, 2000. Microsatellites in different eukaryotic genomes:survey and analysis. Genome Research, 10(7): 967-981 DOI:10.1101/gr.10.7.967 |
Temnykh S, DeClerck G, Lukashova A et al, 2001. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.):frequency, length variation, transposon associations, and genetic marker potential.. Genome Research, 11(8): 1441-1452 DOI:10.1101/gr.184001 |