 2020, Vol. 51
2020, Vol. 51中国海洋湖沼学会主办。
文章信息
- 骆云慧, 李来国, 赵淳朴, 王丹丽, 徐善良, 徐继林. 2020.
- LUO Yun-Hui, LI Lai-Guo, ZHAO Chun-Pu, WANG Dan-Li, XU Shan-Liang, XU Ji-Lin. 2020.
- 凡纳滨对虾-缢蛏生态循环养殖池塘中浮游植物群落结构与水质因子相关性研究
- RELATIONSHIP BETWEEN PHYTOPLANKTON STRUCTURE AND WATER QUALITY FACTORS IN CULTURE PONDS OF LITOPENAEUS VANNAMEI AND SINONOVACULA CONSTRICTA
- 海洋与湖沼, 51(2): 378-387
- Oceanologia et Limnologia Sinica, 51(2): 378-387.
- http://dx.doi.org/10.11693/hyhz20191100231
文章历史
-
收稿日期:2019-11-27
收修改稿日期:2019-12-29




2. 宁波大学海洋生物工程浙江省重点实验室 宁波 315211;
3. 鄞州瞻岐椿霖水产养殖场 宁波 315145
2. Zhejiang Key Laboratory of Marine Bioengineering, Ningbo University, Ningbo 315211, China;
3. Yinzhou Zhanqi Chunlin Aquatic Farm, Ningbo 315145, China
凡纳滨对虾(Litopenaeus vannamei)具有生长快、繁殖期长、抗病能力强、便于活虾运输等优点(Rodríguez et al, 2007), 凡纳滨对虾在全世界范围内的对虾养殖品种中, 其养殖面积最大且养殖产量最高(周井娟, 2016), 是我国水产业中重要的养殖品种。缢蛏(Sinonovacula constricta)属于双壳纲(Bivalvia)、帘蛤目(Veneroida)、截蛏科(Solecurtidae)的滩涂贝类, 具有生长速度快、适应能力强、经济价值高等特点, 在中国沿海各地均已开展养殖, 是我国四大养殖贝类之一。
凡纳滨对虾与缢蛏循环养殖模式是以“独立式供饵式缢蛏养殖模式”(刘招坤, 2014)为基础发展出来的一种新型的海水生态养殖模式, 体现了池塘循环水养殖模式与综合养殖模式的优点。通过对养殖池塘进行改造, 将凡纳滨对虾和缢蛏分别放养在改造后的相邻池塘中, 由对虾养殖池塘内人工投喂的残饵和对虾粪便等代谢物转化而来的营养盐, 被虾池中微藻充分利用, 使养虾池塘中浮游藻类维持较高的种群优势后通过水泵注入贝类养殖塘, 贝类滤食从对虾塘带来的丰富藻类、有机碎屑和菌团等, 滤清后利用水位落差重新流回对虾养殖塘, 使两个养殖池塘之间形成功能互补关系, 构建虾蛏循环套养系统, 从而达到净化水质、循环利用海水的目的。
浮游植物作为水生态系统中重要的初级生产者, 其种类和数量的变化直接或间接地影响其他水生生物的丰度和分布, 甚至影响整个生态系统的稳定(Davis et al, 1998; 胡韧等, 2002)。由于浮游植物生长周期短, 对水环境的变化敏感而迅速, 其生物量和群落结构能很好地反映水质现状及变化趋势, 是评价水环境质量的重要指标(Rott et al, 2006; Sidik et al, 2008)。而环境中的各种因素, 如温度、盐度、pH值、氮磷营养盐及水中的生物均会影响浮游植物种类及数量变化, 浮游植物种类及数量也会对各水质因子产生影响(王旭娜等, 2018)。因此, 了解凡纳滨对虾-缢蛏循环养殖池塘浮游植物群落特征, 研究水质因子和浮游植物相关性, 对维持养殖池塘生态系统平衡有重要作用。
1 材料与方法 1.1 实验条件实验地点位于浙江省宁波市鄞州区瞻岐镇椿霖水产养殖场。实验前改造池塘, 按照虾蛏循环套养模式, 将两个原有生产池塘改造成两个面积约为3000m2的缢蛏养殖塘(TC-1, TC-2)和两个面积约为12000m2的凡纳滨对虾养殖塘(TX-1, TX-2), 缢蛏养殖塘与凡纳滨对虾养殖塘之间设置高度落差, 使得缢蛏养殖塘中水体能够自流至对虾养殖塘, 安装水泵用于抽取对虾养殖塘的水体至缢蛏养殖塘中。缢蛏养殖池塘使用水车式增氧机增氧, 凡纳滨对虾养殖塘使用纳米管底增氧。
实验时间为2018年6—11月, 于2018年6月5日投放蛏苗, 蛏苗密度为250ind./m2; 于6月15日、8月5日投放两批虾苗, 虾苗密度分别为50ind./m2、30ind./m2, 凡纳滨对虾于11月2日全部捕捞完毕。
1.2 样品采集与测定在四个试验池塘四边中心距离岸边约2m处及池塘中央共设置五个采样点, 使用采水器采集中层水样各5L, 充分混匀, 取1L混合水样用于水质因子测定, 除亚硝酸盐氮(NO2--N)测定用水样外均加入50%的硫酸(终浓度为1‰), 另取1L混合水样用于浮游植物定量分析; 浮游植物定性使用25#浮游生物网(网目孔径为64μm)垂直采集(Xu et al, 2010), 采集的浮游植物定量及定性样品需加入15%鲁哥试剂及4%福尔马林溶液进行样品固定。
实验水样的测定:使用衡欣AZ8403测定温度(T)、溶氧(DO)、pH, 使用盐度计测定盐度(S), 其余水质因子依据海洋调查规范, 分别采用次溴酸钠氧化法测定氨氮(TAN)、重氮-偶氮法测定亚硝酸盐氮(NO2--N)、锌镉还原法测定硝酸盐氮(NO3--N)、抗坏血酸还原磷钼蓝法测定活性磷酸盐(AP)、过硫酸钾氧化法测定总氮(TN)和总磷(TP), 每月采集测定一次。
浮游植物定性使用25#浮游生物网(网目孔径为64μm)从表层到深处以大约25cm/s的速度作“∞”形缓慢的循环往复式拖动(Xu et al, 2010), 其时间大概为2.5min, 将样品转移到白色聚乙烯瓶中, 加5%福尔马林溶液固定。
1.3 数据计算与分析定性样品在光学显微镜下, 依据《中国海藻志》、《水生生物学》等资料对浮游植物进行种属鉴定, 定量样品震荡混匀, 取0.1mL于血细胞计数板内计数, 计数的误差为±10%。
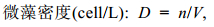 (1)
(1)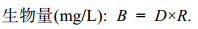 (2)
(2)上述式中: n为计数板所计的微藻个数; V为浓缩后样品的体积。R小型藻 = 2×10–7mg; R中型藻 = 2×10–6mg; R大型藻 = 5×10–6mg (张继民等, 2010)。
浮游植物多样性指数采用Shannon-Wiener物种多样性指数(H′)(沈国英等, 2002):
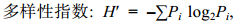 (3)
(3)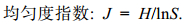 (4)
(4)上述式中均以物种数量作为指标, 其中: Pi = ni/N, ni为物种i的个体数, N为群落样本的总个体数, 即Pi为物种i占总个体数的比例, S为群落物种数; fi表示物种i在该采样点所出现的频率。使用Excel对数据初步处理, 物种优势度经lg(x+1)转换, 理化因子除了pH外, 均经lg(x+1)转换; 利用CANOCO 5软件对虾蛏循环套养池塘优势种和主要理化因子进行CCA分析, 从而获得虾蛏循环套养池塘中影响浮游植物优势种演替的主要生态因子。
2 结果 2.1 水质因子变化养殖池塘水质因子的变化范围见表 1。凡纳滨对虾养殖塘(TX)中总磷(TP)、活性磷酸盐(AP)和硝酸盐氮(NO3--N)含量在养殖期间中呈现明显的先上升后下降的趋势(图 1), 氨氮(TAN)和总氮(TN)含量则是先上升, 后逐渐平稳。凡纳滨对虾养殖塘中亚硝酸盐氮(NO2--N)、总氮(TN)含量始终高于缢蛏养殖塘
| 因子 | TX-1 | TX-2 | TC-1 | TC-2 |
| 温度T (℃) | 22.40—30.50 28.13±3.67 |
22.50—30.70 28.24±3.85 |
21.20—31.80 27.94±4.12 |
21.20—32.60 28.16±4.32 |
| 盐度S | 20.00—25.00 22.25±2.04 |
20.00—24.00 22.85±2.23 |
18.00—23.00 19.23±1.36 |
18.00—23.00 19.56±1.67 |
| 溶解氧DO (mg/L) | 5.17—8.25 6.67±0.47 |
4.98—8.15 6.54±0.56 |
6.27—8.34 7.03±0.86 |
5.78—8.76 6.97±0.78 |
| 酸碱度pH | 7.64—8.62 8.03±0.34 |
7.69—8.36 7.98±0.26 |
7.84—8.58 8.12±0.29 |
7.68—8.42 8.06±0.27 |
| 亚硝酸氮NO2--N (mg/L) | 0.008—0.317 0.17±0.12 |
0.005—0.352 0.19±0.12 |
0.003—0.153 0.08±0.06 |
0.004—0.124 0.07±0.05 |
| 硝酸氮NO3--N (mg/L) | 0.98—1.78 1.28±0.33 |
1.06—1.67 1.32±0.26 |
0.78—1.44 1.14±0.25 |
0.89—1.38 1.16±0.19 |
| 氨氮TAN (mg/L) | 0.11—0.34 0.20±0.08 |
0.04—0.24 0.16±0.07 |
0.08—0.25 0.18±0.06 |
0.06—0.36 0.23±0.11 |
| 活性磷酸盐AP(mg/L) | 0.061—0.175 0.10±0.04 |
0.078—0.169 0.12±0.03 |
0.072—0.154 0.11±0.03 |
0.067—0.182 0.12±0.05 |
| 总氮TN (mg/L) | 1.21—2.46 1.75±0.44 |
1.19—2.57 2.00±0.54 |
1.06—1.79 1.47±0.25 |
1.24—1.86 1.58±0.25 |
| 总磷TP (mg/L) | 0.08—0.20 0.12±0.04 |
0.10—0.19 0.14±0.03 |
0.09—0.17 0.12±0.03 |
0.07—0.18 0.13±0.05 |
| 氮磷比N:P | 22.40—30.50 15.22±4.82 |
22.40—30.50 14.17±2.23 |
22.40—30.50 12.94±1.75 |
22.40—30.50 13.19±3.02 |
 |
| 图 1 各养殖池塘水质因子变化情况 Fig. 1 Ranges of the water quality factors in the ponds 注: a.氨氮(TAN); b.硝酸氮(NO3--N); c.亚硝酸氮(NO2--N); d.活性磷酸盐(AP); e.总氮(TN); f.总磷(TP) |
(TC)。养殖前中期, 对虾养殖塘中活性磷酸盐(AP)含量高于缢蛏养殖塘, 而养殖后期则相反。
2.2 浮游植物种类及组成实验池塘中共鉴定出浮游植物6门、101种(包含9个未定种), 各门具体种类数量分布见图 2, 浮游植物名录见表 2。其中硅藻门(Bacillariophyta)共61种, 占所有种类数的60.4%, 所占比例最高; 其次是甲藻门(Pyrrophyta)、绿藻门(Chlorophyta)、裸藻门(Euglenophyta)、蓝藻门(Cyanophyaita)和隐藻门(Xanthophyta)。
 |
| 图 2 浮游植物各门种类数量 Fig. 2 Species of phytoplankton in each category |
| 种类 | 种类 | ||
| 绿藻门 Chlorophyta |
小球藻Chlorella vulgaris | 巨圆筛藻Coscinodiscus gigas | |
| 蛋白核小球藻Chlorella pyrenoidesa | 小角毛藻Chaetoceros minutissimu | ||
| 眼状微绿球藻Nannochloris oculata | 尖布纹藻Gyrosigma acuminatum | ||
| 波吉卵囊藻Oocystis borgei | 条纹小环藻Cyclotella triata | ||
| 盐生杜氏藻Dunaliella salina | 扭曲小环藻Cyclotella comta | ||
| 空球藻Eudorina elegans | 具槽帕拉藻Paralia sulcata | ||
| 空星藻Coelastrum sphaericum | 菱形海线藻Thalassiosira nitzschioides | ||
| 温声隐杆藻Aphanothece caldariorum | 尖刺伪菱形藻Pseudo pungens | ||
| 银灰平裂藻Merismopedia glauca | 柔弱几内亚藻Guinardia delicatula | ||
| 线形硬毛藻Chaetomorpha linum | 柔弱伪菱形藻Pseudo delicatissim | ||
| 小形卵囊藻Oocystis parva | 短楔形藻Licmophora abbreviate | ||
| 硅藻门 Bacillarionphyta |
直链藻Melosira sp. | 脆指管藻Dactyliosolen fragilissimus | |
| 圆筛藻Coscinodiscus sp. | 线性舟形藻Navicula graciloid | ||
| 辐射圆筛藻Coscinodiscus radiatus | 狭轴舟形藻Navicula verecunda | ||
| 线形圆筛藻Coscinodiscus lineatus | 披针形舟形藻Navicula lanceolat | ||
| 中心圆筛藻Coscinodiscus centrali | 小头桥湾藻Cymbella microcepha | ||
| 星脐圆筛藻Coscinodiscus asteromphalus | 假弯角毛藻Chaetoceros pseudocurvisetus | ||
| 虹彩圆筛藻Coscinodiscu soculusiridis | 短纹异极藻Gomphonema abbreviatum | ||
| 细柱藻Leptocylindrus minimus | 杂菱形藻Nitzschia hybrida | ||
| 洛氏角毛藻Chaetoceros lorenzianus | 海洋菱形藻Nitzschia marina | ||
| 牟氏角毛藻Chaetoceros muelleri | 叉翼羽纹藻Pinnularia stauroptera | ||
| 旋链角毛藻Chaetoceros curvisetus | 甲藻门 Pyrrophyta |
海洋原甲藻Prorocentrum micans | |
| 布氏双尾藻Ditylum brightwellii | 微小原甲藻Prorocentrum minimum | ||
| 舟形藻Navicula sp. | 裸甲藻Gymnodinium sp. | ||
| 新月菱形藻Nitzschia closteriu | 多甲藻Peridinium sp. | ||
| 针状菱形藻Nitzschia acicularis | 三角角藻Ceratium tripos | ||
| 平滑舟形藻Navicula laevissim | 夜光藻Noctilucas cintillans | ||
| 短小舟形藻Navicula exigua | 叉角藻Ceratium fusus | ||
| 系带舟形藻Navicula cinct | 薄甲藻Glenodinium pulvisculus | ||
| 近缘桥弯藻Cymbella aequal | 具尾翅甲藻Dimophusis caudata | ||
| 海洋曲舟藻Pleurosigma pelagicum | 梭状角藻Ceratium fusiform | ||
| 美丽曲舟藻Pleurosigma formosum | 叉状角藻Ceratium forkshape | ||
| 相似曲舟藻Pleurosigma affine | 扁平原多甲藻Protoperidinium depressum | ||
| 波罗的海布纹藻Gyrosigma balticum | 裸藻门 Euglenophyta |
纤细裸藻Euglena vjasgsfajs | |
| 中华盒形藻Biddulphia sinensis | 绿裸藻Euglena virids | ||
| 太阳漂流藻Planktoniella sol | 扁裸藻Phacus sp. | ||
| 美丽星杆藻Asterionella formosa | 扁圆囊裸藻Trachelomonas curta | ||
| 刚毛根管藻Rhizosolenia acuminata | 膝曲裸藻Euglena geniculata | ||
| 诺氏海链藻Thalassiosira nordenskioldi | 密盘裸藻Euglena wangi | ||
| 细弱海链藻Thalassiosira subtilis | 矮小扁裸藻Phacus nannos | ||
| 中肋骨条藻Skeletonema costatum | 尾棘囊裸藻Trachelomonas armata | ||
| 小环藻Cyclotella sp. | 蓝藻门 Cyanophyt |
色球藻Chroococcus sp. | |
| 梅尼小环藻Cyclotella meneghiniana | 具缘微囊藻Microcystis marginata | ||
| 具星小环藻Cyclotella stelligera | 坚实微囊藻Microcystis firma | ||
| 密集海链藻Thalassiosira condensata | 不定微囊藻Microcystis incerta | ||
| 针杆藻Synedra sp. | 弱细颤藻Oscillatoria tenuis | ||
| 简单舟形藻Navicula simple | 威利颤藻Oscillatoria willei | ||
| 库津小环藻Cyclotella kuetzingiana | 窝形席藻Phormidium foveolarum | ||
| 具翼漂流藻Planktoniellablanda | 隐藻门 Cryptophyta |
卵形隐藻Cryptomonas ovata | |
| 膜质半管藻Hemiaulus membranaceus | 长形蓝隐藻Chroomonas oblonga | ||
| 念珠直链藻Melosira moniliformis |
四个试验塘中浮游植物丰度与生物量变化见图 3。
 |
| 图 3 浮游植物丰度、生物量变化 Fig. 3 Phytoplankton biomass in different sampling periods |
养虾塘中浮游植物丰度介于6.8×105—6.1×107cell/L之间, 生物量为6.63—65.72mg/L; 养蛏塘中浮游植物丰度介于2.2×106—2.5×108cell/L之间, 生物量为2.04—17.58mg/L。在6—8月间, 对虾浮游植物生物量随着养殖时间的推移不断升高, 与浮游植物密度变化规律基本相同, 浮游植物生物量在8月出现一个峰值, 随后气温下降, 进入缢蛏生长高峰期, 浮游植物被缢蛏大量摄食, 两个养殖塘之间水交换频繁, 浮游植物密度及生物量下降。由于缢蛏的滤食作用, 缢蛏养殖塘的浮游植物丰度及生物量均显著低于对虾养殖塘。
2.4 浮游植物生物多样性、均匀度变化四个试验塘中浮游植物生物多样性、均匀度变化见图 4。虾蛏循环养殖池塘Shannon-Wiener多样性指数介于1.34—2.56之间, 平均值为1.91, 最高值的出现在8月17日的TX-2池塘。四个池塘的多样性指数均表现为养殖初期最低, 表明养殖初期浮游植物种类较少, 群落的复杂程度小, 随着养殖的进行, 种类开始逐渐增多, 群落的复杂程度逐渐增大, 随后趋于平稳。
 |
| 图 4 生态循环养殖池塘中浮游植物多样性及均匀度指数变化 Fig. 4 The change of phytoplankton diversity and evenness index in the ecological cycle culture pond |
虾蛏循环养殖池塘均匀度指数范围为0.43—0.72, 平均值为0.60。总体来看, 凡纳滨对虾养殖塘TX-1、TX-2变化相对比较平稳, 缢蛏养殖塘TC-1、TC-2波动较大, 说明对虾养殖塘浮游植物群落物种分布的均匀度较好。
2.5 浮游植物优势种及优势度四个试验塘中浮游植物优势种见表 3。在采集获得的101种浮游植物中14种为优势种(含1未定种), 包括新月菱形藻(N. closteriu)、梅尼小环藻(C. meneghiniana)、牟氏角毛藻(C. muelleri)、中肋骨条藻(S. costatum)、桥弯藻(C. aequal)、丹麦细柱藻(L. minimus)、直链藻(Melosira sp.)共7种硅藻门种类; 甲藻门海洋原甲藻(P. micans)、夜光藻(Noctilucas cintillans)2种; 2种蓝藻门种类, 为不定微囊藻(M. incerta)、色球藻(Chroococcus sp.); 2种绿藻门种类, 为小球藻(C. vulgaris)、波吉卵囊藻(O. borgei); 1种裸藻门种类, 绿裸藻(E. virids)。养殖初期硅藻门和绿藻门为主要优势种, 优势度较高, 养殖中期, 气温较高, 对虾生长速度加快, 投喂量逐渐增多, 残饵等有机质逐渐累积, 甲藻门和绿藻门逐渐占据优势, 之后气温开始下降, 缢蛏摄食能力增强, 两个养殖塘之间水交换频繁, 硅藻门又重新成为主要优势种。
| 优势种 | 代码 |
| 新月菱形藻N. closteriu | P1 |
| 牟氏角毛藻 C. muelleri |
P2 |
| 梅尼小环藻 C. meneghiniana |
P3 |
| 小球藻C. vulgaris | P4 |
| 中肋骨条藻 S. costatum |
P5 |
| 海洋原甲藻P. micans | P6 |
| 桥弯藻C. aequal | P7 |
| 色球藻 Chroococcus sp. |
P8 |
| 绿裸藻 Euglena virids |
P9 |
| 夜光藻 Noctilucas cintillans |
P10 |
| 直链藻Melosira sp. | P11 |
| 不定微囊藻 M. incerta |
P12 |
| 波吉卵囊藻 O. borgei |
P13 |
| 丹麦细柱藻 L. minimus |
P14 |
在虾蛏生态循环养殖模式下, 缢蛏养殖塘与对虾养殖塘不同季节中浮游植物优势种差别较小, 但优势度(Y)存在显著差异(见表 4)。对虾养殖塘中的角毛藻、小球藻等粒径较小的藻类在流入缢蛏养殖塘后优势度出现显著下降; 而菱形藻、桥弯藻、丹麦细柱藻、直链藻等粒径较大的藻类优势度出现显著上升。活动能力较强和在水表层集群的微藻, 如绿裸藻等, 在流入缢蛏养殖塘后优势度也呈上升状态。
| 日期 (月.日) |
浮游植物优势种 | 对虾塘优 势度 |
缢蛏塘优 势度 |
| 06.15 | 新月菱形藻N. closteriu | 0.12 | 0.45↑ |
| 牟氏角毛藻C. muelleri | 0.36 | 0.09↓ | |
| 梅尼小环藻C. meneghiniana | 0.09 | 0.08 | |
| 小球藻C. vulgaris | 0.12 | 0.05↓ | |
| 07.16 | 牟氏角毛藻C. muelleri | 0.56 | 0.12↓ |
| 中肋骨条藻S. costatum | 0.08 | 0.24↑ | |
| 梅尼小环藻C. meneghiniana | 0.11 | 0.13 | |
| 海洋原甲藻P. micans | 0.02 | 0.04 | |
| 桥弯藻C. aequal | 0.04 | 0.11↑ | |
| 08.17 | 丹麦细柱藻L. minimus | 0.15 | 0.24↑ |
| 海洋原甲藻P. micans | 0.34 | 0.11↓ | |
| 绿裸藻Euglena virids | 0.05 | 0.35↑ | |
| 牟氏角毛藻C. muelleri | 0.11 | 0.04↓ | |
| 夜光藻Noctilucas cintillans | 0.03 | ||
| 09.13 | 丹麦细柱藻L. minimus | 0.34 | 0.54↑ |
| 直链藻Melosira sp. | 0.05 | 0.12↑ | |
| 海洋原甲藻P. micans | 0.04 | 0.08 | |
| 色球藻Chroococcus sp. | 0.15 | 0.01↓ | |
| 10.10 | 直链藻Melosira sp. | 0.19 | 0.76↑ |
| 小球藻C. vulgaris | 0.38 | 0.05↓ | |
| 波吉卵囊藻O. borgei | 0.06 | 0.08 | |
| 丹麦细柱藻L. minimus | 0.03 | 0.01 | |
| 不定微囊藻M. incerta | 0.05 | 0.02 | |
| 11.02 | 直链藻Melosira sp. | 0.37 | 0.36 |
| 桥弯藻C. aequal | 0.06 | 0.24↑ | |
| 小球藻C. vulgaris | 0.16 | 0.06↓ | |
| 注: ↑表示优势度显著上升, ↓表示优势度显著下降 | |||
首先对14种浮游微藻优势种进行DCA分析, DCA分析结果显示梯度轴长度(lengths of gradient)均小于3, 因此选择线性排序方法CCA(典范对应分析), CCA是分析浮游植物与环境因子关系的有效手段, 通过对环境变量和浮游植物排序作图, 能直观反映浮游植物与环境因子之间的关系。排序轴1、2的特征值分别为0.7818和0.6831, 浮游植物种类与水质因子的相关性分别为0.892和0.789;水质因子与浮游植物的前两轴相关性系数较高, 其线性结合度很好地反映了物种与水质因子之间的关系。
浮游植物优势种与水质因子的CCA排序图(图 5)可见: pH (0.8314)、S (0.7932)、TN (–0.9126)、NO3--N (–0.7623)、NO2--N (–0.7541)主要贡献于排序轴1, 排序轴1与TN的负相关性最大(–0.9126), 与pH的正相关性最大(0.8314)。AP (0.7736)、T (0.7146)、AP (0.7521)、TAN (0.3511)、DO (–0.5789)主要贡献于排序轴2。排序轴2与DO的负相关性最大(–0.5789), 与AP的正相关性最大(0.7736)。
 |
| 图 5 生态循环养殖池塘中浮游植物与环境因子的CCA排序图 Fig. 5 CCA sequencing map of phytoplankton and environmental factors in the captive ponds |
P1(新月菱形藻)、P2(牟氏角毛藻)、P3(梅尼小环藻)、P5(中肋骨条藻)、P7(桥弯藻)等硅藻门种类分布于第1和第4象限, 表明它们与温度T、盐度S和pH等理化因子呈正相关性。P6(海洋原甲藻)、P8(色球藻)、P9(绿裸藻)、P10(夜光藻)、P12(不定微囊藻)分布于第2象限, 与温度和TP、AP呈正相关性。P4(小球藻)、P11(直链藻)、P13(波吉卵囊藻)、P14(丹麦细柱藻)位于第3象限, 与NO2--N、NO3--N、TN呈显著负相关。
3 讨论 3.1 虾蛏循环养殖池塘中水质因子变化特征虾蛏生态循环养殖模式下, 对虾养殖塘的总磷(TP)、活性磷酸盐(AP)和硝酸盐氮(NO3--N)含量在养殖期间中呈现先上升后下降的趋势。这与许多已有的报道不一致, 关仁磊等(2015)研究表明虾池活性磷酸盐与硝酸盐氮含量在养殖前期保持稳定, 在中后期逐渐上升。Ren等(2008)研究表明总磷(TP)在养殖期间始终呈上升趋势。探究其原因, 首先TP、AP上升的原因是在7月、8月水温最高, 达到32.6℃, 对虾生长速度加快, 投喂量逐渐增多, 残饵等有机质逐渐累积; 其次7月、8月水温过高, 缢蛏生长速度减慢, 对藻类摄食减少, 导致两个养殖塘之间水交换减少; 此外可能因为水温逐渐升高, 促进对虾塘底泥中磷的释放, 姜敬龙等(2008)的研究表明, 底泥中磷释放速度随温度的升高而升高; 这些因素造成8月TP、AP浓度最高。后期下降的原因则是9月、10月份水温逐渐降低, 加之缢蛏摄食能力增强, 两个养殖塘之间水交换频繁, 故对虾养殖塘中的氮磷被藻类大量消耗, 并未出现养殖后期氮磷含量急剧升高现象, 始终保持在较低的范围内。
Jones(2001)的研究表明滤食性贝类在减少水体内悬浮的颗粒物、TP和TN的同时, 可能会增加水体中活性磷酸盐的含量。本研究中缢蛏养殖塘中的总氮(TN)含量始终低于对虾养殖塘, 缢蛏养殖塘中活性磷酸盐(AP)含量高于对虾养殖塘, 与Jones(2001)的研究结果相符。Redfield(1963)研究表明海水中磷元素为浮游植物生长主要限制因子, 故大多数生态系统中增加无机磷的含量将会促进浮游植物的生长。实践表明, 对虾收获后, 对虾养殖塘可以继续作为藻类塘使用, 缢蛏养殖塘中富含活性磷酸盐的水进入藻类塘, 用于促进浮游植物的生长。
3.2 虾蛏循环养殖池塘中浮游植物群落结构特征虾蛏循环养殖池塘在凡纳滨对虾养殖期间共鉴定出浮游植物6门101种(包含9个未定种), 从种的数量上来看, 硅藻门 > 甲藻门 > 绿藻门 > 裸藻门 > 蓝藻门 > 隐藻门。李景等(2015)研究温棚高产养虾池中浮游植物群落时绿藻多为常见种。李雪松等(2006)对福建凡纳滨对虾养殖池塘的研究结果显示大部分优势种为硅藻门种类。沈明明(2016)在对凡纳滨对虾工厂化养殖的研究中发现, 养殖前期以绿藻门、硅藻门和部分甲藻门种类为主要优势种, 中后期优势种主要是耐污喜氮的甲藻和绿藻门种类, 蓝藻门的微囊藻和颤藻也会在后期演替成为优势种。杨秀兰等(2002)在研究盐碱地封闭式对虾养殖中浮游生物时认为, 随着养殖时间的不断推移, 水体中剩余饲料及对虾粪便不断累积, 在微生物的转化作用下, 氮磷等营养元素含量迅速升高, 一些喜氮的蓝藻门和绿藻门种类大量繁殖。由此可见, 露天养虾池塘中硅藻更容易成为优势种, 而封闭式或温棚养虾池中则蓝绿藻更盛, 尤其是养虾后期表现突出。
本研究共鉴别出优势种14种, 其中硅藻门7种, 甲藻门2种, 蓝藻门2种, 绿藻门2种, 裸藻门1种。优势度最高的是硅藻门的丹麦细柱藻(0.39), 养殖初期主要优势种为硅藻门, 养殖中期, 甲藻门和绿藻门逐渐占据优势, 硅藻门又重新成为主要优势种。养殖后期优势种与前人研究结果均有所差异, 可能原因为本研究养殖模式为虾蛏生态循环养殖, 对虾养殖后期正值缢蛏生长高峰期, 摄食能力增强, 两个养殖塘之间水交换频繁, 水中氮磷等营养元素被浮游植物大量消耗, 水质保持较好, 故养殖后期主要优势种仍为硅藻门, 蓝藻门、甲藻门种类在后期演替成为优势种的现象并未出现。
表 4结果表明, 在虾蛏生态循环养殖模式下, 缢蛏养殖塘与对虾养殖塘中浮游植物优势种的优势度(Y)存在显著差异。对虾养殖塘中呈现显著优势的角毛藻、小球藻等粒径较小的藻类, 在流入缢蛏养殖塘被缢蛏滤食后出现优势度显著下降; 相反, 菱形藻、桥弯藻、丹麦细柱藻、直链藻等粒径较大的藻类优势度则出现上升, 究其原因应该是缢蛏的选择性摄食作用所致, 优先滤食粒径较小喜食的浮游植物。同时, 那些活动能力较强或在水面集群的微藻(如绿裸藻), 由于缢蛏在底部潜泥难以摄食, 在缢蛏养殖塘中优势度亦出现上升, 故缢蛏优先摄食粒径较小、且活动能力较弱的微藻。
Shannon-Wiener指数基于物种数量反映群落种类多样性, 常作为水质评价的指标, 一般H′值越大, 群落所含的信息量越大, 其数值越大说明水质越好, 受污染程度越低(孙濡泳, 2001)。均匀度指数J反映一个群落中全部物种个体数目的分配状况, 变化平稳表示群落物种分布的均匀度较好(孙雪梅等, 2013)。养殖水体中浮游植物的多样性指数和均匀度指数与浮游藻类种类成正比, 多样性指数和均匀度指数越小, 浮游藻类种类越单一, 影响养殖水环境的稳定, 导致养殖水体对外来变化的缓冲能力相对较弱。张瑜斌等(2009)研究表明多样性指数越大, 物种群落结构越稳定, 对稳定养殖水环境的生态系统具有重要意义。本实验养殖池塘的多样性指数和均匀度指数分别为1.34—2.56、0.43—0.72, 浮游植物生物多样性较好, 群落复杂程度较高, 表明该生态循环养殖系统稳定性较好。
3.3 浮游植物群落与水质因子相关性Piehler等(2004)对凡纳滨对虾养殖池塘的研究显示, 盐度是影响对虾养殖塘浮游植物群落演替的主要因素, 盐度较低时, 水中微藻优势种多以蓝藻种类为主, 而当盐度达到15—30时, 硅藻门的一些种类占据优势。本研究中盐度变化范围为18—25, 故硅藻门在鉴定出的种类中所占比例最高, 且在养殖过程中始终为优势种。
浮游植物的丰度变化、群落演替均与水中氮磷元素含量密切相关, 氮磷等营养盐往往是限制对虾养殖池初级生产力的主要因素, 硅藻门种类对无机氮和无机磷的最低浓度要求分别为0.079mg/L和0.018mg/L, 低于此浓度, 硅藻繁殖速度将受到明显抑制。米振琴等(1999)在对凡纳滨对虾半集约化养殖塘的研究中发现, 养殖池塘水体中总氮(TN)、总磷(TP)含量会影响浮游植物丰度及生物量; 彭聪聪等(2012)对斑节对虾精养池中浮游植物与水质因子相关性的研究表明, 影响浮游植物丰度的水质因子依次为活性磷酸盐 > 氨氮 > 亚硝酸盐氮 > 硝酸盐氮 > 温度 > 溶解氧。此外, 李卓佳等(2008)的研究结果表明绿藻门和蓝藻门种类对于氮磷比的需求差异较大, 大部分绿藻的繁殖依赖高氮磷比, 而氮磷比较低则有利于蓝藻繁殖。在本研究中, 通过对生态循环养殖池浮游植物优势种与水质因子进行CCA分析后发现, 硅藻门的新月菱形藻、牟氏角毛藻、梅尼小环藻、中肋骨条藻、桥弯藻与温度T、盐度S显著正相关; 甲藻门的海洋原甲藻、夜光藻, 蓝藻门的色球藻、不定微囊藻, 裸藻门的绿裸藻与温度和TP、AP正相关; 绿藻门的小球藻、波吉卵囊藻, 硅藻门的直链藻、丹麦细柱藻与NO2--N、NO3--N、TN呈显著负相关。
浮游植物的群落结构变化是水质因子在不同时间和空间作用的结果(王旭娜等, 2018), CCA分析结果表明硅藻门种类主要受盐度、温度、pH的影响, 绿藻门种类主要受氮含量(总氮、氨氮、亚硝酸盐氮)的影响, 甲藻门、蓝藻门种类主要受磷含量(总磷、活性磷)与温度的影响。
4 结论虾蛏生态循环养殖池塘中水体质量能始终维持在较高水平, 在养殖中后期变化较平稳, 氨氮、亚硝酸盐水质因子始终处于较低水平。这种虾蛏生态循环养殖模式有利于改善浮游植物群落结构, 养殖过程中硅藻门种类为主要优势种, 且抑制养殖后期有害蓝藻、甲藻种类的大量繁殖。CCA分析后得到硅藻门种群变化主要与温度、盐度、pH密切相关(P < 0.05);绿藻门种群优势受氮含量(总氮、亚硝酸盐氮、氨氮)的影响较大; 而甲藻门、蓝藻门种类则与磷含量(总磷、活性磷)与温度相关。
王旭娜, 江敏, 钟锐, 等. 2018. 凡纳滨对虾养殖池塘中浮游植物群落结构与水质因子的关系. 水产学报, 42(11): 1787-1803 |
刘招坤. 2014. 独立供饵式缢蛏池塘蓄水养殖系统设计与试验研究. 渔业现代化, 41(6): 18-21 DOI:10.3969/j.issn.1007-9580.2014.06.004 |
关仁磊, 向斌, 陈兆明, 等. 2015. 对虾地膜精养池浮游生物的时空差异. 热带生物学报, 6(1): 30-40 DOI:10.3969/j.issn.1674-7054.2015.01.005 |
米振琴, 谢骏, 潘德博, 等. 1999. 精养虾池浮游植物、理化因子与虾病的关系. 上海水产大学学报, 8(4): 304-308 |
孙雪梅, 夏斌, 过锋, 等. 2013. 青岛崂山近岸海域浮游植物群落结构及其与环境因子的关系. 渔业科学进展, 34(3): 46-53 DOI:10.3969/j.issn.1000-7075.2013.03.006 |
孙濡泳. 2001. 动物生态学原理. 3版. 北京: 北京师范大学出版社
|
李景, 陈昌 福.. 2015. 温棚高产养虾池中浮游植物群落与水化学因子特征. 中国水产, (7): 72-76 DOI:10.3969/j.issn.1002-6681.2015.07.034 |
李卓佳, 梁伟峰, 陈素文, 等. 2008. 虾池常见微藻的光照强度、温度和盐度适应性. 生态学杂志, 27(3): 397-400 |
李雪松, 梁君荣, 陈长平, 等. 2006. 泉州湾虾池浮游植物种类多样性研究. 厦门大学学报(自然科学版), 45(S1): 234-239 |
杨秀兰, 王爱敏, 薄学锋, 等. 2002. 浮游生物在盐碱地封闭式对虾养殖中的生态作用. 齐鲁渔业, (10): 5-8 |
沈国英, 施并 章.. 2002. 海洋生态学. 2版. 北京: 科学出版社
|
沈明明..2016.影响对虾工厂化养殖水体微藻群落演替的主要因子及其与疾病发生的关系.上海: 上海海洋大学硕士学位论文
|
张继民, 刘霜, 张琦, 等. 2010. 黄河口附近海域浮游植物种群变化. 海洋环境科学, 29(6): 834-837 DOI:10.3969/j.issn.1007-6336.2010.06.013 |
张瑜斌, 龚玉艳, 陈长平, 等. 2009. 高位虾池养殖过程浮游植物群落的演替. 生态学杂志, 28(12): 2532-2540 |
周井 娟.. 2016. 中国对虾养殖业发展轨迹及技术变迁. 中国农学通报, 32(8): 22-29 |
胡韧, 林秋奇, 段舜山, 等. 2002. 热带亚热带水库浮游植物叶绿素a与磷分布的特征. 生态科学, 21(4): 310-315 |
姜敬龙, 吴云 海.. 2008. 底泥磷释放的影响因素. 环境科学与管理, 33(6): 47-50 DOI:10.3969/j.issn.1673-1212.2008.06.014 |
彭聪聪, 李卓佳, 曹煜成, 等. 2012. 斑节对虾滩涂养殖池塘浮游微藻群落演变特征. 安全与环境学报, 12(5): 95-101 DOI:10.3969/j.issn.1009-6094.2012.05.021 |
Davis D A, Arnold C R., 1998. The design, management and production of a recirculating raceway system for the production of marine shrimp. Aquacultural Engineering, 17(3): 193-211 DOI:10.1016/S0144-8609(98)00015-6 |
Jones F H., 2001. Teeth and bones:applications of surface science to dental materials and related biomaterials. Surface Science Reports, 42(3-5): 75-205 DOI:10.1016/S0167-5729(00)00011-X |
Piehler M F, Twomey L J, Hall N S et al, 2004. Impacts of inorganic nutrient enrichment on phytoplankton community structure and function in Pamlico Sound, NC, USA. Estuarine, Coastal and Shelf Science, 61(2): 197-209 DOI:10.1016/j.ecss.2004.05.001 |
Redfield D., 1963. Effect of defect fields on the optical absorption edge. Physical Review, 130(3): 916-918 DOI:10.1103/PhysRev.130.916 |
Ren J S, Schiel D R., 2008. A dynamic energy budget model:parameterisation and application to the Pacific oyster Crassostrea gigas in New Zealand waters. Journal of Experimental Marine Biology and Ecology, 361(1): 42-48 DOI:10.1016/j.jembe.2008.04.012 |
Rodríguez S R, Regalado E M, Pérez J A C et al, 2007. Comparison of some reproductive characteristics of farmed and wild white shrimp males Litopenaeus vannamei (Decapoda:Penaeidae). Revista de Biologia Tropical, 55(1): 199-206 |
Rott E, Cantonati M, Füreder L et al, 2006. Benthic algae in high altitude streams of the Alps-a neglected component of the aquatic biota. Hydrobiologia, 562(1): 195-216 DOI:10.1007/s10750-005-1811-z |
Sidik M J, Rashed-Un-Nabi M, Hoque M A., 2008. Distribution of phytoplankton community in relation to environmental parameters in cage culture area of Sepanggar Bay, Sabah, Malaysia. Estuarine, Coastal and Shelf Science, 80(2): 251-260 DOI:10.1016/j.ecss.2008.08.004 |
Xu L P, Xiong B X, Pan Y et al, 2010. Relationship between concentrations of odorous compounds and biomass of phytoplankton and actinomycetes in freshwater ponds of Beijing, China. Aquaculture International, 18(3): 245-254 DOI:10.1007/s10499-009-9240-z |