 2020, Vol. 51
2020, Vol. 51中国海洋湖沼学会主办。
文章信息
- 黄丁勇, 李元超, 王建佳, 郑新庆. 2020.
- HUANG Ding-Yong, LI Yuan-Chao, WANG Jian-Jia, ZHENG Xin-Qing. 2020.
- 亚龙湾西岸造礁石珊瑚种类多样性及恢复潜力分析
- ANALYSIS ON DIVERSITY AND POTENTIAL RECOVERY OF SCLERACTINIA CORALS IN THE WEST BANK OF YALONG BAY
- 海洋与湖沼, 51(3): 444-455
- Oceanologia et Limnologia Sinica, 51(3): 444-455.
- http://dx.doi.org/10.11693/hyhz20191100220
文章历史
-
收稿日期:2019-11-21
收修改稿日期:2020-02-01




2. 海南省海洋与渔业科学院 海口 570100;
3. 福建省海洋生态保护与修复重点实验室 厦门 361005
2. Hainan Academy of Ocean and Fisheries Sciences, Haikou 570100, China;
3. Fujian Provincial Key Laboratory of Marine Ecological Conservation and Restoration, Xiamen 361005, China
海南岛珊瑚礁分布广泛, 造礁石珊瑚种类丰富。三亚位于海南岛南端, 属海洋性热带季风气候, 适宜珊瑚生长(施祺等, 2009; 吴钟解等, 2012)。三亚珊瑚礁属于我国珊瑚礁分布的过渡区, 是最接近大洋典型分布区的珊瑚礁, 对华南沿海珊瑚礁群落的分布有着重要的意义(练健生等, 2010; 王道儒等, 2011; 吴钟解等, 2012; 周红英等, 2017)。
不过, 由于城市污水等原因, 三亚珊瑚礁总体状况不容乐观, 三亚造礁石珊瑚生物多样性热点地区已经从三亚湾的鹿回头转移到亚龙湾(练健生等, 2010)。亚龙湾位于三亚市南部, 向南面向广阔的南海, 其北岸为优质沙滩, 游客众多, 而西岸以石质基底为主, 人迹罕至, 仅太阳湾和白虎湾有游客潜水观光。亚龙湾内的野猪岛、东排岛、西排岛属于三亚珊瑚礁国家级自然保护区的核心区, 造礁石珊瑚和软珊瑚都较为丰富, 针对这三个岛的珊瑚礁已有较多的报道(练健生等, 2010; 李秀保, 2011; 吴钟解等, 2012; 吴川良等, 2019)。亚龙湾西岸位于西排岛的西侧, 距离西排岛仅2km, 推测应该也会有相对丰富的珊瑚资源, 但是目前尚未见到有关该海区的造礁石珊瑚的公开报道。
为了解亚龙湾西岸的造礁石珊瑚现状, 我们于2018年12月在亚龙湾西岸及北岸的西段布设了13个珊瑚礁调查站位, 系统调查了该区域造礁石珊瑚的多样性、分布、覆盖率、死亡率和补充量现状, 并结合文献资料比较了其中3个站位覆盖率和主要优势种的变化, 以及从补充量方面分析了亚龙湾西岸造礁石珊瑚的自然恢复潜力。本文填补了亚龙湾西岸造礁石珊瑚资料的空白, 为保护区的管理及周边涉海工程的环境影响评价等提供了支撑数据。
1 研究材料与方法 1.1 调查的时间和站位本次调查共设置13站位:亚龙湾北岸的西段5个站位(Y1-Y5)和亚龙湾西岸8个站位(Y6-Y10、A3、S52和S24), 如图 1所示。
 |
| 图 1 调查站位图 Fig. 1 Map of sampling site |
采用国际上通用的录像样带法(video transect method)(Page et al, 2001; 李秀保等, 2010), 在所有调查站位分别布设2条平行于海岸线(等深线)的50m长的断面, 断面分别设置于浅水区(1—3m)和深水区(4—10m)2个水深(断面设置与历史资料之保持一致以便比较, 详见后文的讨论)。沿着断面对珊瑚进行录像和拍照, 并采集少量样品, 用于珊瑚种类、覆盖率等分析。另外, 在每个50m断面中每隔2m拍摄一个50cm×50cm的珊瑚样方照片, 即一条断面有25张照片, 用于珊瑚补充量测算。
另对A3、S52和S24站位的深水区底层水(离底2m)进行八项海水水质检测, 检测内容包括溶解氧、化学需氧量(COD)、悬浮物、无机氮、活性磷酸盐、五日生化需氧量(BOD5)、叶绿素a浓度和浊度, 其中无机氮和活性磷酸盐的测定按GB/T 12763.4-2007进行, 叶绿素a浓度的测定按GB 17378.7-2007进行, 其余四项的测定按GB 17378.4-2007进行。
1.3 数据分析在电脑上判读野外调查获得的录像和照片。在分析珊瑚录像时, 将每个50m的样带等距离分为500个“点”, 统计活的造礁石珊瑚分布的“点”的数量并除以500, 即为该样带造礁石珊瑚覆盖率(黄晖等, 2011)。同理, 统计死亡造礁石珊瑚分布的“点”的数量, 除以500, 即为造礁石珊瑚死亡率。珊瑚死亡时间的判断依据珊瑚骨骼的颜色及腐蚀情况进行估计, 其中死亡30d以内的珊瑚骨骼为白色且完整清晰; 死亡1—2a的珊瑚骨骼结构轻微腐蚀但仍可分辨至属阶元; 而死亡2a以上的珊瑚骨骼结构已很难分辨, 或者其上附着的生物已经很难去除(黄晖等, 2012)。
珊瑚种类主要根据所拍的珊瑚录像和照片, 结合采集的样品进行鉴定。
通过珊瑚样方照片, 判读样方内是否有不大于2cm的造礁石珊瑚新补充个体。统计这些个体的数量并除以样方面积, 即为造礁石珊瑚补充量, 单位为ind./m2。
用Primer软件进行多样性指数分析和聚类分析。其中, 多样性指数(H′)、均匀度指数(J′)和优势度的计算方法分别参考Shannon(1948)、Pielou(1966)、徐兆礼和陈亚瞿(1989), 具体计算方法为:
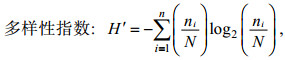 (1)
(1)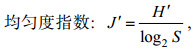 (2)
(2)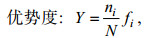 (3)
(3)其中, N为造礁石珊瑚总覆盖率, ni为第i种的覆盖率, S为种类数, fi为第i种在各站位的出现频率。优势种的确定由优势度决定(徐兆礼等, 1989)。
2 结果 2.1 水质亚龙湾西岸3个水质检测站位的结果显示, 亚龙湾西岸海水水质整体良好, 仅S24站位的溶解氧含量略微超第一类海水水质标准(5mg/L<5.99mg/L<6mg/L)外, 其余各项均符合第一类海水水质标准, 浊度、叶绿素a浓度和悬浮物含量也未见明显异常(表 1)。
| 站号 | 溶解氧(mg/L) | COD (mg/L) | 悬浮物(mg/L) | 无机氮(mg/L) | 活性磷酸盐(mg/L) | 浊度(NTU) | BOD5 (mg/L) | 叶绿素a (μg/L) |
| A3 | 6.63 | 0.77 | 11.0 | 0.037 | 0.005 | 1.1 | 0.52 | 2.35 |
| S52 | 6.23 | 0.66 | 24.9 | 0.010 | 0.003 | 0.8 | 0.67 | 2.26 |
| S24 | 5.99 | 0.93 | 20.5 | 0.026 | 0.004 | 3.7 | 0.56 | 2.48 |
| 平均值 | 6.28 | 0.79 | 18.8 | 0.024 | 0.004 | 1.9 | 0.58 | 2.36 |
| 第一类海水水质标准 | >6 | ≤2 | - | 0.20 | ≤0.015 | - | ≤1 | - |
| 注: -表示无数据; COD表示化学需氧量; BOD5:表示五日生化需氧量 | ||||||||
13个珊瑚调查站位中, 亚龙湾北岸的5个站位(Y1到Y5站位)全为沙质底质, 均未发现造礁石珊瑚及其他生物, 亚龙湾西岸8站位共发现造礁石珊瑚12科27属55种和11个未定种(表 2)。蜂巢珊瑚科(Faviidae)的种类最多, 包括9属25种, 其中扁脑珊瑚属(Platygyra)、角蜂巢珊瑚属(Favites)和蜂巢珊瑚属(Favia)三个属的种类最多, 分别有6种、5种和5种, 其余属分别只有1—3种不等。其次为鹿角珊瑚科(Acroporidae), 包括3属17种, 鹿角珊瑚属(Acropra)和蔷薇珊瑚属(Montipora)的种类最多, 皆分别有8种, 也是此次调查中含有最多种类的属。
| 科 | 属 | 序号 | 中文学名 | 拉丁学名 | 站位 | |||||||
| Y6 | Y10 | Y7 | A3 | Y8 | Y9 | S52 | S24 | |||||
| 鹿角珊瑚科 Family Acroporidae |
鹿角珊瑚属 Acropora |
1 | 花柄鹿角珊瑚 | Acropora anthocercis | + | |||||||
| 2 | 两叉鹿角珊瑚 | Acropora divaricata | + | + | ||||||||
| 3 | 花鹿角珊瑚 | Acropora florida | + | |||||||||
| 4 | 芽枝鹿角珊瑚 | Acropra gemmifera | + | |||||||||
| 5 | 风信子鹿角珊瑚 | Acropora hyacinthus | + | + | + | |||||||
| 6 | 壮实鹿角珊瑚 | Acropora robusta | + | |||||||||
| 7 | 强壮鹿角珊瑚 | Acropora valida | + | |||||||||
| 8 | 鹿角珊瑚 | Acropora sp. |
+ | + | + | |||||||
| 蔷薇珊瑚属 Montipora |
9 | 仙掌蔷薇珊瑚 | Montipora cactus | + | + | |||||||
| 10 | 指状蔷薇珊瑚 | Montipora digitata | + | + | + | + | + | |||||
| 11 | 叶状蔷薇珊瑚 | Montipora foliosa | + | |||||||||
| 12 | 变形蔷薇珊瑚 | Montipora informis | + | + | + | + | + | |||||
| 13 | 截顶蔷薇珊瑚 | Montipora truncata | + | + | + | + | ||||||
| 14 | 膨胀蔷薇珊瑚 | Montipora turgescens | + | + | + | |||||||
| 15 | 龟岛蔷薇珊瑚 | Montippora turtlensis | + | + | ||||||||
| 16 | 蔷薇珊瑚 | Montipora sp. |
+ | + | + | + | + | + | ||||
| 星孔珊瑚属 Astreopora |
17 | 多星孔珊瑚 | Astreopora myriophthalma | + | ||||||||
| 菌珊瑚科 Family Agariciidae |
牡丹珊瑚属 Pavona |
18 | 十字牡丹珊瑚 | Pavona decussata | + | + | + | + | + | |||
| 19 | 脉结牡丹珊瑚 | Pavona venosa | + | |||||||||
| 丁香珊瑚科 Family Caryophylliidae |
真叶珊瑚属 Euphyllia |
20 | 肾形真叶珊瑚 | Euphyllia ancora | + | |||||||
| 木珊瑚科 Family Dendrophylliidae |
陀螺珊瑚属 Turbinaria |
21 | 皱折陀螺珊瑚 | Turbinaria mesenterina | + | + | + | + | ||||
| 22 | 陀螺珊瑚 | Turbinaria sp. |
+ | |||||||||
| 蜂巢珊瑚科 Family Faviidae |
扁脑珊瑚属 Platygyra |
23 | 肉质扁脑珊瑚 | Platygyra carnosus | + | |||||||
| 24 | 精巧扁脑珊瑚 | Platygyra daedalea | + | + | + | + | + | + | ||||
| 25 | 片扁脑珊瑚 | Platygyra lamellina | + | |||||||||
| 26 | 小扁脑珊瑚 | Platygyra pipi | + | |||||||||
| 27 | 中华扁脑珊瑚 | Platygyra sinensis | + | + | + | + | ||||||
| 28 | 扁脑珊瑚 | Platygyra sp. |
+ | + | + | + | ||||||
| 肠珊瑚属 Leptoria |
29 | 弗利吉亚肠珊瑚 | Leptoria phrygia | + | + | + | ||||||
| 刺星珊瑚属 Cyphastrea |
30 | 锯齿刺星珊瑚 | Cyphastrea serailia | + | + | + | + | + | + | + | ||
| 31 | 刺星珊瑚 | Cyphastrea sp. |
+ | |||||||||
| 蜂巢珊瑚属 Favia |
32 | 黄癣蜂巢珊瑚 | Favia favus | + | + | + | + | + | ||||
| 33 | 圆形蜂巢珊瑚 | Favia rotundata | + | |||||||||
| 34 | 标准蜂巢珊瑚 | Favia speciosa | + | + | + | + | ||||||
| 35 | 圆突蜂巢珊瑚 | Favia veroni | + | + | + | + | + | + | ||||
| 36 | 蜂巢珊瑚 | Favia sp. |
+ | + | + | + | + | + | + | |||
| 角蜂巢珊瑚属 Favites |
37 | 秘密角蜂巢珊瑚 | Favites abdita | + | + | + | + | + | + | |||
| 38 | 板叶角蜂巢珊瑚 | Favites complanata | + | + | + | + | + | + | ||||
| 39 | 海孔角蜂巢珊瑚 | Favites halicora | + | + | + | + | + | + | + | + | ||
| 40 | 五边角蜂巢珊瑚 | Favites pentagona | + | |||||||||
| 41 | 角蜂巢珊瑚 | Favites sp. |
+ | + | + | + | + | + | + | |||
| 菊花珊瑚属 Goniastrea |
42 | 粗糙菊花珊瑚 | Goniastrea aspera | + | ||||||||
| 43 | 梳状菊花珊瑚 | Goniastrea pectinata | + | |||||||||
| 44 | 菊花珊瑚 | Goniastrea sp. |
+ | + | ||||||||
| 双星珊瑚属 Diploastrea |
45 | 同双星珊瑚 | Diploastrea hlipopra | + | + | + | + | + | + | + | ||
| 同星珊瑚属 Plesiastrea |
46 | 多孔同星珊瑚 | Plesiastrea versipora | + | + | + | + | |||||
| 小星珊瑚属 Leptastrea |
47 | 小星珊瑚 | Leptastrea sp. |
+ | + | + | ||||||
| 裸肋珊瑚科 Family Merulinidae |
刺柄珊瑚属 Hydnophora |
48 | 腐蚀刺柄珊瑚 | Hydnophora exesa | + | + | + | |||||
| 裸肋珊瑚属 Merulina |
49 | 阔裸肋珊瑚 | Merulina ampliata | + | ||||||||
| 褶叶珊瑚科 Family Mussidae |
棘星珊瑚属 Acanthastrea |
50 | 棘星珊瑚 | Acanthastrea echinata | + | + | + | |||||
| 合叶珊瑚属 Symphyllia |
51 | 辐射合叶珊瑚 | Symphyllia radians | + | + | + | ||||||
| 52 | 华贵合叶珊瑚 | Symphyllia nobilis | + | + | ||||||||
| 叶状珊瑚属 Lobophyllia |
53 | 赫氏叶状珊瑚 | Lobophyllia hemprichii | + | + | |||||||
| 枇杷珊瑚科 FamiliyOculinidae |
盔形珊瑚属 Galaxea |
54 | 丛生盔形珊瑚 | Galaxea fascicularis | + | + | + | + | + | + | + | + |
| 梳状珊瑚科Family Pectiniidae | 刺叶珊瑚属 Echinophyllia |
55 | 粗糙刺叶珊瑚 | Echinophyllia aspera | + | |||||||
| 杯形珊瑚科 Family Pocilloporidae |
杯形珊瑚属 Pocillopora |
56 | 鹿角杯形珊瑚 | Pocillopora damicronis | + | + | + | + | + | + | ||
| 57 | 多曲杯形珊瑚 | Pocillopora meandrina | + | + | ||||||||
| 58 | 疣状杯形珊瑚 | Pocillopora verrucosa | + | + | + | + | + | + | + | |||
| 59 | 伍氏杯形珊瑚 | Pocillopora woodjonesi | + | + | + | |||||||
| 滨珊瑚科 Family Poritidae |
角孔珊瑚属 Goniopora |
60 | 角孔珊瑚 | Goniopora sp. |
+ | + | + | + | + | + | + | |
| 滨珊瑚属 Porites |
61 | 澄黄滨珊瑚 | Porites lutea | + | + | + | + | + | + | + | ||
| 62 | 细柱滨珊瑚 | Porites cylindrica | + | |||||||||
| 63 | 火焰滨珊瑚 | Porites rus | + | + | ||||||||
| 64 | 滨珊瑚 | Porites sp. |
+ | + | ||||||||
| 铁星珊瑚科 Family Siderastreidae |
筛珊瑚属 Coscinaraea |
65 | 柱形筛珊瑚 | Coscinaraea columna | + | |||||||
| 沙珊瑚属 Psammocora |
66 | 浅薄沙珊瑚 | Psammocora superficialis | + | ||||||||
亚龙湾西岸8个站位的造礁石珊瑚种类数分别为12—33种, 平均27种(图 2)。除S24站位仅有12种外, 其余站位皆有20种以上, Y6站位的种类数最多(33种)。
 |
| 图 2 亚龙湾西岸造礁石珊瑚的种类数、覆盖率、死亡率和补充量 Fig. 2 Species number, coverage, mortality, and recruitment of scleractinian corals in the west bank of Yalong Bay |
亚龙湾西岸8个造礁石珊瑚平均覆盖率为12.9%, 浅水区平均14.5%, 深水区11.4%(表 3)。Y10站位的造礁石珊瑚覆盖率最高, 为22.9%, 其浅水区覆盖率高达25.2%。S24站位的覆盖率很低, 仅为2.8%。
| 站位 | 覆盖率(%) | 种类数(种) | 优势种 | ||||||
| 浅水区 | 深水区 | 浅水区 | 深水区 | 浅水区 | 深水区 | 站位 | |||
| Y6 | 15.4 | 10.8 | 25 | 19 | 丛生盔形珊瑚 | 丛生盔形珊瑚 | 丛生盔形珊瑚 | ||
| Y10 | 25.2 | 20.6 | 27 | 13 | 丛生盔形珊瑚、精巧扁脑珊瑚、秘密角蜂巢珊瑚、风信子鹿角珊瑚 | 澄黄滨珊瑚、蔷薇珊瑚sp.、丛生盔形珊瑚 | 丛生盔形珊瑚、澄黄滨珊瑚、蔷薇珊瑚sp. | ||
| Y7 | 4.6 | 11.0 | 8 | 18 | 壮实鹿角珊瑚 | 丛生盔形珊瑚 | 丛生盔形珊瑚 | ||
| A3 | 14.6 | 11.2 | 21 | 12 | 丛生盔形珊瑚、膨胀蔷薇珊瑚 | 同双星珊瑚 | 同双星珊瑚、丛生盔形珊瑚、膨胀蔷薇珊瑚 | ||
| Y8 | 12.6 | 8.4 | 19 | 13 | 风信子鹿角珊瑚 | 同双星珊瑚 | 同双星珊瑚 | ||
| Y9 | 16.6 | 14.4 | 27 | 20 | 多孔同星珊瑚、澄黄滨珊瑚、丛生盔形珊瑚 | 同双星珊瑚 | 同双星珊瑚 | ||
| S52 | 22.8 | 13.2 | 24 | 12 | 十字牡丹珊瑚、澄黄滨珊瑚、锯齿刺星珊瑚 | 火焰滨珊瑚 | 火焰滨珊瑚、十字牡丹珊瑚、澄黄滨珊瑚 | ||
| S24 | 4.4 | 1.2 | 11 | 1 | 角孔珊瑚 | 同双星珊瑚 | 同双星珊瑚 | ||
优势度最高的种类如图 3所示。虽然各站位的优势种有所差异, 但就整个调查区而言, 丛生盔形珊瑚(Galaxea fascicularis)优势度最高, 为0.13, 在亚龙湾西岸8个站位中都有分布, 不过平均覆盖率只有1.7%(图 3, 图 4)。丛生盔形珊瑚主要分布在亚龙湾西岸的北段, 特别是青梅河口码头附近的Y10和Y6站位, 覆盖率分别为4.9%和3.1%, 而在亚龙湾西岸的南段覆盖率较低。其次为同双星珊瑚(Diploastrea hlipopra, 主要分布在A3、Y8和Y9站位)和澄黄滨珊瑚(Porites lutea, 主要分布在Y6、Y10和Y9站位), 优势度分别为0.08和0.07, 平均覆盖率分别为1.1%和1.1%。其余种类的平均覆盖率皆小于1%。
 |
| 图 3 亚龙湾西岸18种优势度最高的造礁石珊瑚的覆盖率 Fig. 3 Coverage of 18 dominant scleractinian corals in the west bank of Yalong Bay |
 |
| 图 4 亚龙湾西岸造礁石珊瑚三个优势种的覆盖率分布 Fig. 4 Distribution of the coverage of three dominant scleractinian corals in the west bank of Yalong Bay |
亚龙湾西岸8个站位皆有发现造礁石珊瑚的新生个体, 补充量为0.5—2.8ind./m2, 平均为1.6 ind./m2。整体而言, 与浅水区相比, 深水区的珊瑚补充量相对较高(表 4)。
| 站位 | 补充量(ind./m2) | 死亡率(%) | |||||
| 浅水区 | 深水区 | 平均值 | 浅水区 | 深水区 | 平均值 | ||
| Y6 | 3.0 | 1.6 | 2.3 | 36.2 | 74.8 | 55.5 | |
| Y10 | 0.8 | 0.8 | 0.8 | 44.4 | 47.2 | 45.8 | |
| Y7 | 0.6 | 4.8 | 2.7 | 2.4 | 56.0 | 29.2 | |
| A3 | 1.4 | 1.0 | 1.2 | 69.0 | 71.6 | 70.3 | |
| Y8 | 1.4 | 2.8 | 2.1 | 57.6 | 79.8 | 68.7 | |
| Y9 | 0.6 | 2.8 | 1.7 | 79.2 | 80.6 | 79.9 | |
| S52 | 1.8 | 1.8 | 1.8 | 59.6 | 66.0 | 62.8 | |
| S24 | 0.4 | 0.6 | 0.5 | 41.4 | 46.6 | 44.0 | |
| 平均值 | 1.3 | 2.0 | 1.6 | 48.7 | 65.3 | 57.0 | |
亚龙湾西岸8个站位的造礁石珊瑚死亡率为29.2%—79.9%, 平均为57.0%。除了Y7和Y10站位外, 所占比例均超过50%, 其中Y9站位高达79.9%, A3、Y8和S52站位死亡率也较高, 分别为70.3%、68.7%和62.8%, 可见死亡珊瑚形成的石质基底是该海区的主要底质类型(表 4, 图 5)。大部分站位两个水深的死亡率相近, 但深水区的造礁石珊瑚死亡率皆高于浅水区。
 |
| 图 5 亚龙湾西岸的底质类型 Fig. 5 Habit types in the west bank of Yalong Bay |
本次调查发现, 死亡的造礁石珊瑚几乎都是超过2a的, 仅A3和Y6站位有零星近期死亡的珊瑚(死亡30d以内), 而且近期死亡造礁石珊瑚的最高覆盖率也仅0.2%, 只出现在A3站位。
本次调查没有发现造礁石珊瑚的病害现象, 也未发现会啃食造礁石珊瑚的鹦嘴鱼。核果螺的数量也很少, 长棘海星(Acanthaster planci)仅在Y9和S52站位各发现一只, 目前皆不会对造礁石珊瑚造成明显危害。
2.6 多样性指数在种水平, 以覆盖率为数据矩阵对亚龙湾西岸8个站位进行多样性指数分析和聚类分析(图 6)。多样性指数分析结果显示各站位的均匀性指数(J′)和种类多样性指数(H′)相对都较高(梁文等, 2010; 孙有方等, 2018), 平均分别为0.85和3.99, 表明尽管有优势种的存在, 但大多数种类的覆盖率较为相近, 而且种类数量也较高。聚类分析发现站位间的相似性很低, 最高仅为49.57%(A3和Y8之间), 表明调查区造礁石珊瑚种类空间分布的异质性较高。
 |
| 图 6 多样性指数(a)和聚类分析(b) Fig. 6 Diversity index (a) and cluster analysis (b) |
造礁石珊瑚是珊瑚礁生态系统的最关键组分, 其生长和分布对珊瑚礁区其他生物的栖息和繁育有决定性影响, 所以其多样性状况在一定程度上可反映珊瑚礁区生物多样性的整体特征(Bellwood et al, 2001; 赵美霞等, 2006)。海南岛周边海域的造礁石珊瑚有34属110种和亚种(邹仁林, 2001), 三亚的造礁石珊瑚总记录数为86种(吴钟解等, 2013)。而有关亚龙湾的造礁石珊瑚种类数, 在2001年、2006年和2010—2011年开展的三次比较详细的调查中, 分别发现有46种(练健生等, 2010)、74种(练健生等, 2010)和50种(海南省海洋开发规划设计研究院等, 2011)。本次调查在亚龙湾西岸共发现了66种造礁石珊瑚(含11个未定种)而且平均每站有27种, 表明相对于海南岛其他周边海域而言, 亚龙湾西岸的造礁石珊瑚种类数相对较多, 多样性程度较高(种类多样性指数为3.99), 尽管覆盖率处于较低水平, 平均只有12.9%。该覆盖率水平与海南岛东海岸的铜鼓岭和三亚湾的东岛、西岛相当, 但低于蜈支洲、亚龙湾(岛礁)、鹿回头和红塘湾, 高于长圮港、龙湾、大东海和小东海(孙有方等, 2018; 海南省生态环境厅, 2019)。
调查区的造礁石珊瑚优势种以丛生盔形珊瑚、同双星珊瑚、澄黄滨珊瑚等团块状珊瑚为主, 还发现了一定数量的火焰滨珊瑚、十字牡丹珊瑚等皮壳状或叶状珊瑚, 分枝状的鹿角珊瑚虽然也有发现, 但都不是优势种, 仅在部分站位的浅水区相对较多出现, 如Y7、Y8和Y10站位。可见, 调查区的造礁石珊瑚群落整体处于群落演替初期, 并可能存在向演替中期发展的趋势(于登攀等, 1996; 周红英等, 2017)。与分枝状珊瑚相比, 团块状珊瑚属于宽生态位的种类, 能够适应较为恶劣的环境(Schlöder et al, 2004; 黄晖等, 2012; 周红英等, 2017), 而作为本次调查的优势种之一的同双星珊瑚可能是最耐受沉积物胁迫的种类(李秀保, 2011)。
本次调查的水质检测结果以及郑洋等(2015)、谢海群等(2016)的调查, 都显示亚龙湾水质均符合第一类海水水质标准(除了本次调查的溶解氧含量略微超第一类海水水质标准), 水质良好。不过, 悬浮物和沉积速率仍可能是亚龙湾珊瑚生长和繁育的一个限制因子(李秀保, 2011)。一般认为, 悬浮物含量超过10mg/L, 悬浮物的沉积速率超过10mg/(cm2·d)时, 会对造礁珊瑚造成不利影响, 长期超过此值则会造成严重的胁迫(Roger, 1990; Li et al, 2013a)。李秀宝在2007—2009年的调查中发现, 亚龙湾的沉积速率已达到会对珊瑚礁产生危害的阈值(李秀保, 2011)。特别是在亚龙湾的雨季, 悬浮物的沉积速率甚至会高达20mg/(cm2·d), 这会对一些环境敏感型造礁石珊瑚造成强烈的负面影响(Li et al, 2013b)。本次调查中, 亚龙湾西岸3个水质调查站位的悬浮物含量都超过了10mg/L, 其中S24和S52站位甚至超过了20mg/L, 这对于敏感型造礁石珊瑚而言, 调查区悬浮物含量偏高。
尽管自2014年后, 三亚加大了生态环境保护力度, 海水水质状况已有所好转(吴川良等, 2019)。但是, 正如前文所述, 虽然目前亚龙湾西岸的海水水质良好, 但对于分枝状鹿角珊瑚等环境敏感型的造礁石珊瑚而言, 区域悬浮物含量和沉积速率仍较高, 亚龙湾西岸可能仍不太适宜此类型造礁石珊瑚的生长, 或者说, 随着水体悬浮物含量的增加, 一些环境敏感型的造礁珊瑚已在亚龙湾西岸逐渐消亡从而引起造礁石珊瑚群落的演变, 导致调查区形成以团块状珊瑚为主要优势种的珊瑚群体。这与三亚其他海区(如西岛、鹿回头和大东海)团块状珊瑚优势度较高的情形一致(吴钟解等, 2012)。
3.2 亚龙湾西岸的造礁石珊瑚变化分析目前尚未见到有关亚龙湾西岸珊瑚礁的公开报道, 不过2010—2011年的三亚珊瑚礁国家级自然保护区综合科学考察指出, 三亚造礁石珊瑚平均覆盖率为17.2%(其中有4个站位位于亚龙湾西岸), 处于一个非常低的水平, 反映出三亚大部分区域的珊瑚礁处于严重退化状况(海南省海洋开发规划设计研究院等, 2011)。该调查报告显示, 以造礁石珊瑚覆盖率和种类数为指标, 保护区目前最好的珊瑚礁分布在坎秧湾-太阳湾分区以及亚龙湾野猪岛海域, 这包括了本次调查亚龙湾西岸的中部(A3站位)和南部(S52和S24站位)海域。为了增加对比的可信度, 本次调查中, 我们在2010—2011年的调查经纬度和断面水深重复了这3个站位的珊瑚礁调查。对比这3个站位的两次调查结果, 发现亚龙湾西岸造礁石珊瑚的覆盖率大幅下降, S52和S24两个站位优势种也发生较大变化, 表明在2011—2018年期间亚龙湾西岸珊瑚礁大幅衰退(表 5)。这可能与潜水旅游观光(海南省海洋开发规划设计研究院等, 2011)、恶劣天气(台风、强降雨)(古倩怡等, 2017)、珊瑚敌害生物和亚龙湾堤坝(吴川良等, 2019), 以及青梅河码头等沿岸涉海工程有关。
| 调查时间(年) | 站位 | 覆盖率(%) | 优势种 |
| 2010—2011 | A3 | 39.67 | 丛生盔形珊瑚和同双星珊瑚 |
| S52 | 61.17 | 丛生盔形珊瑚、鬃刺蔷薇珊瑚、火焰滨珊瑚 | |
| S24 | 13.83 | 丛生盔形珊瑚、交替扁脑珊瑚、叶形牡丹珊瑚、澄黄滨珊瑚 | |
| 2018 | A3 | 12.9 | 丛生盔形珊瑚和同双星珊瑚 |
| S52 | 18.0 | 火焰滨珊瑚、十字牡丹珊瑚、澄黄滨珊瑚 | |
| S24 | 2.8 | 同双星珊瑚 |
当环境变得适宜时, 珊瑚礁生态系统能展现出一定程度的自然恢复能力。珊瑚新个体的补充为珊瑚礁提供了主要的生态系统建设者, 是珊瑚恢复的主要机制, 在珊瑚礁恢复过程中具有非常重要的作用(Brandt et al, 2019; Speare et al, 2019; Yanovski et al, 2019)。对于确实能从重大干扰中恢复的珊瑚礁来说, 一个关键因素是具备由有性繁殖产生的幼虫所支持的高水平的珊瑚补充量(Price et al, 2019)。为了更直观地展示亚龙湾西岸造礁石珊瑚补充量的水平, 本文收集了已报道的三亚及周边海域的造礁石珊瑚补充量的相关信息(表 6)。尽管文献中对补充量的判断标准并不一致, 数据无法直接比较, 但从相对数量来看, 亚龙湾西岸与邻近的区域相比有着相对较高的造礁石珊瑚补充量水平。例如, 李元超等(2015)将直径10cm以内的珊瑚新个体的密度视为补充量, 认为与亚龙湾相邻的海棠湾后海海域2014年为4.5ind./m2; 而练建生等(2010)以5cm作为标准, 认为2006年亚龙湾(东排岛、西排岛和野猪岛)高达22ind./m2, 三亚湾和大、小东海为15ind./m2; 吴川良等(2019)同样以5cm作为标准, 认为亚龙湾(东排、西排)2012—2016年补充量只有0.397—1.4ind./m2; 但是海南岛其他海区大多不超过1ind./m2(吴钟解等, 2012, 2013; 李元超等, 2015), 如2018年海南岛东海岸平均为0.64ind./m2(未指明补充量的标准)(海南省生态环境厅, 2019)。本次调查以2cm作为标准, 显示亚龙湾西岸造礁石珊瑚平均补充量为1.6ind./m2, 部分站位达到2.7ind./m2(表 4), 在表 6的补充量数据中处于相对较高的水平, 表明亚龙湾西岸有相对丰富的珊瑚补充来源(吴川良等, 2019)。此外, 该海域适合珊瑚附着的硬基底所占比例很高(包括死亡珊瑚和礁石, 两者之和基本在60%以上)(图 4), 因此我们认为调查区的珊瑚礁有较好的自然恢复潜力。
| 调查时间(年) | 海区 | 判断标准(cm) | 补充量(ind./m2) | 数据来源 |
| 2014 | 海棠湾后海海域 | 10 | 4.5 | 李元超等, 2015 |
| 2006 | 亚龙湾(东排岛、西排岛和野猪岛) | 5 | 22 | 练建生等, 2010 |
| 2012—2016 | 三亚珊瑚保护区 | 5 | 0.397—1.4 | 吴川良等, 2019 |
| 2013—2014 | 海南大洲岛后港 | 5 | 0.02—0.20 | 古倩怡等, 2017 |
| 2007—2016 | 永兴岛和七连屿 | 2 | 0—0.97 | 李元超等, 2018 |
| 2018 | 亚龙湾西岸 | 2 | 0.5—2.7 | 本文 |
| 2018 | 海南岛东海岸 | 未指明 | 0.64 | 海南省生态环境厅, 2019 |
| 2012 | 七洲列岛 | 未指明 | 0.70—2.00 | 何晓娜等, 2018 |
| 2005—2009 | 西沙监控区 | 未指明 | 0.06—1.21 | 吴钟解等, 2011 |
| 2007 | 海南岛 | 未指明 | 0.1—0.86 | 吴钟解等, 2013 |
| 2006—2009 | 蜈支洲 | 未指明 | 0.39—1.87 | 吴钟解等, 2012 |
| 亚龙湾(东排、西排) | 未指明 | 0.42—0.65 | ||
| 大东海 | 未指明 | 0.1—3.9 | ||
| 鹿回头 | 未指明 | 0.65—1 | ||
| 西岛 | 未指明 | 0.4—0.62 |
本次调查共发现造礁石珊瑚12科27属55种和11个未定种, 亚龙湾西岸各站位造礁珊瑚介于12—33种之间, 平均27种。优势种以丛生盔形珊瑚、同双星珊瑚和澄黄滨珊瑚等环境耐受力较强的团块状珊瑚为主。造礁石珊瑚覆盖率为2.8%—22.9%, 平均12.9%。与2010—2011年相比, 亚龙湾西岸珊瑚礁大幅衰退, 悬浮物和沉积速率可能是该区域珊瑚礁退化的主要的原因。相对较高水平的造礁石珊瑚补充量及适宜珊瑚附着的硬基底比例, 预示着当海洋环境条件合适时, 调查区有自然恢复的可能。
从本次调查的造礁石珊瑚种类数、覆盖率和补充量数据来看, 亚龙湾西岸的造礁石珊瑚种类多样性较高且有自然恢复潜力, 而且种类分布异质性也较高, 具有一定的保护价值。为了保护亚龙湾西岸现有的珊瑚礁及加快其恢复速度, 建议加强人工干预, 比如严格管理海洋环境以减少悬浮物浓度和降低沉降速率, 同时控制潜水观光规模以减轻对珊瑚礁的破坏等。
致谢 感谢海南三亚海洋与渔业局在资料收集、野外调查协调等方面给予的众多帮助; 感谢海南亚龙湾海底世界旅游有限公司对此次野外调查所给予的配合和协助; 感谢李君民和张日可两位专业潜水教练在野外调查中的辛苦付出。
于登攀, 邹仁林. 1996. 鹿回头造礁石珊瑚群落多样性的现状及动态. 生态学报, 16(6): 559-564 |
王道儒, 王华接, 李元超, 等. 2011. 雷州半岛珊瑚幼虫补充来源初步研究. 热带海洋学报, 30(2): 26-32 |
古倩怡, 李洪武, 钱军, 等. 2017. 海南大洲岛后港造礁石珊瑚的种类组成与分布. 海南大学学报自然科学版, 35(4): 366-371 |
孙有方, 雷新明, 练健生, 等. 2018. 三亚珊瑚礁保护区珊瑚礁生态系统现状及其健康状况评价. 生物多样性, 26(3): 258-265 |
李元超, 杨毅, 郑新庆, 等. 2015. 海南三亚后海海域珊瑚礁生态系统的健康状况及其影响因素. 生态学杂志, 34(4): 1105-1112 |
李元超, 陈石泉, 郑新庆, 等. 2018. 永兴岛及七连屿造礁石珊瑚近10年变化分析. 海洋学报, 40(8): 97-109 |
李秀保, 2011.三亚造礁石珊瑚群落组成、时空分布及主要影响因子识别研究.广州: 中国科学院研究生院博士学位论文, 1-107
|
李秀保, 练健生, 黄晖, 等. 2010. 福建东山海域石珊瑚种类多样性及其空间分布. 台湾海峡, 29(1): 5-11 |
吴川良, 李长青, 张文勇, 等. 2019. 三亚国家级珊瑚礁自然保护区珊瑚礁资源的多样性. 热带生物学报, 10(1): 14-21 |
吴钟解, 王道儒, 叶翠信, 等. 2012. 三亚珊瑚变化趋势及原因分析. 海洋环境科学, 31(5): 682-685 |
吴钟解, 王道儒, 涂志刚, 等. 2011. 西沙生态监控区造礁石珊瑚退化原因分析. 海洋学报, 33(4): 140-146 |
吴钟解, 陈石泉, 陈敏, 等. 2013. 海南岛造礁石珊瑚资源初步调查与分析. 海洋湖沼通报, (2): 44-50 |
何晓娜, 吴钟解, 陈敏, 等. 2018. 海南七洲列岛周边珊瑚分布及其健康状况评价. 湿地科学与管理, 14(1): 11-15 |
邹仁林. 2001. 中国动物志:珊瑚虫纲. 北京: 科学出版社, 1-377
|
周红英, 姚雪梅, 黎李, 等. 2017. 海南岛周边海域造礁石珊瑚的群落结构及其分布. 生物多样性, 25(10): 1123-1130 |
郑洋, 倪建宇, 林钟扬, 等. 2015. 海南省三亚市亚龙湾珊瑚礁区水体环境特征研究. 海洋开发与管理, (9): 103-106 |
练健生, 黄晖, 黄良民, 等. 2010. 三亚珊瑚礁及其生物多样性. 北京: 海洋出版社, 1-106
|
赵美霞, 余克服, 张乔民. 2006. 珊瑚礁区的生物多样性及其生态功能. 生态学报, 26(1): 186-194 |
施祺, 赵美霞, 张乔民, 等. 2009. 海南三亚鹿回头造礁石珊瑚碳酸盐生产力的估算. 科学通报, 54(10): 1471-1479 |
徐兆礼, 陈亚瞿. 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 8(4): 13-15 |
海南省生态环境厅, 2019. 2018年海南省海洋环境状况公报. 2019. http://hnsthb.hainan.gov.cn/hjzl/hjzlxx/hjzkgb_51008/201907/t20190709_2626968.html
|
海南省海洋开发规划设计研究院, 中国科学院南海海洋研究所, 2011.三亚珊瑚礁国家级自然保护区综合科学考察报告(报批稿)
|
黄晖, 尤丰, 练健生, 等. 2011. 西沙群岛海域造礁石珊瑚物种多样性与分布特点. 生物多样性, 19(6): 710-715 |
黄晖, 尤丰, 练健生, 等. 2012. 海南岛西北部海域珊瑚礁造礁石珊瑚种类组成与分布. 海洋科学, 36(9): 64-74 |
梁文, 黎广钊, 张春华, 等. 2010. 20年来涠洲岛珊瑚礁物种多样性演变特征研究. 海洋科学, 34(12): 78-87 |
谢海群, 孙婆援, 庞巧珠, 等. 2016. 三亚珊瑚礁国家级自然保护区海域水质环境状况分析及评价. 热带农业科学, 36(9): 66-71 |
Bellwood D R, Hughes T P, 2001. Regional-scale assembly rules and biodiversity of coral reefs. Science, 292(5521): 1532-1535 |
Brandt M E, Olinger L K, Chaves-Fonnegra A et al, 2019. Coral recruitment is impacted by the presence of a sponge community. Marine Biology, 166(4): 49 |
Li X B, Huang H, Lian J S et al, 2013a. Coral community changes in response to a high sedimentation event:A case study in southern Hainan Island. Chinese Science Bulletin, 58(9): 1028-1037 |
Li X B, Huang H, Lian J S et al, 2013b. Spatial and temporal variations in sediment accumulation and their impacts on coral communities in the Sanya Coral Reef Reserve, Hainan, China. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 96: 88-96 |
Page C, Coleman G, Ninio R et al, 2001. Surveys of benthic reef communities using underwater video. Long-term monitoring of the Great Barrier Reef, standard operational procedure number 7. Townsville: Australian Institute of Marine Science, 45
|
Pielou E C, 1966. Species-diversity and pattern-diversity in the study of ecological succession. Journal of Theoretical Biology, 10(2): 370-383 |
Price N N, Muko S, Legendre L et al, 2019. Global biogeography of coral recruitment:tropical decline and subtropical increase. Marine Ecology Progress Series, 621: 1-17 |
Rogers C S, 1990. Responses of coral reefs and reef organisms to sedimentation. Marine Ecology Progress Series, 62: 185-202 |
Schlöder C, D'Croz L, 2004. Responses of massive and branching coral species to the combined effects of water temperature and nitrate enrichment. Journal of Experimental Marine Biology and Ecology, 313(2): 255-268 |
Shannon C E, 1948. A mathematical theory of communication. Bell System Technical Journal, 27(4): 623-656 |
Speare K E, Duran A, Miller M W et al, 2019. Sediment associated with algal turfs inhibits the settlement of two endangered coral species. Marine Pollution Bulletin, 144: 189-195 |
Yanovski R, Abelson A, 2019. Structural complexity enhancement as a potential coral-reef restoration tool. Ecological Engineering, 132: 87-93 |