 2020, Vol. 51
2020, Vol. 51

巢湖沿岸带底栖动物群落结构及其环境质量评价

http://dx.doi.org/10.11693/hyhz20191100222
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 赵永晶, 王洪铸, 崔永德. 2020.
- ZHAO Yong-Jing, WANG Hong-Zhu, CUI Yong-De. 2020.
- 巢湖沿岸带底栖动物群落结构及其环境质量评价
- COMMUNITY STRUCTURE OF MACROZOOBENTHOS AND ENVIRONMENTAL QUALITY ASSESSMENT IN LITTORAL ZONE OF CHAOHU LAKE
- 海洋与湖沼, 51(3): 520-527
- Oceanologia et Limnologia Sinica, 51(3): 520-527.
- http://dx.doi.org/10.11693/hyhz20191100222
文章历史
-
收稿日期:2019-11-22
收修改稿日期:2020-02-24
引用本文
赵永晶, 王洪铸, 崔永德. 2020. 巢湖沿岸带底栖动物群落结构及其环境质量评价[J]. 海洋与湖沼, 51(3): 520-527

ZHAO Yong-Jing, WANG Hong-Zhu, CUI Yong-De. 2020. COMMUNITY STRUCTURE OF MACROZOOBENTHOS AND ENVIRONMENTAL QUALITY ASSESSMENT IN LITTORAL ZONE OF CHAOHU LAKE[J]. Oceanologia et Limnologia Sinica, 51(3): 520-527.
巢湖沿岸带底栖动物群落结构及其环境质量评价
中国科学院水生生物研究所 淡水生态与生物技术国家重点实验室 武汉 430072
摘要:2013年3月、6月、9月和11月四个季度对巢湖沿岸带24个站点(12个区域)底栖动物进行调查,分析了群落结构特征并进行环境质量评价。本次调查共采集到底栖动物64属种,以寡毛类和水生昆虫为主,合计占总种类数的68.7%。底栖动物的平均密度和平均生物量(无灰干重)分别是(1005±233)ind./m2和(2.24±1.29)g/m2。巢湖沿岸带底栖动物优势类群为颤蚓类和摇蚊幼虫。利用底栖动物种类数、寡毛类密度占比、摇蚊幼虫密度占比和软体动物密度占比四个指标,对巢湖沿岸带环境质量进行综合评价,结果显示巢湖东岸环境质量优于西岸,靠近合肥市的万年埠处于严重污染状态。典型对应分析结果表明,影响底栖动物群落结构的主要环境因子有底质、电导率和总氮。建议通过改善底质环境,恢复水生植被,削减总氮等措施来恢复巢湖沿岸带底栖动物群落多样性,提高巢湖生态环境质量。
关键词:底栖动物 巢湖 沿岸带 群落结构 环境因子 评价
COMMUNITY STRUCTURE OF MACROZOOBENTHOS AND ENVIRONMENTAL QUALITY ASSESSMENT IN LITTORAL ZONE OF CHAOHU LAKE
State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
Abstract: Macrozoobenthos in the littoral zone of Chaohu Lake were investigated at 24 sites (12 regions) in four seasons of 2013 to understand the macrozoobenthos community structure and assess the environmental quality. Sixty-four taxa of macrozoobenthos were recorded, in which Oligochaetes and Aquatic Inscets were dominant, accounting for 68.7% of the taxa in number. The mean density and biomass (ash free dry weight) of macrozoobenthos were (1005±233)ind./m2 and (2.24±1.29)g/m2, respectively. The dominant taxa of macrozoobenthos were tubificids and chironomid larvae. The species number of macrozoobenthos and the ratios of Oligochaetes density, chironomid larvae density, and Molluscs density to the corresponding totals were used for the environmental quality assessment. Results show that the environmental quality of the eastern coast was better than the western coast, and that of Wannianbu near the Hefei City was poor as being heavily polluted. The substrate, conductivity, and total nitrogen were the main environmental factors affecting the macrozoobenthos community structure indicated by the CCA analysis. Therefore, to recovery the diversity of macrozoobenthos communities and improve the ecological environment quality, sediment environment shall be improved, aquatic vegetation restored, and total nitrogen reduced.
Key words:
macrozoobenthos Chaohu Lake littoral zone community structure environmental factors assessment
湖泊在维持生态系统能量转换、信息传递以及区域气候变化中起到了特殊生态作用(蔡阳等, 2019)。沿岸带是湖泊生态系统中复杂的区域, 是水生-陆地生态系统之间进行物质能量和信息交换的关键生物过渡区, 具有生境复杂、营养结构多元化和生物多样性高等特点(Kiss et al, 2003; Strayer et al, 2010)。由于人类及其他大型动物的频繁活动, 导致沿岸带水文剧烈波动, 水环境状态极其不稳定(Liboriussen et al, 2005; 杨凯等, 2013)。
巢湖是我国五大淡水湖之一, 位于皖中江淮之间(31°25′28″—31°43′28″N, 117°16′54″—117°51′46″E), 流域总面积1.35万平方公里, 东西长55公里、南北宽21公里, 湖岸线周长176公里, 是典型的大型浅水湖泊。历史上巢湖由于自然资源丰富、汇水面积广阔而素有“日出斗金”的美称。随着人口增长, 生活水平不断提高, 沿湖社会经济快速发展, 人为活动干扰日益频繁, 植被消亡和生态系统逐步退化, 环湖岸线和水向湖滨带丧失了削减污染、阻滞沉积物再悬浮等重要生态功能(王洪铸等, 2012)。
底栖动物是指生活史的全部或大部分时间在水体底部的一类水生生物, 是水生态系统中重要的生态类群(梁彦龄等, 1999)。底栖动物栖息地比较固定, 活动范围小, 生命周期相对较长, 对外界环境改变反应敏感, 以及对水质污染耐受程度不同, 通常可以用于环境质量现状评价(彭松耀等, 2017)。关于巢湖底栖动物生态调查的报道较多, 但大多研究集中在敞水区和入湖流域(胡菊英等, 1981; 高峰等, 2010; 宁怡等, 2012; 张又等, 2017), 专门针对沿岸带的研究较少(赵永晶等, 2012)。本文分析了巢湖沿岸带底栖动物群落现状和分布格局, 基于底栖动物对其环境质量进行综合评价, 并探讨了巢湖底栖动物群落健康的主要影响因素, 以期为恢复巢湖生态环境提供科学依据。
1 材料与方法 1.1 样点设置根据巢湖沿岸入湖河流的分布, 以及受周边城市、村镇等干扰程度, 同时结合沿岸带生境特点, 综合考虑以上因素, 在巢湖沿岸带布设12个区域(24个采样点)。小河村(S5)、兆河(S4)、槐林镇(S3)、散兵镇(S1)、中垾镇(S8)和小汤-邹梁村(S9)6个断面, 每个断面设置3个采样点, 自岸边至水向延伸, 即50、500、1000m;另设6个常规点, 靠近高林镇(S2)、浅水湾(S6)、小徐村(S11)、汪家嘴(S7)、黄麓镇(S10)和万年埠(S12)区域, 距岸边500m左右(图 1)。
 |
| 图 1 巢湖沿岸带采样点分布图 Fig. 1 Distribution of sampling in the littoral zone of Chaohu Lake |
底栖动物使用1/16m2改良彼德森采泥器进行采集, 每个采样点采两次。样品采集后经60目网筛洗净, 然后带回室内置于白色解剖盘中将样品分拣出来。随后放入标本瓶并用10%的甲醛溶液固定, 所有样品带回实验室进行镜检分类、计数称重。底栖动物湿重的测定是将样品放置在滤纸上吸干水分, 然后计数, 用精确度为万分之一电子天平称重, 根据干湿比换算系数将底栖动物生物量湿重换算为无灰干重(Wetzel et al, 2005; Edwards et al, 2009; 赵伟华, 2010)。标本鉴定参考国内外相关书籍(刘月英等, 1979; 蔡如星等, 1991; Morse et al, 1994; 王洪铸, 1995; 王俊才等, 2011; 何雪宝, 2011), 尽可能鉴定到科、属、种水平。
使用便携式YSI多参数水质监测仪(美国)现场测定气温(TT)、水体的温度(WT)、酸碱度(pH)、溶解性固体总量(TDS)、溶解氧(DO)、电导率(Cond)、浊度(Turb)等理化指标。水深使用测深仪测定, 透明度的测量使用Secchi盘。现场采集的水样带回实验室, 用于测量水体总磷(TP)、总氮(TN)、氨氮(NH4+)、硝氮(NO3-)和叶绿素a (chl a)等质量浓度, 测定方法参考《水和废水监测分析方法》(2002)。根据粒径大小, 参考相关资料, 将底质分为砾石、粗砂、细砂、黏土、淤泥, 分别赋值5、4、3、2、1(鲁如坤, 1999; Allan et al, 2007)。
1.3 数据处理与分析采用EXCEL 2016和ArcGIS 10.1软件分析绘制大型底栖动物种类数、密度和生物量分布图。采用CANOCO 5.0软件的典型对应分析(canonical correspondence analysis, CCA)筛选影响底栖动物群落的关键环境因子, 保留能通过蒙特卡罗置换检验的显著影响因子(Monte Carlo test, 9999 random permutations, P < 0.05)。采用EXCEL 2016和多元统计软件PRIMER 6.1计算大型底栖动物群落优势度指数(Y), 计算公式如下:
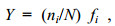 (1)
(1)式中, N为总个体数, ni为第i种的个体数, fi为第i种在各位点的出现率。Y > 0.02的种类确定为优势种。
本文选取底栖动物种类数、寡毛类密度所占比例、摇蚊类群密度所占比例和软体动物密度所占比例四个指数, 对沿岸带环境质量进行综合评价。评价标准见表 1。
表 1 巢湖沿岸带底栖动物各生物指标评价标准 Tab. 1 Assessment criteria for the biological metrics of macrozoobenthos in the littoral zone of Chaohu Lake
| 生物指标 | 优(得分) | 良(得分) | 中(得分) | 差(得分) | 极差(得分) |
| 总种类数(指标Ⅰ) | > 30 | 26—30 | 22—26 | 18—22 | < 18 |
| 寡毛类所占比例(指标Ⅱ) | < 25% | 25%—40% | 40%—55% | 55%—70% | > 70% |
| 摇蚊类所占比例(指标Ⅲ) | < 25% | 25%—40% | 40%—55% | 55%—70% | > 70% |
| 软体动物所占比例(指标Ⅳ) | > 25% | 20%—25% | 15%—20% | 10%—15% | < 10% |
巢湖沿岸带24个采样点中共采集到底栖动物64种, 其中寡毛类21种(占总种类数32.8%), 水生昆虫23种(占总种类数35.9%), 软体动物13种(占总种类数20.4%), 其他类群7种(占总种类数10.9%)(图 2)。
 |
| 图 2 巢湖沿岸带底栖动物种类数、密度和生物量 Fig. 2 Species, density and biomass of macrozoobenthos in the littoral zone of Chaohu Lake |
各区域大型底栖动物种类变化幅度为15—33种。不同区域来看, 出现频率较高的均为耐污种类, 如水丝蚓属(Limnodrilus sp.)、厚唇嫩丝蚓(Teneridrilus mastix)、摇蚊属(Chironomus sp.)、小摇蚊属(Microchironomus sp.), 出现频率均为100%。就分布而言, 巢湖东北沿岸物种较为丰富, 尤其是小汤-邹梁村(S9)区域, 采集到33种。整体来看, 巢湖东岸底栖动物种类数高于西岸。
巢湖沿岸带底栖动物平均密度为(1005± 366)ind./m2, 其中水生昆虫为优势类群, 总密度为(656±273)ind./m2 (占总密度的65%), 其次为寡毛类, 占总密度31%, 软体动物和其他类群合计占4%。从不同区域看, 靠近巢湖市和合肥市区域底栖动物密度较高, 尤其是靠近巢湖市的汪家嘴断面, 密度高达3488ind./m2。巢湖南岸如小河村断面、兆河河滩断面及槐林镇断面等密度较低(图 2)。
底栖动物平均生物量(无灰干重, 下同)为(2.24± 1.29)g/m2, 其中软体动物生物量最高, 为(1.64± 1.13)g/m2, 占总生物量73.0%, 其次为水生昆虫, 占总生物量18.4%, 寡毛类和其他类群合计占8.6%。各区域底栖动物生物量的分布与密度分布存在较大的差异性, 高生物量主要分布在浅水湾、散兵镇、槐林镇和汪家嘴断面等, 低生物量分布区基本与低密度分布区较一致, 如高林镇、小河村和兆河断面(图 2)。
计算优势度, 以Y > 0.02为标准, 巢湖沿岸带底栖动物优势类群为寡毛类和水生昆虫。优势种为水丝蚓属、厚唇嫩丝蚓、苏氏尾鳃蚓(Branchiura sowerbyi)、摇蚊属、小摇蚊属、裸须摇蚊属(Propsilocerus sp.)和长足摇蚊属(Tanypus sp.)。
2.2 影响底栖动物群落的关键环境因子采样点的环境参数采用四季平均值, 测定结果为:水温(WT)(16.79±3.37)℃, 气温(TT)(18.51± 3.59)℃, 溶氧(DO)(10.12±0.97)mg/L, pH值8.63±0.22, 电导率(Cond)(318.91±9.95)μS/cm, 透明度(ZSD) (41.23±4.39)cm, 水深(SS)(1.64±0.22)m, 溶解性固体总量(TDS)(0.21±0.01)mg/L, 浊度(Turb)(40.45± 8.17)NTU, 总氮(TN)(2.44±0.83)mg/L, 总磷(TP) (0.13±0.12)mg/L, 氨氮(NH4+)(0.49±0.19)mg/L, 硝氮(NO3-)(0.99±0.32)mg/L, 叶绿素a (chl a)(36.00± 18.97)μg/L。本研究对底栖动物各类群密度与15个环境因子之间进行典型对应分析, 筛选出影响底栖动物群落的关键环境因子。结果表明, 影响底栖动物的主要环境因子有底质粒径、总氮和电导率(P < 0.05) (图 3)。第一轴和第二轴的特征值较大, 分别解释了46.02%和18.23%的类群数据方差变异和59.97%和23.75%的类群-环境关系变异。底栖动物不同类群之间的差异较大。寡毛类密度分布与总氮和溶解性固体总量的关系最密切, 都呈正相关。软体动物密度分布与底质粒径和电导率关系最为密切, 与前者呈正相关, 与后者呈负相关关系。水生昆虫密度分布与其他几个类群相反, 与总氮和浊度关系最密切, 且都呈负相关。其他类群中的蛭类、甲壳类等与底质粒径和电导率的关系最密切, 且与前者正相关, 后者负相关。
 |
| 图 3 底栖动物密度与环境因子的典型对应分析排序图 Fig. 3 CCA plots of macrozoobenthos density and environmental factors 注: Sed:底质; Turb:浊度; TN:总氮; TDS:溶解性固体总量; Cond:电导率; Molluscs:软体动物; Aquatic Insects:水生昆虫; Oligochaetes:寡毛类; Others:其他类群; Total:合计 |
通过全年四次野外调查, 选取底栖动物种类数、寡毛类密度所占比例、摇蚊类群密度所占比例和软体动物密度所占比例四个指数, 对沿岸带环境质量进行综合评价。依据调查数据, 各指标均分为5个等级, 即优、良、中、差及极差, 分别赋值100、80、60、40及20分, 再将四个指标进行综合, 依据综合后的分值即为各水体划分等级的依据, 为0—20、21—40、41—60、61—80、81—100分, 依次对应的等级为极差、差、中、良和优。依据上述四个指标对巢湖沿岸带环境质量状况进行综合评价, 在调查范围内暂未发现环境质量为优的区域; 评价为良的兆河河口和槐林镇等; 散兵镇、高林镇和邹梁村等水质处于中等污染状态; 评价结果较差的区域有黄麓镇和万年埠; 尤其是靠近合肥市的万年埠, 位于派河流域附近, 环境质量接近重度污染状态(图 4)。
 |
| 图 4 巢湖沿岸带不同区域综合评价结果 Fig. 4 Comprehensive assessment in the littoral zone of Chaohu Lake in different regions |
本次调查共发现底栖动物64种, 主要由寡毛类和水生昆虫组成。优势种主要以水丝蚓属、厚唇嫩丝蚓、苏氏尾鳃蚓、摇蚊属、裸须摇蚊属和长足摇蚊属等耐污种为主。本研究与胡菊英等(1981年)、龚志军等(2001年)和赵永晶等(2012年)的研究相比较, 整体来看, 巢湖底栖动物总种类数是先减少后增加, 寡毛类和水生昆虫的种类数明显增加, 软体动物的种类数由最初的29种下降到13种, 而在2009年沿岸带的调查中仅发现了5种。系统比较2009年与2013年夏秋季巢湖沿岸带底栖动物群落结构特征变化发现, 本次调查底栖动物种类数、密度(38种, 670ind./m2)与2009年(33种, 506ind./m2)相比变化不大, 但生物量却不到2009年的1/2, 优势种由2009年的水丝蚓属、菱跗摇蚊属、河蚬和环棱螺属等演变为目前的颤蚓类和摇蚊幼虫等(图 5)。有研究表明, 随着水体富营养化程度加剧, 底栖动物敏感种逐渐消失, 寡毛类快速繁殖(Dalu et al, 2017)。底栖动物总数量虽然变化不大, 但由于生物量主要贡献者软体动物数量明显下降, 从而导致总生物量显著下降。巢湖湖滨带水生植物逐渐减少是造成软体动物变化的主要原因。水生植被减少的主要原因分析如下:一是建闸导致水位变化(俞汉青, 1992)和堤岸硬化。有研究表明水位的不正常波动(长期高水位、长期低水位或非周期性波动)会对湖滨带植物的分布格局及多样性产生较大影响(Poff et al, 1997; Hebb, 2003; 丁庆章, 2013)。自巢湖闸和裕溪闸建成后, 巢湖水位波幅大幅度减小, 波动型也发生了改变, 导致挺水植物难以萌发, 最终导致水生植被盖度由25%以上减少到不足全湖面积的1% (刘学勤等, 2012)。二是以往采取抛石加固、构建护坡等物理生态修复措施, 尤其是水泥堤岸的修筑导致自然生境丧失, 生态环境遭破坏(吴志华等, 2006)。王洪铸等(2012)研究表明, 不同岸线应采取不同的修复技术, 比如采用生态混凝土进行绿化, 通过在水泥造孔等方式改变基底, 并对现有的自然岸线给以充分保护, 从而构建健康的生态修复带, 提高生物多样性。
 |
| 图 5 巢湖底栖动物历史变化 Fig. 5 The history variation of macrozoobenthos in the littoral zone of Chaohu Lake |
底栖动物各个类群对外界环境变化的适应性及对污染程度的耐受性和敏感性存在一定差异, 因此底栖动物群落结构、现存量及生物多样性均可随着环境的变化而改变。本研究表明, 影响底栖动物的关键环境因子有底质、总氮和电导率。
一般认为底栖动物的现存量与底质的稳定性和有机质含量关系密切(Quinn et al, 1994), 如粗砂和细砂底质不稳定, 底栖动物数量偏低; 黏土和淤泥有机碎屑含量高, 食物来源丰富, 故密度偏高。汪家嘴、中垾镇和万年埠区域底质为淤泥, 采集到大量寡毛类, 密度高达456、740和600ind./m2。由于巢湖沿岸带底栖动物密度受寡毛类和水生昆虫密度支配, 在底质以淤泥为主且较为稳定的区域最高; 而生物量取决于软体动物的多少, 因此在以细砂和粗砂底质为主的区域(槐林镇和浅水湾)较高。本研究区域位于岸边50—1000m范围内, 浅水沿岸带以其复杂的基质提供了多样化的生境, 相比底质单一、生境简单的敞水区, 底栖动物数量偏多。
总氮含量可反映水体的污染程度和营养状态(吴召仕等, 2011)。龚志军等(2001)研究表明, 湖泊富营养化会直接导致底栖动物多样性下降、现存量减少。近年来, 随着巢湖流域经济的快速发展, 工业生产迅猛发展及城市化进程不断加快, 蓝藻水华频繁暴发, 巢湖水体富营养化程度日益加剧, 从而导致底栖动物群落结构单一, 敏感种逐渐下降直至消失, 取而代之的是耐污种增加。
电导率作为指示水生生态系统健康状态的指标之一, 同时也是影响底栖动物群落结构的主要因子(Berger et al, 2017)。水体电导率与盐度、温度等存在着一定的关系, 用电导率测定盐度的方法已成为日常广泛应用的方法之一(陈国华等, 1980)。有研究表明人为活动干扰如污水排放、堤岸修复及路基修建等会导致水体电导率的迅速升高(Leps et al, 2015)。近年来, 巢湖周边生产生活等各种原因导致水质严重污染, 由此开展了大量巢湖湖滨带生态修复工程(刘学勤等, 2012)。本研究发现沙蚕科等咸淡水种类主要分布在盐度相对较高的区域, 也说明与以上观点相符。
3.3 环境质量评价基于理化指标的水质评价更多关注水体中营养盐和污染物浓度与环境阈值的关系, 而利用水生生物评价除了反映水质质量, 还考虑了生境信息(底质、栖息地环境、植被覆盖度、人类活动干扰强度等), 因此其评价结果更能准确全面地反映环境质量优良(周笑白等, 2014)。以底栖动物作为环境质量评价指标较为常见, 并被广泛认可和应用(王备新等, 2001; 蔡琨等, 2014)。本研究通过多指标综合评价显示, 巢湖沿岸带水体处于中-富营养化状态。整体来看, 巢湖西半湖沿岸带污染比东半湖严重, 主要是因为西半湖靠近合肥市、肥西县等城市, 城市工业废水、生活污水等大量排入, 水体氮磷含量明显高于其他区域。余秋梅等(2001)研究结果表明, 西半湖水质为Ⅳ类的监测点所占比例较高, 东半湖水质为Ⅱ—Ⅲ类占比相对较高。不同区域水质状况有所不同, 其中兆河河口和槐林镇断面水生植被种类最为丰富, 覆盖度亦较高(未发表数据), 该区域为轻污染; 散兵镇、高林镇和邹梁村等评价为中污染; 第三类较差水体为黄麓镇和万年埠, 尤其是万年埠, 评价得分接近极差。理化数据显示, 靠近合肥市的万年埠总氮(6.8mg/L)、总磷(0.3mg/L)及叶绿素a (62.55μg/L)含量极高。王书航等(2011)研究表明, 南淝河和派河水质状况为劣Ⅴ类, 属于重污染; 兆河、柘皋河水质现状为Ⅲ—Ⅳ类, 水质轻度污染, 裕溪闸下段主要为Ⅱ—Ⅲ类(张又等, 2017)。这些入湖流域的水质评价结果与入湖区域的评价结果吻合度较高, 说明巢湖全湖的营养状况与入湖流域水质的污染状况有着紧密的关系。
4 结论本文于2013年四个季度对巢湖沿岸带12个区域底栖动物进行采样调查, 共发现底栖动物64属种, 优势种为水丝蚓属、厚唇嫩丝蚓、摇蚊属、裸须摇蚊属和长足摇蚊属等。巢湖沿岸带底栖动物平均密度和平均生物量(无灰干重)分别是(1005±233)ind./m2和(2.24±1.29)g/m2。利用底栖动物种类数、寡毛类密度所占比例、摇蚊幼虫密度所占比例和软体动物密度所占比例四个指标, 对巢湖沿岸带环境质量进行综合评价, 结果表明巢湖东岸环境质量优于西岸, 靠近合肥市的万年埠处于严重污染状态。典型对应分析结果表明, 影响底栖动物群落结构的主要环境因子有底质、总氮和电导率(P < 0.05)。巢湖沿岸带底栖生态系统修复应考虑改善底质环境, 恢复水生植被, 削减总氮。
致谢 感谢彭玉在标本鉴定过程中给予帮助, 感谢和雅静、吴俊燕在数据处理方面给予帮助。
参考文献
丁庆章, 2013.水位波动对湖滨带植被基底环境的影响.北京: 中国科学院大学硕士学位论文
|
王书航, 姜霞, 金相灿. 2011. 巢湖入湖河流分类及污染特征分析. 环境科学, 32(10): 2834-2839 |
王备新, 杨莲芳. 2001. 大型底栖无脊椎动物水质快速生物评价的研究进展. 南京农业大学学报, 24(4): 107-111 |
王俊才, 王新华. 2011. 中国北方摇蚊幼虫. 北京: 中国言实出版社
|
王洪铸, 1995.中国小蚓类研究-附中国南极长城站附近地区两新种.武汉: 中国科学院水生生物研究所博士学位论文
|
王洪铸, 宋春雷, 刘学勤, 等. 2012. 巢湖湖滨带概况及环湖岸线和水向湖滨带生态修复方案. 长江流域资源与环境, 21(S2): 62-68 |
宁怡, 高峰, 邓建才, 等. 2012. 巢湖流域水质生物学评价——以大型底栖动物为例. 生态学杂志, 31(4): 916-922 |
刘月英, 张文珍, 王跃先, 等. 1979. 中国经济动物志:淡水软体动物. 北京: 科学出版社
|
刘学勤, 邢伟, 张晓可. 2012. 巢湖水向湖滨带生态修复工程实践. 长江流域资源与环境, 21(S2): 51-55 |
杨凯, 董静, 郭亮亮, 等. 2013. 滇池沿岸带刚毛藻种群结构及分布特征. 水生态学杂志, 34(3): 8-16 DOI:10.3969/j.issn.1674-3075.2013.03.002 |
吴召仕, 蔡永久, 陈宇炜, 等. 2011. 太湖流域主要河流大型底栖动物群落结构及水质生物学评价. 湖泊科学, 23(5): 686-694 |
吴志华, 王晓辉. 2006. 巢湖东端湖滨带物理基底及生态修复. 合肥工业大学学报(自然科学版), 29(9): 1068-1071, 1076 DOI:10.3969/j.issn.1003-5060.2006.09.003 |
何雪宝, 2011.西藏和四条大型河流水栖寡毛类区系研究.武汉: 中国科学院水生生物研究所博士学位论文
|
余秋梅, 印士勇, 周良伟. 2001. 巢湖水环境质量现状分析. 人民长江, 32(7): 29-30 DOI:10.3969/j.issn.1001-4179.2001.07.010 |
张又, 程龙, 伊洪斌, 等. 2017. 巢湖流域不同水系大型底栖动物群落结构及影响因素. 湖泊科学, 29(1): 200-215 |
陈国华. 1980. 中国标准海水电导盐度的标定. 海洋与湖沼, 11(2): 115-120 |
周笑白, 张宁红, 张咏, 等. 2014. 太湖水质与水生生物健康的关联性初探. 环境科学, 35(1): 271-278 |
赵永晶, 刘学勤. 2012. 巢湖水向湖滨带底栖动物群落结构研究. 长江流域资源与环境, 21(S2): 39-44 |
赵伟华, 2010.中国河流底栖动物宏观格局及黄河下游生态需水研究.武汉: 中国科学院水生生物研究所博士学位论文
|
胡菊英, 姚闻卿. 1981. 巢湖底栖动物调查. 安徽大学学报(自然科学版), (2): 159-173 |
俞汉青. 1992. 巢湖水资源开发问题的探讨. 自然资源, (6): 27-31 |
高峰, 尹洪斌, 胡维平, 等. 2010. 巢湖流域春季大型底栖动物群落生态特征及与环境因子关系. 应用生态学报, 21(8): 2132-2139 |
龚志军, 谢平, 唐汇涓, 等. 2001. 水体富营养化对大型底栖动物群落结构及多样性的影响. 水生生物学报, 25(3): 210-216 DOI:10.3321/j.issn:1000-3207.2001.03.002 |
梁彦龄, 王洪铸. 1999. 底栖动物(第十章).见:刘建康.高级水生生物学. 北京: 科学出版社, 241-259
|
彭松耀, 李新正, 徐勇, 等. 2017. 十年间黄海大型底栖动物优势种的变化. 海洋与湖沼, 48(3): 536-542 |
鲁如坤. 1999. 土壤农业化学分析方法. 北京: 中国农业科技出版社
|
蔡阳, 陆欣鑫, 巴秋爽, 等. 2019. 镜泊湖春、夏两季浮游生物群落结构及其与环境因子的关系. 海洋与湖沼, 50(1): 116-128 |
蔡琨, 张杰, 徐兆安, 等. 2014. 应用底栖动物完整性指数评价太湖生态健康. 湖泊科学, 26(1): 74-82 |
蔡如星. 1991. 浙江动物志:软体动物. 杭州: 浙江科学技术出版社
|
Allan J D, Castillo M M, 2007. Stream Ecology: Structure and Function of Running Waters. Dordrecht: Springer
|
Berger E, Haase P, Kuemmerlen M et al, 2017. Water quality variables and pollution sources shaping stream macroinvertebrate communities. Science of the Total Environment, 587-588: 1-10 DOI:10.1016/j.scitotenv.2017.02.031 |
Dalu T, Wasserman R J, Tonkin J D et al, 2017. Water or sediment? Partitioning the role of water column and sediment chemistry as drivers of macroinvertebrate communities in an austral South African stream. Science of the Total Environment, 607-608: 317-325 DOI:10.1016/j.scitotenv.2017.06.267 |
Edwards F K, Lauridsen R B, Armand L et al, 2009. The relationship between length, mass and preservation time for three species of freshwater leeches (Hirudinea). Fundamental and Applied Limnology, 173(4): 321-327 |
Hebb A, 2003. Implementation of a GIS to assess the effects of water level fluctuations on the wetland complex at long point, Ontario. Waterloo: Master Dissertation of University of Waterloo
|
Kiss M K, Lakatos G, Borics G et al, 2003. Littoral macrophyte-periphyton complexes in two Hungarian shallow waters. Hydrobiologia, 506-509(1-3): 541-548 DOI:10.1023/B:HYDR.0000008594.48184.ca |
Leps M, Tonkin J D, Dahm V et al, 2015. Disentangling environmental drivers of benthic invertebrate assemblages:the role of spatial scale and riverscape heterogeneity in a multiple stressor environment. Science of the Total Environment, 536: 546-556 |
Liboriussen L, Jeppesen E, Bramm M E et al, 2005. Periphyton-macroinvertebrate interactions in light and fish manipulated enclosures in a clear and a turbid shallow lake. Aquatic Ecology, 39(1): 23-39 |
Morse J C, Yang L, Tian L, 1994. Aquatic Insects of China Useful for Monitoring Water Quality. Nanjing: Hohai University Press
|
Poff N L, Allan J D, Bain M B et al, 1997. The natural flow regime. BioScience, 47(11): 769-784 |
Quinn J M, Hickey C W, 1994. Hydraulic parameters and benthic invertebrate distributions in two gravel-bed New Zealand rivers. Freshwater Biology, 32(3): 489-500 DOI:10.1111/j.1365-2427.1994.tb01142.x |
Strayer D L, Findlay S E G, 2010. Ecology of freshwater shore zones. Aquatic Sciences, 72(2): 127-163 |
Wetzel M A, Leuchs H, Koop J H E, 2005. Preservation effects on wet weight, dry weight, and ash-free dry weight biomass estimates of four common estuarine macro-invertebrates:no difference between ethanol and formalin. Helgoland Marine Research, 59(3): 206-213 |