 2020, Vol. 51
2020, Vol. 51中国海洋湖沼学会主办。
文章信息
- 武亚男, 张昊飞, 王璐, 李正炎. 2020.
- WU Ya-Nan, ZHANG Hao-Fei, WANG Lu, LI Zheng-Yan. 2020.
- 长江口无机氮和活性磷酸盐水质基准研究
- WATER QUALITY CRITERIA OF DISSOLVED INORGANIC NITROGEN AND SOLUBLE REACTIVE PHOSPHORUS IN CHANGJIANG RIVER ESTUARY
- 海洋与湖沼, 51(5): 1066-1079
- Oceanologia et Limnologia Sinica, 51(5): 1066-1079.
- http://dx.doi.org/10.11693/hyhz20200400109
文章历史
-
收稿日期:2020-04-05
收修改稿日期:2020-05-26




2. 国家海洋局东海环境监测中心 上海 201206;
3. 自然资源部海洋生态监测与修复技术重点实验室 上海 201206;
4. 中国环境科学研究院 北京 100012;
5. 中国海洋大学海洋环境与生态教育部重点实验室 青岛 266100
2. East China Sea Environment Monitoring Center, SOA, Shanghai 201206, China;
3. Key Laboratory of Marine Ecological Monitoring and Restoration Technology, Ministry of Natural Resources, Shanghai 201206, China;
4. Chinese Research Academy of Environmental Sciences, Beijing 100012, China;
5. Key Laboratory of Marine Environmental Science and Ecology, Ministry of Education, Qingdao 266100, China
受淡水径流和海洋潮汐共同作用影响, 河口作为咸淡水的“过滤器”, 盐度变化剧烈, 生态系统复杂敏感(沈焕庭等, 2001; 陆健健, 2003)。氮、磷等营养物质大量输入容易造成河口及其邻近海域富营养化, 藻类大量繁殖, 破坏生态系统完整性(李俊龙等, 2015)。作为我国第一大河口, 长江口紧邻上海、浙江和江苏三个重要经济区, 氮、磷等营养盐的污染持续加重, 近50年来, 长江口门区域可溶性无机氮(dissolved inorganic nitrogen, DIN)的浓度升高了四倍以上, 可溶性活性磷酸盐(soluble reactive phosphorus, SRP)升高了两倍左右, 尤其在2000年以后, 赤潮事件暴发频率和面积显著上升, 对生态安全、海水养殖业和人类健康都构成了严重威胁(刘录三等, 2011a; 张璇, 2012; 于仁成等, 2017)。但目前我国尚没有适用于河口环境的水质标准, 现行的《地表水环境质量标准》(GB 3838-2002)和《海水水质标准》(GB 3097-1997)难以满足河口区的管理和保护(孟伟等, 2006)。因此, 制定符合我国区域特点的河口营养物基准十分必要, 这不仅是营养物监测、评价和管理的科学依据, 而且可以为营养物标准的制定提供理论和方法(Wu et al, 2010)。
河口营养物基准(nutrient criteria in estuarine waters)是指河口营养状态参数对生态环境不产生不良或有害影响的最大阈值(USEPA, 1998)。美国系统地研究了河流、湖泊和水库、河口和近岸海域以及湿地等生态系统营养物基准的理论方法, 相继颁布了不同环境类型的营养物基准技术指南, 并对佛罗里达州19个河口的营养物基准进行了初步研究, 在生态学分区的基础上, 主要运用频数分布法和压力-响应模型法对营养物基准值进行推导和验证, 目前已经形成比较成熟的营养物基准体系(USEPA, 2000a, 2000b, 2001, 2008)。
在这些理论方法的基础上, 我国学者也在探索河口营养基准和标准制定技术。我国河口众多, 河口的分类和分区是营养物基准制定首先需要考虑的因素, 也是河口近岸海域环境管理的重要手段之一(郑丙辉等, 2013)。营养物基准制定过程中, 频数分布法和压力-响应模型是最常用的方法, 前者利用收集到的河口历史和现状数据进行统计分析确定营养物基准值, 后者通过建立营养指标与响应指标的关系来确定营养指标阈值(霍守亮等, 2017)。本文以我国长江口为研究对象, 根据河口内部环境特征开展生态学分区, 在此基础上, 运用频数分布法和压力-响应模型对不同分区的营养指标基准值进行推导, 将赤潮优势藻培养实验得到的生态响应值作为参考。
1 材料与方法 1.1 研究区域和数据来源长江口是中国的第一大河口, 位处东海和黄海的分界, 整体呈现三级分汊、四口入海的格局, 窄口端江面宽度5.8公里, 宽口江面宽度90公里, 其邻近海域相对开阔, 水深变化较为明显, 在123°E以西的区域内, 水深在60m以内, 而在123°E以东的海域, 水深可达到90m (中国海湾志编纂委员会, 1998)。长江口水域是长江径流和近岸海域多种暖流、水团的交汇区, 气象地形条件和生物化学特征都比较复杂(苏纪兰, 2001; Wei et al, 2007)。
我国学者将长江口的范围划定为自江阴(120.26°E)到水下三角洲前缘(123°E), 按普理查德理论来看, 长江口上缘应为徐六泾(121°E), 所以本研究的研究区域定为30°30′-32°00′N, 121°00′-123°00′E(沈焕庭等, 2003)。研究数据主要来源于原国家海洋局海洋环境调查数据。长江口盐度空间分布分析主要基于2016和2017年33个采样点的表底层数据(图 3); 分区检验运用2018年3月、5月、8月和10月96个站位的调查数据, 调查指标包括水温、pH、溶解氧(DO)、化学需氧量(COD)、总氮(TN)、总有机碳(TOC)等; 营养物基准指标研究运用了2016-2017年DIN、SRP、pH、DO、悬浮物(SS)、叶绿素a (chl a)和盐度(S)数据; 基准推导采用2015-2018年春夏秋各季节的监测数据。监测站位覆盖了长江口的咸淡水混合区, 所有监测指标的样品采集、预处理、分析与鉴定均按《海洋监测规范》(GB 17378.4-2007)等相关标准方法进行。
 |
| 图 3 长江口不同年份盐度分布 Fig. 3 Saltine distribution of Changjiang (Yangtze) River estuary in different years 注: a. 2016年表层; b. 2016年底层; c. 2017年表层; d. 2017年底层 |
河口生态分区的出发点是河口生态系统对营养物的敏感性, 河口区域的盐度和地理形态特征是影响其敏感性的主要自然因素(Liu et al, 2018)。地理形态特征是流体动力学和混合因素的产物, 盐度是生物地球化学过程的主要控制参数, 因此, 这些因素被认为是河口生态分区的主要因素(刘录三等, 2011b)。另外, 基于每个采样点的盐度值, 运用Surfer中的内插法对离散数据格网化获得盐度变化的空间模式。在此基础上, 运用单因素方差分析(One-way ANOVA), 对水体多参数指标进行分区之间的差异显著性检验, 以验证分区的合理性。
1.3 营养物基准指标研究结合调查数据, 运用相关性分析(correlation analysis, CA)、主成分分析(principal component analysis, PCA)以及线性回归分析(linear regression analysis, LRA)等方法进行分析。相关性分析是研究两两指标的相关性, 通常用Pearson相关系数评价相关性; 主成分分析是通过正交变换将一组可能存在相关性的变量转换为一组线性不相关的变量, 转换后的这组变量叫主成分, 以此来实现降维的效果; 线性回归是利用线性回归方程的最小平方函数对一个或多个自变量和因变量之间关系进行建模的一种回归分析。通过以上方法来分析与营养状况显著相关的因子, 进而确定长江口水体营养物基准的关键指标。
1.4 基准推导方法频数分布法是对营养物数据进行统计分析, 基准值的得出是基于区域内生境状况的判断, 生境状况有完好、部分退化和完全退化三类。参照点一般选取区域内受人类活动损害最小且对该水体或邻近水体的生物学完整性具有代表性的点位, 以此来衡量区域内相对未受干扰的营养状态(USEPA, 1998)。将营养物数据按从小到大的顺序分别排列, 对于生境状况完好的区域, 选择所有参照点数据分布曲线的中值作为基准值; 对于生境部分退化, 但参照点数量充足(大于等于所有观测点数量的10%)的区域, 取参照点营养指标频数分布曲线的上25%数值作为基准值; 对于生境部分退化, 参照点不足(小于所有观测点数量的10%)的区域, 取所有观测点营养指标频数分布曲线的下25%数值作为基准值, 需要指出的是, 对于原始数据大多处于退化状态, 可选取5%作为基准值; 而生境完全退化的区域, 不能使用频数分布法(图 1)。
 |
| 图 1 频数分布法推导营养物基准技术流程 Fig. 1 Process of deriving nutrient criteria by frequency distribution method |
压力-响应模型通过构建原因变量(TN、TP、DIN、SRP等)与响应变量(chl a、DO、TOC、透明度等)之间的关系模型来确定营养物基准值。Chl a是反映藻类生物量最常用的指标, DO是生态健康和栖息地功能评价的重要指标, 受水体扰动、光强和温度、浮游植物及大型动植物的种类和生物量等因素影响, 本研究将两者作为响应指标以建立压力-响应模型(USEPA, 2010; 李薇, 2014)。
Chl a和DO含量与营养物浓度之间的关系比较细微, 而分类回归树(classification and regression tree, CART)是解决响应变量和原因变量之间非线性、分层及高阶关系的有效方法, 并能测定导致生态发生变化的数字化数值(De'Ath et al, 2000)。分类回归树是一种非参数模拟方法, 其通过构建一个二元递归函数, 将由自变量定义的因变量划分为尽可能同质的类别, 每一次划分都由自变量的一次最佳划分值来完成, 分组后响应指标对应的最大降幅或变异性被分成两个独立的节点, 重复此过程, 直到形成一组按照某种标准使节点无法再分的终节点。本研究采用CART模型来揭示原因指标与响应变量之间复杂的相互作用关系, 从而确定营养物基准值。长江口3-10月份有发生赤潮的历史记录, 所以对2016-2018年3-10月的数据进行CART分析(刘录三等, 2011a)。
以上的营养物基准指标、频数分布统计法和分区检验(One-way ANOVA)的分析和作图在SPSS 24.0和Origin 9.2软件中进行, 运用R语言(R x64 3.5.1)实现CART的建立。
1.5 生态响应值推导方法 1.5.1 研究方法本校验试验以中肋骨条藻(Skeletonema costatum)和东海原甲藻(Prorocentrum donghaiense)为研究藻种。中助骨条藻属于温带和热带海洋硅藻类, 有广温广盐的特征, 虽然无毒, 但增殖速率很快, 春夏秋季均是其繁殖高峰期(王江涛等, 2012; 中华人民共和国自然资源部, 1989-2019)。中肋骨条藻是长江口的赤潮种之一, 温度25℃左右和盐度14-20条件下最常见, 是口门区赤潮的主要优势藻(李照, 2018)。东海原甲藻属温带甲藻类, 最适温度为20℃左右, 最适盐度为25-34, 是我国长江口及其邻近海域发生赤潮事件的关键种, 与中肋骨条藻相比, 东海原甲藻赤潮面积更大, 主要在口外区出现, 持续时间更长, 会导致爆发海域的大面积低氧现象, 对其海洋生物和区域内生态系统产生极其不良的影响(陈翰林等, 2006; 张清春, 2011; 戴鑫烽等, 2014; 黄备等, 2014)。
根据长江口现场调查监测数据, 设置合适的氮磷浓度, 以S-logistic 1模型拟合浮游藻生长曲线, 具体形式如下:
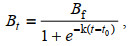 (1)
(1)式中, Bt代表t时浮游藻的生物量(cells/mL), Bf代表终止生物量, 即浮游藻生长达到稳定时的生物量(cells/mL), k为S-logistic 1模型参数, t为培养时间(d), t0是拐点处的培养时间, 即Bt=Bf/2时的培养时间。
营养物敏感性是指水生态系统对营养物浓度和结构变化响应的敏感程度, 用生态学指标的数值随营养物浓度变化而变化的规律来确定(张远等, 2006)。本研究中浮游藻终止生物量随营养盐浓度的变化规律基本呈“S”形, 也采用S-logistic 1来拟合, 具体形式见公式(2), 生态响应值的计算方法如图 2。
 |
| 图 2 生态响应值的计算方法 Fig. 2 Calculation method of ecological response value |
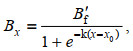 (2)
(2)式中, B'f表示每个梯度任意抽取的Bf值, x为营养盐浓度值(mg/L), x0为Bt=Bf/2时营养盐的浓度值(mg/L), 将营养盐浓度控制在x0以下, 既能保证正常的初级生产量, 又能避免浮游藻的过量繁殖, 所以本研究中将x0作为生态响应值。以上的随机抽样步骤在MATLAB R2014b软件中进行, 曲线拟合和拐点值的计算运用Origin Pro 9.2来实现。
1.5.2 浮游藻的培养浮游藻的培养采用人工海水(石岩峻, 2004), 将其置于2L的锥形瓶中, 中肋骨条藻的培养海水盐度调至20, 东海原甲藻盐度为30, 以分别接近两分区的实际海水盐度和最适生长盐度, 高压蒸汽灭菌(121℃, 0.1MPa) 30min后自然冷却, 并充分振荡, 以恢复原有的溶解气体量。两藻种的实验均分两组进行, 分别加入NaNO3和NaH2PO4以设置氮磷的浓度, 氮组固定磷的浓度, 磷组固定氮的浓度, 每个浓度水平设置三个平行样。培养实验初始将生长良好的指数期藻液置于100mL离心管中, 于1700r/min的速度下离心10min, 去除上清液保留下层藻液, 以尽量减少藻液中残留的营养盐。中肋骨条藻和东海原甲藻的相应培养浓度如表 1, 其他的营养液均按照f/2配方加入(Ryther et al, 1962; Guillard, 1975)。培养瓶置于GXZ恒温培养箱, 设置中肋骨条藻培养温度为25℃, 东海原甲藻20℃, 光强3000-5000lx, 光暗比12︰12, 每天定时摇动三次, 保持气体的溶解量, 防止浮游藻聚集。每天同一时间, 用BD Accuri C6 Plus流式细胞仪测定浮游藻量, 指标下降时结束。
| 氮组磷酸盐浓度 | 磷组硝酸盐浓度 | |
| 中肋骨条藻: 氮组固定硝酸盐浓度为120μmol/L; 磷组固定磷酸盐浓度为3.0μmol/L |
0 | 0 |
| 0.5 | 20 | |
| 1.0 | 40 | |
| 1.5 | 60 | |
| 2.0 | 80 | |
| 3.0 | 120 | |
| 4.0 | 160 | |
| 东海原甲藻: 氮组固定硝酸盐浓度为120μmol/L; 磷组固定磷酸盐浓度为2.0μmol/L |
0 | 0 |
| 0.5 | 20 | |
| 1.0 | 40 | |
| 1.5 | 60 | |
| 2.0 | 80 | |
| 3.0 | 120 | |
| 4.0 | 160 |
利用插值法, 得出表层和底层的盐度空间格局(图 3)。从盐度分布来看, 与底层相比, 表层冲淡水向外海扩展较远, 且不同年份盐度梯度变化差异较大; 而底层盐度在不同年份的空间格局非常相似, 河口拦门沙附近梯度变化最剧烈。根据沉积物特点来看, 口门拦门沙附近10m等深线以内的区域, 冲积滩特征明显, 主要为粗颗粒沙, 而口外区10-60m等深线的区域, 沉积物较细(陈沈良等, 2009)。而沉积物分布特征可以反映水动力条件、地貌类型以及泥沙来源等因素的影响, 所以是长江口生态分区的重要参考因素(陈沈良等, 2009)。在分析以上因素的基础上, 同时考虑行政管理的便利性, 前哨海岸、佘山和鸡骨礁为几个标志性位置点, 所以将其的弧形连线作为分区的界限。
根据以上结果, 将长江口分为两个区域: Ⅰ区(口门区)和Ⅱ区(口外区)(图 4)。Ⅰ区西起徐六泾, 此为冬季盐水入侵锋所及位置, 河口分汊的重要节点; 东至江苏启东前哨海岸、佘山(31°25.3′N, 122°14.4′E)和鸡骨礁(31°12′N, 122°19′E)大致呈弧形的连接分界线。Ⅰ区属于咸淡水过渡区, 盐度为7.78±7.88(平均值±标准差), 90%的采样点盐度值在20以内, 此区域分布于长江口最大浑浊带, 透明度较低, 强烈受淡水径流的影响, 陆源污染物随长江流域直接排入此区域, 营养物污染严重。Ⅱ区西起分界线, 东至123°E河口锋前沿附近, 盐度为28.76±4.44, 90%的采样点盐度值在33以内, 盐度变化相对较小, 此区域表层会受冲淡水的影响小, 而底层主要为盐度大于30的咸水, 受海水冲刷和自净作用, 营养物浓度较低。
 |
| 图 4 长江口分区结果 Fig. 4 Preliminary results of classifications on Changjiang River estuary |
首先对两个分区水体表层的水温、pH、DO、COD、TOC和TN等进行方差齐性检验, 筛选出方差齐性的数据, 然后进行单因素方差分析, 各指标具体统计结果详见表 2。结果显示, 两个分区之间各个参数的差异显著性水平均小于0.01, 差异性显著, 说明该分区基本合理。
| 指标 | 分区 | 平均值 | 标准差 | 标准误差 | 平均值的95% | 置信区间 | 最小值 | 最大值 | 显著性 |
| COD (mg/L) | Ⅰ区 | 1.878 | 0.4927 | 0.0626 | 1.753 | 2.004 | 1.090 | 3.160 | P < 0.01 |
| Ⅱ区 | 1.176 | 0.4126 | 0.0708 | 1.032 | 1.320 | 0.430 | 2.710 | ||
| pH | Ⅰ区 | 7.914 | 0.1140 | 0.0145 | 7.885 | 7.943 | 7.630 | 8.060 | P < 0.01 |
| Ⅱ区 | 8.101 | 0.1053 | 0.0181 | 8.064 | 8.137 | 7.910 | 8.330 | ||
| 水温(℃) | Ⅰ区 | 29.309 | 1.1666 | 0.1482 | 29.012 | 29.605 | 26.420 | 31.490 | P < 0.01 |
| Ⅱ区 | 27.331 | 0.8766 | 0.1503 | 27.025 | 27.637 | 26.220 | 28.850 | ||
| TOC(mg/L) | Ⅰ区 | 1.504 | 0.2309 | 0.0293 | 1.446 | 1.563 | 1.130 | 2.030 | P < 0.01 |
| Ⅱ区 | 1.292 | 0.2862 | 0.0491 | 1.192 | 1.392 | 0.710 | 1.960 | ||
| DO(mg/L) | Ⅰ区 | 8.268 | 0.6254 | 0.0794 | 8.109 | 8.427 | 7.230 | 9.880 | P < 0.01 |
| Ⅱ区 | 7.436 | 0.4642 | 0.0796 | 7.274 | 7.598 | 6.350 | 8.320 | ||
| TN(mg/L) | Ⅰ区 | 1.427 | 0.3705 | 0.0470 | 1.333 | 1.521 | 0.613 | 2.580 | P < 0.01 |
| Ⅱ区 | 0.504 | 0.3546 | 0.0608 | 0.380 | 0.628 | 0.146 | 2.130 | ||
| 注: Ⅰ区62站, Ⅱ区34站 | |||||||||
通过主成分分析, 得出指标间的相关系数矩阵(表 3)、主成分特征值及累积贡献率(表 4)。根据主成分分析的基本要求, 发现前三个主成分的特征值大于1, 且累积贡献率达到了74.703%, 所以提取三个主成分, 从而达到数据降维的目的。为了对各主成分相关指标进行分析, 将每个主成分与各指标相关系数进行旋转, 得出旋转后的成分与原始指标变量的因子载荷矩阵(表 5)。
| 指标 | pH | DO | SS | SRP | DIN | chl a | S |
| pH | 1 | 0.288** | -0.028 | -0.342** | -0.340** | 0.205** | 0.313** |
| DO | 0.288** | 1 | 0.138** | 0.181** | 0.331** | -0.026 | -0.297** |
| SS | -0.028 | 0.138** | 1 | 0.238** | 0.198** | -0.103** | -0.105** |
| SRP | -0.342** | 0.181** | 0.238** | 1 | 0.641** | -0.265** | -0.621** |
| DIN | -0.340** | 0.331** | 0.198** | 0.641** | 1 | -0.148** | -0.903** |
| chl a | 0.205** | -0.026 | -0.103** | -0.265** | -0.148** | 1 | 0.064 |
| S | 0.313** | -0.297** | -0.105** | -0.621** | -0.903** | 0.064 | 1 |
| 注: **表示在P < 0.01级别(双尾), 相关性显著 | |||||||
| 主成分 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| 特征值 | 2.845 | 1.347 | 1.038 | 0.853 | 0.451 | 0.379 | 0.087 |
| 方差百分比 | 40.637 | 19.240 | 14.825 | 12.190 | 6.443 | 5.414 | 1.25 |
| 累积贡献率(%) | 40.637 | 59.877 | 74.703 | 86.893 | 93.336 | 98.750 | 100 |
| 指标 | S | DIN | SRP | DO | pH | chl a | SS |
| 主成分1 | -0.948 | 0.943 | 0.758 | 0.347 | -0.43 | -0.074 | 0.102 |
| 主成分2 | -0.046 | 0.058 | -0.090 | 0.792 | 0.755 | 0.252 | 0.260 |
| 主成分3 | 0.038 | 0.093 | 0.331 | 0.093 | -0.119 | -0.742 | 0.715 |
其中第一个主成分的特征值和贡献率最大, 其包含的信息最多。从因子载荷矩阵来看, 与第一主成分关系密切的监测指标有三个, 分别是DIN、SRP和盐度。它反映了两方面的信息, 一方面是营养指标, 河口区营养盐含量是富营养化评价的重要指标; 另一方面是盐度, 盐度是河口生态分区的关键指标之一, 它在一定程度上反映了河口区域的本底状况, 是生物地球化学过程的主要控制参数(刘录三等, 2011b)。所以将第一主成分定义为营养物基准分区指标和营养指标。
对长江口各指标因子间进行两两相关性分析, 并对其进行双尾显著性检验(表 3)。相关关系表明, DIN、SRP、DO和SS与盐度呈显著负相关, 且与DO和SS相比, DIN和SRP与盐度的相关性较强, 而pH与盐度呈显著正相关, chl a与盐度相关性不显著。从线性回归图中可以看出, 随着盐度的增加, DIN和SRP浓度均逐渐降低, 与SRP相比, DIN表现出更好的保守性, 在该地区的其他研究中也观察到了类似的相关性(张平, 2001; 辛明, 2014)(图 5)。
 |
| 图 5 长江口营养盐浓度与盐度的相关关系 Fig. 5 Relationship between nutrient concentrations and salinity in Changjiang River estuary |
本研究中生境状况的判断运用生态系统健康评价的结果, 健康及以上等级为完好, 亚健康等级为部分退化, 不健康及以下等级为严重退化, 研究表明, 长江口生态系统处于亚健康状态, 即生境部分退化(周晓蔚等, 2011; 中华人民共和国生态环境部, 2019)。参照点的选择参考了生态环境脆弱性评价结果和历史赤潮记录。口门区接近海岸线, 工业、渔业和旅游业等产业发达, 较20世纪90年代初期, 区域内生物多样性明显降低, 浮游生物物种密度年际间变幅较大, 生态系统结构不稳定, 且历史记录中佘山附近的水域发生过赤潮事件, 参照点难寻, 所以选取所有监测数据营养指标频数分布曲线的下5%对应值作为口门区的营养物基准值(周晓蔚等, 2011; 何彦龙等, 2019)。口外区虽离海岸较远, 但也受到来自长江径流污染物的影响, 营养物结构和水平发生了显著变化, 此外还受水体层化、上升流加强、黑潮入侵等多种因素影响, 区域内有大面积的赤潮现象, 选取没有赤潮记录以及处于微度和轻度脆弱等级的点位作为参照点, 将参照点营养指标频数分布曲线的上25%对应值作为口外区营养物基准值(刘录三等, 2011a; 于仁成等, 2017)。利用SPSS 24进行频数分析拟合, 绘制长江口DIN和SRP的频数分布统计表(表 6)和频数分布直方图(图 6, 图 7), 得到口门区DIN和SRP基准值分别为0.614和0.029mg/L; 口外区DIN和SRP基准值分别为0.300和0.022mg/L。
| 类别 | 口门区(所有观测点) | 口外区(参照点) | ||||
| DIN | SRP | DIN | SRP | |||
| 样本量 | 594 | 597 | 195 | 186 | ||
| 平均值(mg/L) | 1.448 | 0.048 | 0.251 | 0.018 | ||
| 标准差 | 0.482 | 0.016 | 0.199 | 0.009 | ||
| 最小值(mg/L) | 0.189 | 0.003 | 0.024 | 0 | ||
| 最大值(mg/L) | 3.208 | 0.260 | 1.269 | 0.059 | ||
| 百分位数统计值(mg/L) | 5% | 0.614 | 0.029 | 0.078 | 0.004 | |
| 25% | 1.132 | 0.039 | 0.127 | 0.012 | ||
| 50% | 1.458 | 0.047 | 0.187 | 0.017 | ||
| 75% | 1.739 | 0.055 | 0.300 | 0.022 | ||
| 95% | 2.146 | 0.071 | 0.696 | 0.038 | ||
 |
| 图 6 口门区DIN和SRP频数分布直方图 Fig. 6 Histogram of frequency distribution of DIN and SRP in the mouth area 注: cDIN表示可溶性无机氮浓度, cSRP表示可溶性活性磷酸盐的浓度 |
 |
| 图 7 口外区DIN和SRP频数分布直方图 Fig. 7 Histogram of frequency distribution of DIN and SRP in the outside-mouth area 注: lncDIN表示可溶性无机氮浓度的对数值, lncSRP表示可溶性活性磷酸盐浓度的对数值 |
运用CART法建立压力-响应模型, 将分类回归树的根节点营养指标浓度值作为基准值。SRP和DIN浓度分别是影响区域内chl a和DO浓度变化的首要原因变量, SRP浓度值0.05705和0.00845mg/L分别为图 8口门区和口外区分类回归树的根节点, DIN浓度值0.6273和0.2951mg/L分别为图 9口门区和口外区分类回归树的根节点, 保留三位小数, 得到口门区DIN和SRP的基准值分别为0.627和0.057mg/L, 口外区DIN和SRP基准值分别为0.295和0.008mg/L。
 |
| 图 8 口门区(a)和口外区(b) DIN和SRP浓度值对chl a浓度值划分的分类回归树模型 Fig. 8 The CART of values of chl a divided by DIN and SRP concentrations in the mouth area (a) and outside-mouth area (b) 注:箱图代表每个终端节点chl a的浓度分布, 按分类数据中平均chl a浓度从小到大排列 |
 |
| 图 9 口门区(a)和口外区(b)DIN和SRP浓度值对DO浓度值划分的分类回归树模型 Fig. 9 The CART of values of DO divided by DIN and SRP concentrations in the mouth area (a) and outside-mouth area (b) 注:箱图代表每个终端节点DO的浓度分布, 按分类数据中平均DO浓度从小到大排列 |
为了尽量实现对河口生态系统的保护, 选取频数分布法和压力-响应模型结果的低值作为基准推荐值, 最终得出口门区DIN和SRP的基准推荐值分别为0.614和0.029 mg/L, 口外区DIN和SRP的基准推荐值分别为0.295和0.008mg/L。与长江口的其他研究相比, 本研究口门区和口外区的基准推荐值均小于其结果, 这一方面是所选用数据不同的原因, 另一方面可能是由于频数分布法的运用有差别, 这就需要建立更加完整的河口基准数据库, 探索系统的分类分区体系, 完善营养物基准推导方法(表 7)(Yang et al, 2019)。
| 地理位置 | 推导方法 | 口门区(mg/L) | 口外区(mg/L) | 参考文献 | |||
| DIN限值 | SRP限值 | DIN限值 | SRP限值 | ||||
| 优(第一类) | 优(第一类) | ||||||
| 英国水框架指令健康水质等级 | 参照状态法 | 0.28 | NA | 0.18 | NA | (European Commission, 2003) | |
| 九龙江口 | 参照状态法 | 0.341-0.702 | 0.016 | 0.231 | 0.016 | (Liu et al, 2018) | |
| 模型法 | 0.294-0.899 | 0.028 | 0.200 | 0.028 | |||
| 频数分布法 | 0.297-0.964 | 0.024-0.037 | 0.272 | 0.028 | |||
| 基准推荐值 | 0.294-0.896 | 0.024-0.028 | 0.196 | 0.028 | |||
| 长江口水域 | 频数分布法 | 1.05-1.24 | 0.030-0.036 | 1.05-1.24 | 0.030-0.036 | (Yang et al, 2019) | |
| 长江口水域 | 频数分布法 | 0.614 | 0.029 | 0.300 | 0.022 | 本研究 | |
| 压力-响应模型 | 0.627 | 0.057 | 0.295 | 0.008 | |||
| 基准推荐值 | 0.614 | 0.029 | 0.295 | 0.008 | |||
| 海水水质标准(GB3097-1997) | NA | NA | NA | 0.20 | 0.015 | (国家环境保护局, 2004) | |
| 注: NA:数据缺失 | |||||||
根据公式(1)曲线拟合, 得到两种赤潮敏感藻的生长曲线图(图 10, 11)。对于中肋骨条藻, 当DIN浓度固定为120μmol/L时, 随着SRP浓度的增大, 浮游藻的最终生物量逐渐增加, 但增加变缓, 固定SRP为3.0μmol/L时, 随着DIN浓度的增大浮游藻的最终生物量也呈现同样的规律。而对于东海原甲藻, 当DIN固定时为120μmol/L时, 随着SRP浓度的增大, 浮游藻的最终生物量逐渐增加, 而当固定SRP为2.0μmol/L时, DIN浓度在0-80μmol/L的范围内变化时, 浮游藻的最终生物量逐渐增加, 但超过80μmol/L时, 最终生物量开始下降, 这可能是由于东海原甲藻对磷酸盐的吸收效率高, 氮磷比超过40则生长受限(张璇, 2012)。
 |
| 图 10 中肋骨条藻氮组(a)和磷组(b)的生长曲线拟合图 Fig. 10 Growth curves of Skeletonema costatum of nitrogen group (a) and phosphorus group (b) |
 |
| 图 11 东海原甲藻氮组(a)和磷组(b)的生长曲线拟合图 Fig. 11 Growth curves of Prorocentrum donghaiensis of nitrogen group (a) and phosphorus group (b) 注:误差棒表示三个平行样的标准差 |
运用1.5.1小节中生态响应值的计算方法, 得到中肋骨条藻对DIN和SRP的生态响应值分别为0.535和0.025mg/L, 东海原甲藻对DIN和SRP的生态响应值分别为0.254和0.043mg/L。分别将中肋骨条藻和东海原甲藻的响应值作为Ⅰ区和Ⅱ区的生态响应值, 得到长江口Ⅰ区DIN和SRP的生态响应值分别为0.535和0.025mg/L, Ⅱ区DIN和SRP的生态响应值分别为0.254和0.043mg/L (表 8)。
| 浮游藻 | DIN生态响应值 | 95%置信区间CL | SRP生态响应值 | 95%置信区间CL | ||
| 下限LCL | 上限UCL | 下限LCL | 上限UCL | |||
| 中肋骨条藻 | 0.535 | 0.499 | 0.585 | 0.025 | 0.022 | 0.027 |
| 东海原甲藻 | 0.254 | 0.241 | 0.268 | 0.043 | 0.029 | 0.052 |
本研究对长江口的生态分区结果符合连续性和便利性原则(Perillo, 1995)。口门区淡水和海水的混合过程非常强烈, 当地的水产养殖业集中在这一区域, 且来自地表径流的污染物在此处排放入海, 所以, 对此区域水文和营养物污染等方面的评估和管理非常关键(林晓娟, 2018)。口外区主要以海水为主, 虽然营养物浓度较低, 但发生赤潮的风险仍然较大, 所以对营养指标的总量控制也很重要(刘录三等, 2011a)。两个分区的动力学特征和地理特征有明显不同, 同时与水行政管理部门对河海的划界非常接近(张华明, 2009)。此外, 单因素方差分析的结果也表明, 两个分区在多项指标中都有显著性差异, 这与河口中的生物、化学和物理过程密切相关, 由此说明此分区的合理性(Telesh et al, 2010)。然而, 河口区域的水质和生物群落还会受河口的气候、水域特征(例如土地利用)和人类活动等多种因素的影响, 此研究初步实现了分区管理, 但对一些重点管控区域缺乏保护, 例如自然保护区、饮用水水源保护区、特别保护海岛、重要渔业海域等, 这就需要在此基础上根据其功能和海洋生态红线等因素, 通过进一步的生态分区或水体功能分级的标准制定来有效评估污染状况并实施污染控制, 实现对河口的管理和保护(Elliott et al, 2002; 席北斗等, 2013)。
本研究主要关注DIN和SRP, 而未对TN和TP做相关分析, 这是由我国水环境管理的实际情况决定的, 现行的《海水水质标准》(GB 3097-1997)中, 水质监测与评价中只要求DIN和SRP, 造成TN和TP数据缺失。本研究运用了近几年的监测数据对营养物基准值进行推导, 还可以参考历史数据, 将未被开发的状态作为参照状态。20世纪80年代长江口基本无富营养化现象, 1984-2015年间, 长江口及其邻近水域DIN和SRP浓度有线性上升趋势, 1988年是营养盐含量快速增长的起始年, 因此, 将1984-1987年营养盐要素的平均值作为参考值, DIN和SRP的平均浓度值分别为0.0705和0.0008mg/L (范海梅等, 2015, 2019)。可以发现, 本研究得到的基准推荐值远远大于此参考值, 说明近30年来长江口营养物的排放已严重超标, 要使水质恢复到接近未受干扰的状态需要大量的时间和努力, 如何把营养物浓度控制在一个恰当的范围是最关键的问题, 需既能保证初级生产量以及渔业的正常发展, 又不发生富营养化造成生态系统的失衡。另外一个值得关注的问题是营养盐结构, 本研究中得到口门区和口外区的基准推荐值氮磷比分别为46.82和29.65, 与Redfield比率16相比, 明显偏大, 特别是口门区, 将近是16的3倍(Redfield, 1960)。这与长江口的氮通量大有关, 区域内有较大程度的磷限制, 这与之前的研究一致(李磊等, 2010; 陈慧敏等, 2011)。
为了有效控制富营养化, 需要考虑河口生态系统(尤其是浮游藻)对营养物的敏感性, 而长江口区域中肋骨条藻和东海原甲藻是对营养物最敏感的赤潮藻类(张远等, 2006; 中华人民共和国自然资源部, 1989-2019)。本研究中, 生态响应值是指区域内赤潮优势藻对营养盐浓度产生大量繁殖响应的临界值, 通过浮游藻生物量随营养指标浓度变化的“S”型曲线拐点值得到, 以防止河口区浮游藻的大量暴发。然而, 虽然单物种藻类培养可以初步得出生态响应值, 但未考虑种间竞争、浮游动物等的影响, 所以需要进一步进行多种浮游藻的微宇宙群落试验, 以校验营养物基准值。
我国对长江口的研究相对完整, 本研究以长江口为例探讨了不同生态分区的营养指标基准值, 然而, 为了实现河口及近岸的环境管理, 我们必须对河口区域进行长期的监测和研究, 建立完整的河口基准数据库, 探索科学合理的河口分类和分区方法, 对多项营养物指标进行分析和推导, 以建立完整的营养物基准体系, 从而实现对赤潮等有害事件的防控, 保证河口生态系统的可持续性。
4 结论(1) 根据盐度分布、地形特点和行政管理的便利性, 将长江口分为两个生态区: Ⅰ区(口门区)和Ⅱ区(口外区), 分区检验发现两生态区各指标差异性显著, 分区基本合理。
(2) 基于主成分分析、相关性分析和线性回归分析, 得出DIN和SRP是制定长江口营养物基准的关键指标, 而盐度是其重要影响因素之一。DIN、SRP、DO和SS与盐度呈显著负相关, 与SRP相比, DIN表现出更好的保守性。
(3) 采用频数分布法和压力-响应模型法得到口门区DIN和SRP基准推荐值分别为0.614和0.029mg/L, 口外区基准推荐值分别为0.295和0.008mg/L。
(4) 通过中肋骨条藻和东海原甲藻室内培养实验, 得到口门区DIN和SRP的生态响应值分别为0.535和0.025mg/L, 口外区DIN和SRP的生态响应值分别为0.254和0.043mg/L。
于仁成, 张清春, 孔凡洲, 等. 2017. 长江口及其邻近海域有害藻华的发生情况、危害效应与演变趋势. 海洋与湖沼, 48(6): 1178-1186 |
王江涛, 曹婧. 2012. 长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 31(3): 310-315 |
中华人民共和国生态环境部, 2019.2018年中国海洋生态环境状况公报. http://hys.mee.gov.cn/dtxx/201905/P020190529532197736567.pdf
|
中华人民共和国自然资源部, 1989-2019.1989-2018年中国海洋灾害公报. http://www.mnr.gov.cn/sj/sjfw/hy/gbgg/zghyzhgb/
|
中国海湾志编纂委员会. 1998. 中国海湾志, 第十四分册(重要河口). 北京: 海洋出版社
|
石岩峻, 2004.赤潮藻对营养盐的吸收及生长和相关特性研究.北京: 北京化工大学博士学位论文, 20-21
|
刘录三, 李子成, 周娟, 等. 2011a. 长江口及其邻近海域赤潮时空分布研究. 环境科学, 32(9): 2497-2504 |
刘录三, 郑丙辉, 孟伟, 等. 2011b. 基于自然地理特征的长江口水域分区. 生态学报, 31(17): 5042-5054 |
苏纪兰. 2001. 中国近海的环流动力机制研究. 海洋学报, 23(4): 1-16 |
李磊, 沈新强. 2010. 春、夏季长江口海域营养盐的时空分布特征及营养结构分析. 生态环境学报, 19(12): 2941-2947 |
李薇, 2014.溶解氧水平对富营养化水体底泥氮磷转化影响的研究.南京: 南京理工大学硕士学位论文, 1-67
|
李俊龙, 郑丙辉, 刘永, 等. 2015. 中国河口富营养化对营养盐负荷的敏感性分类. 中国科学:地球科学, 45(4): 455-467 |
李照, 2018.长江口邻近海域浮游植物群落特征及其碳沉降研究.青岛: 中国科学院大学(中国科学院海洋研究所)博士学位论文, 1-127
|
何彦龙, 袁一鸣, 王腾, 等. 2019. 基于GIS的长江口海域生态系统脆弱性综合评价研究. 生态学报, 39(11): 3918-3925 |
辛明, 2014.长江口海域关键环境因子的长期变化及其生态效应.青岛: 中国海洋大学博士学位论文, 102-104
|
沈焕庭, 茅志昌, 朱建荣. 2003. 长江河口盐水入侵. 北京: 海洋出版社, 175
|
沈焕庭, 贺松林, 茅志昌, 等. 2001. 中国河口最大浑浊带刍议. 泥沙研究, (1): 23-29 |
张平, 2001.长江口营养盐结构变化研究.青岛: 中国科学院海洋研究所硕士学位论文, 1-69
|
张远, 郑丙辉, 富国, 等. 2006. 河道型水库基于敏感性分区的营养状态标准与评价方法研究. 环境科学学报, 26(6): 1016-1021 |
张璇, 2012.长江口及邻近海域营养盐的历史演变及其在赤潮中的作用研究.青岛: 中国海洋大学硕士学位论文, 1-76
|
张华明. 2009. 长江口管理界线的现状和分析. 海洋学研究, 27(S1): 10-16 |
张清春, 2011.东海大规模甲藻赤潮种吞噬特性及其在赤潮形成中的作用初探.青岛: 中国科学院海洋研究所博士学位论文, 1-119
|
陆健健. 2003. 河口生态学. 北京: 海洋出版社, 318
|
陈沈良, 严肃庄, 李玉中. 2009. 长江口及其邻近海域表层沉积物分布特征. 长江流域资源与环境, 18(2): 152-156 |
陈慧敏, 孙承兴, 仵彦卿. 2011. 近23a来长江口及其邻近海域营养盐结构的变化趋势和影响因素分析. 海洋环境科学, 30(4): 551-553, 563 |
陈翰林, 吕颂辉, 张传松, 等. 2006. 2004年东海原甲藻赤潮爆发的现场调查和分析. 生态科学, 25(3): 226-230 |
范海梅, 高秉博, 余江, 等. 2015. 上海海域营养盐趋势与长江排海量相关性研究. 上海环境科学, 34(1): 1-5, 25 |
范海梅, 蒋晓山, 纪焕红, 等. 2019. 长江口及其邻近海域生态环境综合评价. 生态学报, 39(13): 4660-4675 |
林晓娟, 2018.基于模糊综合评价法的长江口海水富营养化评价方法研究.厦门: 厦门大学硕士学位论文, 58-59
|
国家环境保护局. 2004. GB 3097-1997海水水质标准. 北京: 环境科学出版社
|
周晓蔚, 王丽萍, 郑丙辉. 2011. 长江口及毗邻海域生态系统健康评价研究. 水利学报, 42(10): 1201-1208 |
郑丙辉, 周娟, 刘录三, 等. 2013. 长江口及邻近海域富营养化指标原因变量参照状态的确定. 生态学报, 33(9): 2780-2789 |
孟伟, 张远, 郑丙辉. 2006. 水环境质量基准、标准与流域水污染物总量控制策略. 环境科学研究, 19(3): 1-6 |
席北斗, 陈艳卿, 苏婧, 等. 2013. 湖泊营养物标准方法学及案例研究. 北京: 科学出版社, 264
|
黄备, 邵君波, 魏娜, 等. 2014. 2014年春季东海大面积甲藻赤潮的生态特征. 生态环境学报, 23(9): 1457-1462 |
霍守亮, 马春子, 席北斗, 等. 2017. 湖泊营养物基准研究进展. 环境工程技术学报, 7(2): 125-133 |
戴鑫烽, 陆斗定, 夏平, 等. 2014. 2010-2011年东海藻华高发区水体层化对东海原甲藻(Prorocentrum donghaiense Lu)藻华的影响. 海洋与湖沼, 45(2): 217-224 |
De'ath G, Fabricius K E, 2000. Classification and regression trees:a powerful yet simple technique for ecological data analysis. Ecology, 81(11): 3178-3192 DOI:10.1890/0012-9658(2000)081[3178:CARTAP]2.0.CO;2 |
Elliott M, McLusky D S, 2002. The Need for Definitions in Understanding Estuaries. Estuarine, Coastal and Shelf Science, 55(6): 815-827 DOI:10.1006/ecss.2002.1031 |
European Commission, 2003. Common implementation strategy for the water framework directive. Transitional and coastal waters: typology, reference conditions and classification systems. Luxembourg: Office for Official Publications of the European Communities
|
Guillard R R L, 1975. Culture of phytoplankton for feeding marine invertebrates. In: Smith W L, Chanley M H eds. Culture of Marine Invertebrate Animals. Boston: Springer, 26-60
|
Liu B L, Cao W Z, Huang Z et al, 2018. Developing nutrient criteria for the Jiulong River Estuary, Southeast China. Acta Oceanologica Sinica, 37(2): 1-13 DOI:10.1007/s13131-017-1121-0 |
Perillo G M E, 1995. Definitions and geomorphologic classifications of estuaries. Developments in Sedimentology, 53: 17-47 DOI:10.1016/S0070-4571(05)80022-6 |
Redfield A C, 1960. The biological control of chemical factors in the environment. Science Progress, 11: 150-170 DOI:10.1086/646891 |
Ryther J H, Guillard R R L, 1962. Studies of marine planktonic diatoms:Ⅱ. Use of Cyclotella nana Hustedt for assays of vitamin B12 in sea water. Canadian Journal of Microbiology, 8(4): 437-445 DOI:10.1139/m62-057 |
Telesh I V, Khlebovich V V, 2010. Principal processes within the estuarine salinity gradient:A review. Marine Pollution Bulletin, 61(4-6): 149-155 DOI:10.1016/j.marpolbul.2010.02.008 |
USEPA, 1998. EPA-822-R-98-002 National strategy for the development of regional nutrient criteria. Washington DC: USEPA
|
USEPA, 2000a. EPA-822-B-00-024 Estuarine and coastal marine waters: bioassessment and biocriteria technical guidance. Washington DC: USEPA
|
USEPA, 2000b. EPA-822-B-00-001 Nutrient criteria technical guidance manual lakes and reservoirs. Washington DC: USEPA
|
USEPA, 2001. EPA-822-B-01-003 Nutrient criteria technical guidance manual. Estuarine and coastal marine waters. Washington DC: USEPA
|
USEPA, 2008. EPA-822-B-08-001 Nutrient criteria technical guidance manual: wetlands. Washington DC: USEPA
|
USEPA, 2010. EPA-820-S-10-001 Using stressor-response relationships to derive numeric nutrient criteria. Washington, DC: USEPA
|
Wei H, He Y C, Li Q J et al, 2007. Summer hypoxia adjacent to the Changjiang Estuary. Journal of Marine Systems, 67(3-4): 292-303 DOI:10.1016/j.jmarsys.2006.04.014 |
Wu F C, Meng W, Zhao X L et al, 2010. China embarking on development of its own national water quality criteria system. Environmental Science & Technology, 44(21): 7992-7993 DOI:10.1021/es1029365 |
Yang F X, Mi T Z, Chen H T et al, 2019. Developing numeric nutrient criteria for the Yangtze River Estuary and adjacent waters in China. Journal of Hydrology, 579: 124188 DOI:10.1016/j.jhydrol.2019.124188 |