 2020, Vol. 51
2020, Vol. 51
黑潮延伸体区域叶绿素季节变化以及中尺度涡对其影响机制研究

http://dx.doi.org/10.11693/hyhz20191100234
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 胡松, 李敏华, 刘必林, 周相乾, 余为. 2020.
- HU Song, LI Min-Hua, LIU Bi-Lin, ZHOU Xiang-Qian, YU Wei. 2020.
- 黑潮延伸体区域叶绿素季节变化以及中尺度涡对其影响机制研究
- SEASONAL CHARACTERISTICS OF CHLOROPHYLL A CONCENTRATION IN KUROSHIO EXTENSION AND INFLUENCES OF MESOSCALE EDDIES
- 海洋与湖沼, 51(6): 1370-1378
- Oceanologia et Limnologia Sinica, 51(6): 1370-1378.
- http://dx.doi.org/10.11693/hyhz20191100234
文章历史
-
收稿日期:2019-11-28
收修改稿日期:2020-03-15
引用本文
胡松, 李敏华, 刘必林, 周相乾, 余为. 2020. 黑潮延伸体区域叶绿素季节变化以及中尺度涡对其影响机制研究[J]. 海洋与湖沼, 51(6): 1370-1378

HU Song, LI Min-Hua, LIU Bi-Lin, ZHOU Xiang-Qian, YU Wei. 2020. SEASONAL CHARACTERISTICS OF CHLOROPHYLL A CONCENTRATION IN KUROSHIO EXTENSION AND INFLUENCES OF MESOSCALE EDDIES[J]. Oceanologia et Limnologia Sinica, 51(6): 1370-1378.
黑潮延伸体区域叶绿素季节变化以及中尺度涡对其影响机制研究
1. 上海海洋大学海洋科学学院 上海 201306;
2. 国家远洋渔业工程技术研究中心 上海 201306;
3. 上海河口海洋测绘工程技术研究中心 上海 201306
2. 国家远洋渔业工程技术研究中心 上海 201306;
3. 上海河口海洋测绘工程技术研究中心 上海 201306
摘要:中尺度涡是影响西北太黑潮延伸体南北海域叶绿素分布的重要因子。通过对2012—2018年NOAA遥感水色卫星数据和法国COPERNICUS流场资料的统计分析,发现中尺度涡对表层叶绿素浓度的影响具有明显的季节性特征。在春季有17%的气旋涡导致中心叶绿素高值结构且多数向西传播,而其余季节对叶绿素浓度影响较弱。反气旋涡导致环状叶绿素高值结构,在春、秋两季常有发生,在冬夏出现频次少。中尺度涡影响叶绿素能力的季节差异可能与营养盐背景场、光限制、等密线等季节性变化有关,不能只考虑风和流场的垂向输运机制。叶绿素浓度在一定程度上反映浮游植物生物量,对黑潮延伸体海域有重要生态意义。因此,该区域服务渔业的中尺度涡研究应该考虑这种季节特征。
关键词:黑潮延伸体 中尺度涡 叶绿素 生态系统动力学
SEASONAL CHARACTERISTICS OF CHLOROPHYLL A CONCENTRATION IN KUROSHIO EXTENSION AND INFLUENCES OF MESOSCALE EDDIES
1. College of Marin Sciences, Shanghai Ocean University, Shanghai 201306, China;
2. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
3. Shanghai Engineering Research Center for Estuarine and Oceanographic Mapping, Shanghai 201306, China
2. National Engineering Research Center for Oceanic Fisheries, Shanghai 201306, China;
3. Shanghai Engineering Research Center for Estuarine and Oceanographic Mapping, Shanghai 201306, China
Abstract: The mesoscale eddy is one of the most important factors influencing the distribution of chlorophyll a (chl a) in the Kuroshio extension of the Northwest Pacific. Based on statistical analysis on the weekly remote-sensing ocean surface chl a concentration data from NOAA,and geostrophic current data from Copernicus for the period of 2012—2018, we found that the influence of mesoscale eddy on the surface chl a concentration showed significant seasonal characteristics. About 17% of the cyclonic eddies in spring had high chl a concentration in the center and propagate westward. However,the cyclonic eddies in other seasons have a little effect on chl a concentration. The anticyclonic eddies generate a high chl a ring in both spring and autumn,with a little effect on chl a concentration in summer and winter. The seasonal characteristics of chl a in mesoscale eddies may relate to seasonal nutrient-limitation,light-limitation or the change of pycnocline,indicating that wind- and flow-induced vertical nutrient transport is not the only factor to be considered. The chl a concentration can represent phytoplankton biomass to some extents,which is important for the regional ecosystem. Thus,the effect of seasonal feature of mesoscale eddy should be considered in the studies for fisheries.
Key words:
Kuroshio extension mesoscale eddy chlorophyll a ecosystem dynamics
中尺度涡是大洋环流运动中的主要组成部分(Chelton et al, 2011;汤博等, 2019), 其直径通常在数百公里之内, 并且能够维持数周乃至数月。随着海洋学动力理论的发展以及观测手段日益丰富, 发现海洋中的大部分动能并非是稳定环流而是普遍存在的中尺度涡, 尤其在永久温跃层以上的海域(The MODE Group, 1978)。富有活力的西边界流例如黑潮是西北太平洋涡动能聚集区, 黑潮的行进轨迹经常摆动, 在黑潮延伸体北部易产生反气旋涡, 在南部易产生气旋涡(Olson, 1991), 这些涡生成后大部分向西传播。
黑潮暖流与亲潮寒流交汇处是重要的经济渔场。黑潮的物理环境和输运机制, 结合生物-化学过程, 是理解该区域海洋生态系统的基础, 并可能对柔鱼、秋刀鱼、沙丁鱼等中上层海洋经济种类的渔场有重要影响(Chen et al, 2011)。以高度洄游的头足类柔鱼为例(图 1), 其产卵场位于黑潮东南侧, 而成年柔鱼广泛分布在黑潮东北侧的渔场(余为等, 2013;刘金立等, 2014; Yu et al, 2015;余为, 2016)。另外柔鱼生命周期较短, 种群资源量基本取决于补充量, 因此黑潮与周围中尺度涡对柔鱼在不同生活史阶段的生存与迁移均存在一定影响。中尺度涡, 尤其是气旋涡, 可能通过水体输运将分布在黑潮南部的幼年柔鱼向西输送, 然后携带着这部分生物在靠近西边界处重新汇入黑潮, 从而对该区域的生态系统起到一定作用。中尺度涡对于热量、质量、营养盐上涌等海洋要素的物理输运影响是近年来的研究热点, 但是中尺度涡输运过程对于生物种群的影响在该海区目前还缺乏深入研究。
 |
| 图 1 黑潮延伸体位置和涡旋分布以及柔鱼分布情况示意图 Fig. 1 Schematic diagram of the location of Kuroshio extension, eddy, and the squid spawning and fishing grounds |
在中尺度涡运动过程中, 中心或周边经常出现浮游植物生长-爆发-消亡-沉降过程, 支撑起远洋食物链底层, 形成一个中尺度涡生态系统。国内外对中尺度涡和叶绿素(或浮游植物)的关系进行了广泛研究, 主要聚焦在中尺度涡对营养盐供给的动力机制及其对叶绿素浓度的影响研究等(Oschlies et al, 1998; Gruber et al, 2011; Gaube et al, 2014; Wang et al, 2016; Xu et al, 2019)。然而, 这些研究往往是对某个区域多个涡旋的全年统计来理解生态响应, 较少区分各个季节的情况。中尺度涡的空间结构处于中尺度范围, 其时间结构也要对应中尺度动力学, 浮游植物暴发时间尺度更短, 要厘清中尺度涡和叶绿素的关系应当考虑季节内的时间尺度。浮游植物暴发不仅与营养盐上涌有关, 还与光限制、混合层厚度、浮游动物摄食等季节性因子有重要关联。因此, 本文聚焦黑潮延伸体南北海域, 按照不同季节分析这些中尺度涡和叶绿素的对应关系, 并探讨这些关系可能对渔业资源带来的影响。
1 材料和方法 1.1 数据来源叶绿素多年气候态月平均数据来自COPERNI CUS海洋环境监测服务中心提供的产品(http://marine.copernicus.eu/), 是1998-2018年数据求平均所获得, 空间分辨率为4km。用于计算多年平均涡动能的海表面地转异常流速数据同样来自COPERNICUS提供的多颗卫星数据融合产品, 空间分辨率为0.25°×0.25°, 时间间隔为1周, 涡动能计算公式为
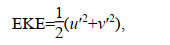 (1)
(1)其中, u′和v′为地转异常流速。短波辐射、2m气温、10m风速的气候态(1981-2010年)月平均数据来自NOAA提供的NCEP/NCAR再分析数据集(Kalnay et al, 1996), 混合层气候态月平均数据来自NOAA提供的MIMOC数据集(Schmidtko et al, 2013)。
用于分析季节内中尺度涡叶绿素变化的数据来自美国NOAA网站水色卫星产品的叶绿素α浓度值(Wang et al, 2016), 时间跨度为2012-2018年, 空间分辨率为4km, 时间分辨率为每周平均。用于统计2012-2018年中尺度涡数量的数据同样来自上述的COPERNICUS所提供的多颗卫星数据融合产品, 空间分辨率为0.25°×0.25°, 时间间隔为1周。
1.2 数据分析方法黑潮延伸体流轴南、北两侧叶绿素浓度差异直接影响中尺度涡内的表层叶绿素浓度和分布。利用多年月平均气候态数据计算得到目标区域叶绿素浓度气候分布(图 2)。为体现出延伸体南、北两侧浓度的季节变化, 选取了涡旋生成较多区域内一条经向断面(146°E)上的五个点, 计算并绘制了每个点的多年月平均叶绿素浓度(图 3)。
 |
| 图 2 1998-2018年目标区域海表叶绿素浓度气候态分布 Fig. 2 Distribution of climatological mean chlorophyll a concentration in the study area 注: A、B、C、D、E为146°E经向断面5个站位 |
采用董昌明等(2017)提出的基于海表流场的涡旋探测方法, 满足四个约束条件即被定义为涡旋中心。在识别涡旋中心后, 结合黑潮延伸体区域叶绿素分布情况做出如下处理: (1)将叶绿素周平均数据与海表面流场数据绘制出2012-2018年周平均海表流场与叶绿素叠加分布图, 总计336周; (2)将涡旋范围内叶绿素有效像素点占涡旋范围总像素点数30%以上的涡作为有效涡旋, 用以仔细分析涡旋与表层叶绿素相互影响, 共计1055个; (3)逐一检查数据后, 将涡旋范围内的典型海表叶绿素浓度分布分为涡旋中心显著和环状结构显著两种, 由于黑潮延伸体南、北侧叶绿素多年平均浓度的差异, 对于有效气旋涡, 涡旋中心叶绿素浓度高于涡旋边界处0.2mg/m3时定义为涡中心叶绿素浓度显著, 而有效反气旋涡旋中心浓度低于涡旋边界处0.3mg/m3时定义为叶绿素环状结构显著。根据上述定义, 对全部336周的卫星叶绿素和流场叠加图片的有效涡旋进行了季节统计。
2 数据分析结果 2.1 目标区域叶绿素分布特征目标区域内叶绿素浓度分布总体呈现出沿纬向呈带状分布(图 2), 并且存在北高南低、西高东低、近岸高远海低的特点。涡旋生成数量较多的区域(142°E以东), 叶绿素浓度从北至南逐渐降低。在此区域内, 37°N以北叶绿素浓度平均值为0.29mg/m3。在黑潮延伸体33°-37°N的涡旋活跃范围内, 海表叶绿素浓度平均值为0.21mg/m3, 略低于北部区域, 而在33°N以南区域, 叶绿素浓度平均值仅为0.11mg/m3。
黑潮延伸体主轴范围内, 叶绿素分布受海流影响呈现明显的蛇形状分布。135°E/32°N附近叶绿素带状结构向北凸起, 而在伊豆群岛附近受地形影响叶绿素舌向南弯曲, 随后在142°E/35°N附近随着黑潮延伸体主轴向东延伸。
与叶绿素浓度分布相对应的是, 表征涡旋活动剧烈程度的涡动能呈现出明显的纬向高值带状分布(图 3)。黑潮延伸体将此区域内叶绿素浓度分隔成明显的北侧高值区与南侧低值区。黑潮主轴以及黑潮延伸体流轴涡动能达到峰值, 延伸体南北两侧涡旋活动剧烈, 北侧反气旋式涡旋位于叶绿素浓度较高的环境中, 反之, 南侧气旋式涡旋多位于叶绿素浓度较低的环境。
 |
| 图 3 1998-2018年黑潮延伸体区域平均涡动能 Fig. 3 Eddy kinetic energy in Kuroshio extension region averaged from 1998 to 2018 |
目标区域内叶绿素浓度存在明显的季节性变化。图 4为选取的经向断面上5个点的叶绿素多年月平均浓度值, 最北端A点叶绿素浓度最高, 5月达到最大值0.97mg/m3, 8月达到最小值0.21mg/m3。E点叶绿素浓度最低, 4月达到最大值0.24mg/m3, 9月达到最低值0.06mg/m3。总体上看, 北部叶绿素浓度波动较大, 差值最大达0.76mg/m3, 向南波动逐渐减小, 最南端浓度差异值最大仅为0.18mg/m3。另外, 各点在一年内均有两次峰值点, A点在5月达到第一次峰值, 相比其余各点滞后一个月, 而A点在10月达到第二次峰值, 相比其余各点提前一个月。结果表明, 黑潮延伸体以北海域存在明显的春秋两次藻华暴发现象, 而冬夏季则消散衰亡。这种现象在南部海域逐渐减弱。
 |
| 图 4 A-E点1998-2018年月平均叶绿素浓度 Fig. 4 Monthly average chl a concentration at points A-E from 1998 to 2018 |
海表叶绿素浓度的季节性变化除了浮游植物生物化学过程外, 还与气象因子例如温度、风速、短波辐射(光强)、以及混合层深度等物理条件相关。图 5为1981-2010年的月平均数据, 显示出目标海域的季节性差异。图 5a显示夏季8月海表气温最高, 冬季2月气温最低。图 5b显示出风速各月份变化明显, 且存在明显的南北差异。风速在北部(A, B, C)1月达到最大值, 5月达到最小值。与北部不同的是, 南部(D, E)点4月开始风速出现小的增幅, 最小值出现在11月份。图 5c显示了各点各月份的短波辐射变化, 从图中可以看出各点变化趋势大体一致, 12月份最低, 7月达到最大。
 |
| 图 5 A-E点1981-2010年气候场气候月平均数据 Fig. 5 Monthly climatology of meteorological field at points A-E from 1981 to 2010 |
混合层的深度变化是体现海水垂向结构变化的重要指标。当混合层深度过深, 将受到水体混合带来的稀释效应, 阻碍浮游植物生长和叶绿素浓度上升。因此混合层的深度变化影响着浮游植物的活动空间和光照强度, 进而影响叶绿素分布。图 5d为南北区域混合层深度的变化, 总体来看, 随着降温对流和风力增强, 混合层在2或3月份深, 夏季浅。在黑潮延伸体主轴附近(C, D), 混合层深度于3月达到最大, 在150m深度左右。
可以看出, 气候态叶绿素的季节变化与光强和混合层的变化基本吻合。3月份之后, 混合层深度迅速减小, 此时光强逐渐增加, 温度升高, 携同冬季积累的营养盐, 导致3-5月份出现高叶绿素浓度。值得一提的是, 混合层深度不仅受到气象因子的影响, 中尺度涡也会导致混合层和密跃层发生变化。例如抽吸作用导致气旋式涡旋中心等密线抬升, 反气旋式涡旋中心等密线下陷, 此机制对中尺度涡范围内浮游植物的生长消亡及其带来的叶绿素浓度分布可能存在影响。
2.3 中尺度涡旋对叶绿素分布的影响对1055个叶绿素数据充足的有效涡旋进行逐一观察后, 发现多数中尺度涡对叶绿素无显著影响, 但是在特定季节中有一定比例的中尺度涡过程能大幅度提高叶绿素浓度。特别是有些涡在延伸体南部提高了涡中心的叶绿素浓度, 形成孤立的叶绿素高值斑块且多数向西传播, 这对于生产力原本较低的南部区域可能有重要意义。反气旋涡多发生在延伸体北部, 在春季和秋季可发生环状的叶绿素高值。气旋涡多发生在延伸体南部, 在春季涡旋可发生中心叶绿素高值, 而在夏、秋、冬季节对叶绿素浓度分布影响不大。
从1055个数据中选取了8个案例展示中尺度涡的典型叶绿素分布(图 6)。图 6a、c、e、g分别为2013年春、夏、秋、冬季的反气旋涡与叶绿素浓度分布示意图, 春季与秋季环状叶绿素高值时有发生, 图 6a中反气旋涡中心叶绿素浓度平均值为0.30mg/m3, 反气旋涡边界处叶绿素浓度平均值为0.93mg/m3, 差值达到0.63mg/m3; 图 6e中反气旋涡中心叶绿素浓度为0.50mg/m3, 边界为0.81mg/m3, 差值为0.31mg/m3, 同样存在环状高值结构。图 6c、g中的夏季与冬季反气旋涡中心与边界处叶绿素浓度皆处于较低水平, 无显著环状叶绿素高值结构产生。图 6b、d、f、h为2016年春、夏、秋、冬季的气旋涡与叶绿素浓度分布示意图。可以看到气旋涡仅在春季出现中心叶绿素高值结构(图 6b), 此时涡中心浓度平均值达到0.66mg/m3, 边界处叶绿素平均浓度仅为0.38mg/m3, 差值达到0.28mg/m3。图 6d、f、h显示夏、秋、冬三季气旋涡并无中心高值结构产生。
 |
| 图 6 四个季节中尺度涡与叶绿素典型分布案例 Fig. 6 Typical cases of mesoscale eddy and chl a distribution for four seasons 注:黑色箭头代表海表面异常流场; a、c、e、g分别为2013年第18周春季、第35周夏季、第44周秋季、第49周冬季反气旋; b、d、f、h分别为2016年第14周春季、第34周夏季、第43周秋季、第51周冬季气旋; 图中空白部分表示此区域遥感数据缺失 |
季节统计结果表明, 叶绿素浓度对中尺度涡有明显响应的占比也呈现季节性特征(图 7)。春季3-5月涡旋对叶绿素影响最大, 135个有效气旋涡中17%存在中心叶绿素高值的特点, 185个有效反气旋涡中33%存在明显的环状叶绿素高值结构。夏季6-8月只存在极少数的气旋涡中心叶绿素高值与反气旋涡环状叶绿素高值结构。秋季9-11月叶绿素环状结构占比达到23%, 但并没有出现气旋涡中心叶绿素高值结构。冬季12-2月叶绿素浓度整体较低, 涡旋的存在仍会略微影响叶绿素浓度分布, 但是不足以产生显著的叶绿素结构。
 |
| 图 7 2012-2018年不同月份中尺度涡叶绿素分布典型结构统计结果 Fig. 7 Statistical results of typical chl a distribution in mesoscale eddies in different months from 2012 to 2018 |
上述这种季节性差异也可能来自于中尺度涡自身数目的季节性差异。为此, 我们对目标区域中尺度涡季节平均数量进行了比较(图 8)。总体来看, 黑潮延伸体区域中尺度涡数量特征略显“夏强冬弱”的特点, 此结果与胡冬等(2018)的统计结果相似, 但各季节生成的气旋涡与反气旋涡的数量季节性差异并不显著, 说明涡旋对叶绿素浓度分布影响的季节性特征并不是由涡旋本身数量的季节变化所单方面决定的。
 |
| 图 8 2012-2018年各季节涡旋平均数量统计 Fig. 8 Statistical results of the mean number of eddies in each season from 2012 to 2018 |
中尺度涡对叶绿素和生态系统的影响一直受到广泛关注, 以往的研究已经取得了许多进展, 主要聚焦在中尺度涡动力过程对营养盐通量输运的作用。Oschlies等(1998)等利用卫星数据同化拟合中尺度涡的生态-环流耦合数值模式, 指出在北大西洋中尺度涡可能贡献了三分之一的深层向海表层营养盐通量。以往研究(Wang et al, 2016; Xu et al, 2019)总结了四种主要的动力机制:涡旋水平方向的平流和混合、中尺度涡环流场对浮游植物的“捕获”限制、中尺度涡的抽吸作用、中尺度涡流场和表层风Ekman输运的相互作用。然而, 以往的研究对于季节性关注不多, 大部分分析都是采用全年统计的结果, 忽略了光和混合层或密跃层季节性变化对藻华暴发的影响。对于渔业资源而言, 很多鱼类的洄游和产卵都存在明显的季节性, 早期生活史是重要的“窗口”期。因此, 我们的观点是有必要考虑这些中尺度涡过程在不同季节的作用, 不能仅考虑营养盐动力学补充过程, 还应包含例如光限制、层化条件等季节性差异因子。
3.1 中尺度涡对叶绿素影响的季节特征我们的研究结果表明, 黑潮延伸体区域的叶绿素分布存在季节性规律, 北部呈现显著的春季藻华和一个规模较小的秋季藻华。南部叶绿素总体浓度低, 为低生产力区域。在夏季、秋季和冬季, 气旋涡中心和周边的叶绿素浓度绝大部分处于较低水平, 极少发现气旋涡能够大幅度提升涡中心的表层叶绿素浓度。然而在春季, 特别是3-5月期间, 从黑潮延伸体分离出来的气旋涡在30°N左右区域向西移动过程中, 一部分气旋涡(占比17%)能保持一个高叶绿素浓度中心, 该区域在空间上与西北太平洋柔鱼产卵场位置存在重叠, 在时间上与柔鱼的产卵高峰期(1-5月)交叉。气旋涡带来的高叶绿素浓度意味着较高的浮游植物浓度, 从而能够在周边低生产力的海域保持一个相对旺盛的生态环境, 支撑起浮游动物和小型鱼类的食物网。柔鱼仔幼体被携带在这样一个生态系统中, 能够保持食物来源, 可能随着气旋涡向西传播重新融入黑潮主轴中, 也可以在传播过程中逃逸来到更南边的区域。2019年3月上海海洋大学渔业资源调查船“淞航号”西北太生物资源调查航次在黑潮延伸体区域的气旋涡边缘区域发现了大量的鱿鱼类幼体聚集, 支撑了上述过程, 相关数据将在另一篇论文中进行详细分析。
从全球角度而言, 中尺度涡对叶绿素浓度产生的作用存在明显的地域差异。例如在东边界上升流区域, 中尺度涡会抑制生产力, 这有可能是中尺度涡把近岸高营养盐水体稀释到了大洋(Gruber et al, 2011)。Gaube等(2014)对不同的区域中尺度涡对近表层叶绿素的影响进行了统计, 发现在西边界流区域气旋式涡旋能够提升表层叶绿素浓度, 而反气旋涡旋降低表层叶绿素浓度, 而南印度洋则相反。与他们的研究相比, 我们更细致的阐明, 在黑潮延伸体南北海域, 只有在春季这个窗口期, 气旋涡才能大幅度提升叶绿素浓度, 且主要发生在涡中心。对于反气旋涡而言, 在更长的窗口期(3-5月, 9-11月), 都会形成叶绿素环结构, 显著增强涡边缘的叶绿素浓度, 降低涡中心的叶绿素浓度。
3.2 混合层变化与叶绿素的季节性变化在混合层气候态平均值分析中, 涡旋的贡献被掩盖, 但对于一个特定涡旋系统而言, 系统内与浮游植物相关的温盐光属性、水体混合、营养盐补充等过程均以涡旋垂向结构为背景场。大洋生态系统动力学难以构建一个普适理论, 但垂向水体混合扰动之后的再层化导致浮游植物暴长是最常见情况。Sverdrup(1953)经典理论指出春季混合层变浅是导致春季藻华暴发的重要原因。Hu等(2011)基于一维FVCOM-NPZD研究藻华, 模型表明风、冷却对流、热力、淡水浮力输入等动力机制导致的各种类型水体扰动后再层化均能快速提升表层水体浮游植物生物量, 揭示了再层化过程和光限制条件对浮游植物的影响机制。
南部气旋涡引发的中心叶绿素高值现象多数出现在春季, 与黑潮以北区域的春季藻华出现的季节基本一致, 意味着除了对于中尺度涡垂向营养盐输运的物理海洋动力学热点之外, 混合层或等密线变化在中尺度涡生态系统中的重要性也需要进一步探讨。气旋涡的流场会导致中心处等密线抬升, 将浮游植物限制在中心较浅的水层中, 同理, 反气旋涡形成中心等密线低陷, 周边等密线抬升弯曲, 浮游植物在环状的周边被限制在光强充足的较浅水层中。然而, 要厘清这种可能机制, 后续需要足够的中尺度涡区域光强垂向分布数据和物理-生物耦合模型来验证。经典的春季藻华理论在大洋中应用并不广泛, 营养盐和其他可能限制浮游植物生长的微量生源要素都可能制约浮游植物的生长和聚集, 因此这一现象需要更深一步的研究。
同样, 目前对于秋季中尺度涡的影响机制也缺乏深入理解。由于数据的局限, 我们虽然注意到秋季气旋涡对于叶绿素浓度影响有限, 但是这种现象背后的机制存在多种可能性: 1)大面海域表层营养盐在夏季消耗殆尽, 2)秋季对流导致混合层加深过快以及光照衰减, 3)表层可能存在较高的浮游动物摄食压力等。因此, 有必要在秋季开展专门针对中尺度涡区域的生物资源量调查, 获取营养盐分布和浮游动物数据。
4 结论与展望本文通过分析黑潮延伸体区域气旋涡和反气旋涡对叶绿素的影响, 得到以下结论:
(1) 黑潮延伸体区域南北空间上存在叶绿素浓度梯度差异, 时间上存在季节性差异。以黑潮延伸体为界限, 北部叶绿素浓度高, 南部叶绿素浓度低。北部存在显著的春季藻华和一个次级规模的秋季藻华, 这一季节特征向南逐渐减弱。
(2) 靠近渔场的黑潮延伸体北部区域, 许多向西传播的反气旋涡虽然会降低中心叶绿素浓度, 但极大增高涡边缘的叶绿素浓度。
(3) 靠近产卵场的黑潮延伸体南部区域, 小部分气旋涡可以形成长时间的高叶绿素斑块且多数向西传播, 有助于支撑一个食物链, 可能起到生物传输带的作用。
(4) 中尺度涡对叶绿素的影响能力存在季节差异。气旋涡中心叶绿素高值多发生在春季, 在其他季节几乎对叶绿素提升没有显著影响。反气旋涡的叶绿素环状结构多发生在春、秋两个季节。
根据上述结论, 我们提出以下后续研究展望:
(1) 以往的研究多以各种中尺度涡营养盐补给的动力机制来解释叶绿素分布, 而我们的研究表明, 至少在黑潮延伸体区域, 中尺度涡叶绿素浓度存在显著季节性特征。同样的涡动力机制下, 叶绿素在不同的季节有不同的响应。因此, 借鉴考虑光限制和混合层深度的Sverdrup藻华理论, 在营养盐垂向补给机制之外, 后续研究还应当重视营养盐背景场、中尺度涡垂向光限制和等密线深度等季节性变化的研究。
(2) 与物理海洋学发展历程相似, 渔业生态系统的研究也逐渐从大尺度气候态逐渐细化到中尺度过程。对于中尺度涡这样典型的中尺度过程, 特定季节对渔业生态系统的贡献可能远远大于其他季节, 有必要在较短的时间尺度上进行研究, 而不建议用全年统计量来进行分析研究。后续研究建议针对渔业的需求, 开展不同季节的中尺度涡生物量资源调查和特定中尺度涡生态结构的跟踪调查。
(3) 中尺度涡的三维物理结构已经获得了极大关注和重视, 并取得丰富的动力学成果。但是中尺度涡的生态动力学研究仍存在巨大的挑战, 如何建立针对性的观测计划, 有效捕捉到这些中小尺度的生物暴发聚集现象并进行持续的三维生化要素监测, 从而获取数据结合物理场通过物理-生物耦合建模来理解生态动力学过程, 是持续开展这方面工作的关键。
参考文献
刘金立, 陈新军, 李纲, 等. 2014. 西北太平洋柔鱼资源综合配置模型及管理策略探讨. 海洋与湖沼, 45(5): 1016-1022 |
汤博, 侯一筠, 殷玉齐, 等. 2019. 北太平洋副热带逆流区中尺度涡旋的统计特征及其分布规律. 海洋与湖沼, 50(5): 937-947 |
余为, 2016.西北太平洋柔鱼冬春生群对气候与环境变化的响应机制研究.上海: 上海海洋大学博士学位论文, 20-21
|
余为, 陈新军, 易倩, 等. 2013. 北太平洋柔鱼早期生活史研究进展. 上海海洋大学学报, 22(5): 755-762 |
胡冬, 陈希, 毛科峰, 等. 2018. 黑潮延伸体邻近区域中尺度涡特征统计分析. 海洋与湖沼, 49(3): 497-511 |
董昌明, 蒋星亮, 徐广珺, 等. 2017. 海洋涡旋自动探测几何方法、涡旋数据库及其应用. 海洋科学进展, 35(4): 439-453 |
Chelton D B, Schlax M G, Samelson R M, 2011. Global observations of nonlinear mesoscale eddies. Progress in Oceanography, 91(2): 167-216 |
Chen X J, Cao J, Chen Y et al, 2011. Effect of the Kuroshio on the spatial distribution of the red flying squid Ommastrephes bartramii in the Northwest Pacific Ocean. Bulletin of Marine Science, 88(1): 63-71 |
Gaube P, McGillicuddy Jr D J, Chelton D B et al, 2014. Regional variations in the influence of mesoscale eddies on near-surface chlorophyll. Journal of Geophysical Research:Ocean, 119(12): 8195-8220 DOI:10.1002/2014JC010111 |
Gruber N, Lachkar Z, Frenzel H et al, 2011. Eddy-induced reduction of biological production in eastern boundary upwelling systems. Nature Geoscience, 4(11): 787-792 DOI:10.1038/ngeo1273 |
Hu S, Chen C S, Ji R B et al, 2011. Effects of surface forcing on interannual variability of the fall phytoplankton bloom in the Gulf of Maine revealed using a process-oriented model. Marine Ecology Progress Series, 427(2): 29-49 |
Kalnay E, Kanamitsu M, Kistler R et al, 1996. The NCEP/NCAR 40-year reanalysis project. Bulletin American Meteorological Society, 77(3): 437-470 DOI:10.1175/1520-0477(1996)077<0437:TNYRP>2.0.CO;2 |
Olson D B, 1991. Rings in the ocean. Annual Review of Earth and Planetary Sciences, 19: 283-311 DOI:10.1146/annurev.ea.19.050191.001435 |
Oschlies A, Garçon V, 1998. Eddy-induced enhancement of primary production in a model of the North Atlantic Ocean. Nature, 394(6690): 266-269 DOI:10.1038/28373 |
Schmidtko S, Johnson G C, Lyman J M, 2013. MIMOC:a global monthly isopycnal upper-ocean climatology with mixed layers. Journal of Geophysical Research:Oceans, 118(4): 1658-1672 DOI:10.1002/jgrc.20122 |
Sverdrup H U, 1953. On conditions for the vernal blooming of phytoplankton. ICES Journal of Marine Science, 18(3): 287-295 DOI:10.1093/icesjms/18.3.287 |
The MODE Group, 1978. The mid-ocean dynamics experiment. Deep Sea Research, 25(10): 859-910 DOI:10.1016/0146-6291(78)90632-X |
Wang M H, Son S, 2016. VIIRS-derived chlorophyll-a using the ocean color index method. Remote Sensing of Environment, 182: 141-149 DOI:10.1016/j.rse.2016.05.001 |
Xu G J, Dong C M, Liu Y et al, 2019. Chlorophyll rings around ocean eddies in the North Pacific. Scientific Reports, 9(1): 2056 DOI:10.1038/s41598-018-38457-8 |
Yu W, Chen X J, Yi Q et al, 2015. A review of interaction between neon flying Squid (Ommastrephes bartramii) and oceanographic variability in the North Pacific Ocean. Journal of Ocean University of China, 14(4): 739-748 DOI:10.1007/s11802-015-2562-8 |