 2021, Vol. 52
2021, Vol. 52中国海洋湖沼学会主办。
文章信息
- 徐飞, 王军强, 蔡文倩, 魏静梅, 韩雪萌. 2021.
- XU Fei, WANG Jun-Qiang, CAI Wen-Qian, WEI Jing-Mei, HAN Xue-Meng. 2021.
- 天津近岸海域大型底栖动物摄食功能群特征及其与环境因子的关系
- MACROZOOBENTHOS FUNCTIONAL FEEDING GROUPS AND ITS RELATIONSHIP WITH ENVIRONMENTAL FACTORS IN THE TIANJIN COASTAL AREA
- 海洋与湖沼, 52(1): 186-195
- Oceanologia et Limnologia Sinica, 52(1): 186-195.
- http://dx.doi.org/10.11693/hyhz20200400108
文章历史
-
收稿日期:2020-04-03
收修改稿日期:2020-04-23




2. 中国环境科学研究院环境基准与风险评估国家重点实验室 北京 100012;
3. 中国科学院南海海洋研究所 广州 510301
2. State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012, China;
3. South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China
大型底栖动物作为河口海湾生态系统中的重要生物组分, 尽管其群落结构及功能指标本身并无直接的生物学特征, 但时空序列上的高低变化却能表征整个生态系统的物质循环和能量流动状况, 反映其栖居地的环境状况, 因此常被人们用以评估河口或近海的生态质量状况(Borja et al, 2009; Van Hoey et al, 2013; Borja, 2014)。随着沿岸经济的快速发展, 受人为干扰的影响, 过去60年来, 以营养盐为代表的大量陆源污染物输入致使近岸海域生态系统受到严重威胁(Xin et al, 2019), 生态系统结构和功能变化显著(Gao et al, 2014), 进而影响到栖居于此的大型底栖动物(陈琳琳等, 2016)。
作为渤海湾最大的沿岸城市, 天津市每年向海输入大量的污染物(Duan et al, 2010)。根据《中国海洋生态环境状况公报》, 2018年天津市直排口排污量为1866万t, 其中COD为615 t。据报道, 天津海域面积在2005-2015年间减少约280 km2。高强度的围填海活动使得该区域的水体交换能力显著下降(靳玉丹等, 2017), 污染物扩散时间延长。多因素叠加致使渤海湾天津近岸海域污染物生态风险状况处于中等以上水平(Chen et al, 2019; Tong et al, 2019), 富营养化严重(秦延文等, 2012; Peng, 2015; 李俊龙等, 2016; 孙欢等, 2017), 栖居于此的大型底栖动物群落受到严重的干扰(张萍等, 2016; 胡桂坤等, 2019), 物种明显小型化、低质化, 群落结构简单(Zhou et al, 2012; 周然等, 2014; Cai et al, 2014)。尤其是在人为干扰严重的蓟运河口、北塘排污口和南、北排水河口区等, 大型底栖动物群落结构更为脆弱(蔡文倩等, 2013)。
大型底栖动物群落功能特征可用来指示其栖居地的生态质量状况, 研究方法分为单一指标法和综合指标法。目前, 我国大型底栖动物综合功能特征指标法主要应用在黄河口(Yang et al, 2017)和石岛湾天鹅泻湖(Hu et al, 2019), 单一的摄食功能群指标法则主要应用在渤海湾(Peng et al, 2013; 蔡文倩等, 2016a, b)、烟台近岸海域(Han et al, 2016)和长江口(Lv et al, 2018)。研究证实, 渤海湾大型底栖动物以小型的碎屑食者如多毛类为主, 群落摄食均匀度较低(Peng et al, 2013; 蔡文倩等, 2016a, b)。
综上所述, 有关天津近岸海域污染物时空分布趋势及大型底栖动物群落结构特征方面的研究开展的较多, 但在摄食功能群方面则研究的较少, 研究所用数据也多是2015年之前。因此, 本研究拟以2015年至2016年3个航次的数据为基础, 分析天津近岸海域大型底栖动物摄食功能群组成现状, 辨析影响其变化的主要环境因子, 判断天津近岸海域大型底栖动物摄食功能群变化状况, 为渤海大型底栖动物生态学研究提供基础数据支撑。
1 材料与方法 1.1 样品采集与数据获取为研究天津近岸海域大型底栖动物群落状况, 于2015年9月、2016年5月和2016年9月进行了3个航次的生物和环境样品采集工作(图 1)。采用0.05 m2的箱式采泥器采集大型底栖动物样品, 每个站位设3个平行样, 样品冲洗后放入样品瓶中并用75%酒精固定。同时, 每个站位现场采集500 mL水, 迅速装入样品瓶后加入2-3滴浓硫酸, 冷藏保存至实验室测定营养盐; 采集50 mL水迅速装入棕色玻璃瓶, 冷藏保存并带到实验室进行测定总有机碳; 采集1 L水并现场过滤, 滤膜用锡箔纸包好冷藏至实验室测定叶绿素a含量。同步现场采集表层泥样, 用锡箔纸包好后送至实验室测定沉积物粒度, 粒度分级依据Buchanan(1984)中的方法。环境因子测定方法详见国家环保总局(2002)推荐的方法。
 |
| 图 1 天津近岸海域采样站位示意图 Fig. 1 Map of sampling site in the Tianjin coastal area |
将所有大型底栖动物划分为肉食者(carnivore, CA)、碎屑食者(detritivore, DE)、植食者(herbivore, HE)、杂食者(omnivore, OM)、浮游生物食者(planktivore, PL)等5个摄食功能群。各物种的摄食功能群分组参考蔡文倩等(蔡文倩等, 2016a, b)。参考以往的研究经验, 仅采用任一站位中丰度占该站点总丰度比例大于5%的物种进入摄食功能群特征分析(Peng et al, 2013)。摄食功能群特征指标包括各摄食类群物种组成、丰度、生物量和摄食多样性(H’FD)。其中, 摄食多样性计算方法如下。
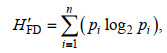
式中, pi为样方中的第i摄食群所占的比例。本研究中, n值为5。
1.3 数据统计分析Spearman相关分析用以检验各摄食功能群特征参数(物种数、丰度和生物量)与环境因子之间的相关性(双尾检验); 采用主成分分析(principal component analysis, PCA)筛选主要的环境因子, 并依据其分析结果对所有站位进行分组, 据此采用MANOVA法判断环境因子时空差异是否显著。上述分析在SPSS 13.0软件包中进行。聚类法用以分析大型底栖动物摄食功能群结构。数据分析前, 原始丰度数据经四次方根转换后, 构建Bray-Curtis相似性矩阵。聚类分析在PRIMER 6.0软件包中进行。RDA分析用以表征各摄食群分布和环境因子之间的关系, 分析在Canoco 5.0软件包中进行。摄食多样性指数图和采样站位示意图由Surfer 8.0软件包进行绘制。
2 结果与分析 2.1 环境因子各环境因子的取值范围见表 1。根据《海水水质标准》(GB 3097-1997), 23个站位的活性磷酸盐值均不超过0.015 mg/L, 满足Ⅰ类水质标准, 占比38.33%; 22个站位的活性磷酸盐值均位于0.015-0.03 mg/L之间, 满足Ⅱ类水质标准, 占比36.67%; 6个站位的活性磷酸盐值位于0.03-0.045 mg/L之间, 满足Ⅳ类水质要求, 占比10%;剩余9个站位的活性磷酸盐值大于0.045 mg/L, 满足Ⅴ类水质标准, 占比15%。从空间分布的角度看, 整体上活性磷酸盐浓度分布无明显的规律, 但Ⅴ类水质标准的站位均位于东沽航道内, 其中最大值出现在2015年9月航次的9号站, 浓度超出Ⅱ类水质标准7倍。总的来看, 其他9个环境因子空间分布也无明显的分布规律, 但与活性磷酸盐一样, 营养盐、总无机碳和叶绿素a的高值区均集中在东沽航道内的8-10号站, 且航道内各站位底质以粉砂为主, 其次为黏土。
| 活性磷酸盐(mg/L) | 总磷(mg/L) | 硝酸盐(mg/L) | 铵氮(mg/L) | 总有机碳(mg/L) | 总氮(mg/L) | 叶绿素a (mg/L) | 黏土(%) | |
| 最大值 | 0.21 | 2.24 | 4.14 | 1.48 | 211.20 | 15.37 | 21.92 | 46.29 |
| 最小值 | 0.01 | 0.01 | 0.00 | 0.01 | 2.20 | 0.04 | 2.66 | 11.95 |
| 均值 | 0.03 | 0.25 | 0.71 | 0.17 | 8.44 | 4.74 | 7.94 | 20.77 |
| 标准差 | 0.03 | 0.48 | 0.68 | 0.21 | 26.67 | 3.68 | 4.48 | 5.69 |
| 粉砂(%) | 极细砂(%) | 细砂(%) | 中砂(%) | 粗砂(%) | 极粗砂(%) | 砂砾(%) | 中值粒径 | |
| 最大值 | 72.04 | 18.98 | 18.04 | 18.57 | 25.01 | 20.39 | 1.35 | 45.83 |
| 最小值 | 34.77 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.94 |
| 均值 | 59.98 | 7.06 | 3.18 | 1.66 | 3.51 | 3.71 | 0.14 | 10.26 |
| 标准差 | 8.27 | 5.01 | 4.09 | 3.27 | 5.55 | 5.24 | 0.29 | 6.29 |
根据环境因子的PCA分析结果, 1-7号站位为第Ⅰ组, 集中在研究区北部; 8-10号为第Ⅱ组, 集中在研究区东沽航道内; 11-14号为第Ⅲ组, 集中在东沽航道外的延伸线上; 15-20号为第Ⅳ组, 集中在研究区南部。在进行MANOVA分析时, 17个环境因子为因变量, 3个航次和4个组为固定因子。结果表明, 3个航次之间、4站组之间的环境因子差异显著(P < 0.05), 4个组之间的环境差异也显著(P < 0.05), 而不同航次不同组之间的两两比较结果也显示差异显著(P < 0.05)。
单个环境因子组间方差分析结果表明, 水体中的总磷和总有机碳与沉积物中的黏土、粉砂、极细砂、细沙、粗砂、极粗砂、中值粒径和粒径多样性在4个组间未表现出显著性差异(P > 0.05), 其他的环境因子均表现出了显著性差异(P < 0.05);沉积物中的粉砂、中砂、粗砂、中值粒径和粒径多样性在3个航次未表现出显著性差异(P > 0.05), 其他因子均表现出了显著性差异(P < 0.05)。总体上看, 不同航次不同组之间, 水体中的磷酸盐、铵盐和叶绿素a以及沉积物中的极粗砂和砂砾含量表现出了显著性差异, 其他因子均未表现出显著性差异。
2.2 摄食群特征 2.2.1 物种组成3个航次共获得大型底栖动物110种, 其中包括碎屑食者44种, 肉食者34种, 杂食者17种以及浮游生物食者15种。其中, 碎屑食者以多毛动物为优势类群, 占比高达55%, 其次为甲壳动物, 占比达20%;肉食者以多毛动物为优势类群, 占比达53%, 其余4个类群占比差别不大; 杂食者以甲壳和多毛动物为主, 分别占比为41%和24%;浮游生物食者以软体动物占据优势地位, 占比高达73%, 其次为甲壳动物, 占比为20%。由此可见, 研究区多毛动物分布于4个食性之中, 且均占有一定的比例, 可见该类群的摄食生态位最大, 软体动物和甲壳动物次之, 棘皮动物最小(图 2)。
 |
| 图 2 天津近岸海域3个航次大型底栖动物摄食功能群组成 Fig. 2 Composition of macrozoobenthos functional feeding groups for the three voyages in the Tianjin coastal area |
2015年9月航次, 除多毛类残体无相应的摄食功能群外, 碎屑食者和肉食者占据优势地位, 杂食者最少; 2016年5月和2016年9月航次的各类群分布比例与2015年9月航次的相同(表 2)。总的来说, 3个航次的肉食者和碎屑食者整体占比基本一致, 均处于优势地位, 而杂食者和浮游生物食者则占比较低。以优势类群碎屑食者和肉食者为例, 2个摄食群物种数均在近岸区较多, 如1、4、6、12、13及18站。其中, 碎屑食者整体上呈现北高南低的空间分布模式, 而肉食者则无明显规律。
| 摄食群 | 2015.09 | 2016.05 | 2016.09 |
| 肉食者CA | 24(36%) | 28(37%) | 22(38%) |
| 碎屑食者DE | 27(41%) | 29(39%) | 22(38%) |
| 杂食者OM | 9(14%) | 8(11%) | 9(15%) |
| 浮游生物食者PL | 6(9%) | 10(13%) | 6(9%) |
| 总计 | 66(100%) | 75(100%) | 59(100%) |
| 注:括号内数据代表整体占比, 下同 | |||
3个航次中, 碎屑食者的丰度均较高, 浮游生物食者均最低。总体上, 2015年9月航次摄食功能群平均丰度最高, 2016年9月最低(表 3)。从空间分布的角度看, 2015年9月航次, 整体上以海河口东沽航道为界, 各摄食群丰度分布基本呈现北高南低的模式。其中, OM丰度空间分布呈从近岸至离岸海域逐渐增高的趋势。2016年5月航次, 高密度的碎屑食者丝异蚓虫使得东沽航道内的8号和9号站DE占据绝对优势。其中, DE和CA丰度呈从近岸至离岸海域逐渐降低的趋势。2016年9月航次, 4个摄食功能群丰度空间分布基本呈从近岸至离岸海域逐渐升高的趋势。总的来看, 3个航次的OM丰度空间分布呈从东沽航道向外逐渐增加的趋势(图 3)。
| 摄食群 | 2015.09 | 2016.05 | 2016.09 |
| 肉食者CA | 119.00(32%) | 92.33(27%) | 44.00(29%) |
| 碎屑食者DE | 196.33(53%) | 89.00(27%) | 31.00(21%) |
| 杂食者OM | 31.00(8%) | 48.33(14%) | 72.00(48%) |
| 浮游生物食者PL | 27.67(7%) | 108.67(32%) | 3.33(2%) |
| 总计 | 374.00(100%) | 338.33(100%) | 151.67(100%) |
 |
| 图 3 天津近岸海域大型底栖动物各摄食功能群丰度空间分布图 Fig. 3 Spatial distributions of macrozoobenthos feeding groups abundance in the Tianjin coastal area 注: a. 2015年9月航次; b. 2016年5月航次; c. 2016年9月航次 |
3个航次中, 杂食者生物量占据显著优势, 且以2016年9月航次的最高; 肉食者无明显变化; 碎屑食者生物量明显较低, 且基本无变化; 浮游生物食者生物量占比仅次于杂食者, 2015年9月和2016年5月相差不大, 但2016年9月航次下降较为明显(表 4)。从空间分布的角度看, 2015年9月航次, CA、DE和OM空间分布呈现由近岸至离岸逐渐升高的趋势; 2016年5月航次, 除PL外, 其他3个功能群生物量分布模式与2015年9月的基本一致; 2016年9月航次, 除OM生物量空间分布呈从近岸至离岸逐渐升高的趋势外, 其他3个功能群的空间分布无明显的规律(图 4)。
| 摄食群 | 2015.09 | 2016.05 | 2016.09 |
| 肉食者CA | 4.62(14%) | 10.52(12%) | 8.46(14%) |
| 碎屑食者DE | 2.24(7%) | 13.49(16%) | 0.56(1%) |
| 杂食者OM | 15.13(47%) | 37.77(43%) | 47.04(80%) |
| 浮游生物食者PL | 10.20(32%) | 25.47(29%) | 2.75(5%) |
| 总计 | 32.20(100%) | 87.25(100%) | 58.81(100%) |
 |
| 图 4 天津近岸海域大型底栖动物各摄食功能群生物量空间分布图 Fig. 4 Spatial biomass distributions of macrozoobenthos feeding groups in the Tianjin coastal area 注: a. 2015年9月航次; b. 2016年5月航次; c. 2016年9月航次 |
2015年9月航次大型底栖动物各站位摄食多样性平均值为1.37±0.74, 2016年5月航次的为1.65±0.42, 2016年9月航次的为0.67±0.54。3个航次相比, 不论是总平均摄食多样性指数值, 还是各站位摄食多样性指数值, 基本以2016年5月航次的最高, 2016年9月航次的最低。3个航次中, 摄食多样性指数沿9-12号站逐渐升高, 而12-14站略有降低; 与同一航次其他站位相比, 8-14号站的多样性指数值均较低(图 5)。
 |
| 图 5 天津近岸海域大型底栖动物各站位摄食多样性指数值 Fig. 5 Index values of feeding diversity for all sampling sites in the Tianjin coastal area 注: a. 2015年9月航次; b. 2016年5月航次; c. 2016年9月航次 |
SIMPEROF分析表明, 在23.19%相似度下, 所有站位为1个聚类组(π=4.87, P < 0.01);进一步以46.73%相似度为划分依据(π=1.62, P < 0.01), 2016年9月航次的8号(48)、9号(49)和10号站(50)为1个组, 剩余的57个站位为1个组; 以80.15%相似度为划分依据(π=0.56, P < 0.01)的情况下, 所有站位可划分为10个组。以42号站(2016年9月航次的2号站)为边界, 无证据表明可以划分为其他任何有显著差异的组(相似度为88.17%, π=0.14, P > 0.05)。故而, 选取80.15%的相似度为最终划分依据, 将3个航次60个站位共划分为10个聚类组, 分别为第1组(44号和60号站)、第2组(58号)、第3组(51号)、第4组(9号和30号)、第5组(17、23、29号)、第6组(10号站)、第7组(38号站)、第8组(49、50号站)、第9组(48号站), 剩余的47个站为第10组。由此可见, 第6-9聚类组均为3个航次的8-10号站, 而ANOSIM分析结果表明10个聚类组之间差异显著(R=0.727, P < 0.01), 也说明了3个航次航道内的8-10号站的摄食功能群与其他站位的差异显著。
2.3 各摄食群丰度与环境因子的关系RDA分析结果表明, 3个航次中, 16个环境因子与4个摄食功能群前两个排序轴的相关性分别为0.45和0.29, 前两个排序轴累计可解释变异度为87.47%, 分析结果可靠。由图 6可知, 总氮与浮游生物食者分布的相关性最高, 其次为总磷、中砂含量百分比与叶绿素a; 总氮与肉食者分布的相关性最高, 其次为硝酸盐和叶绿素a; 极细砂含量与碎屑食者分布的相关性最高, 其次为硝酸盐和叶绿素a; 总有机碳与杂食者的相关性最高, 其次为砂砾含量百分比和铵盐。
 |
| 图 6 3个航次摄食群丰度与环境因子之间的关系 Fig. 6 Relation between abundance of feeding groups and environmental parameters 注: Axis1和Axis2表示解释环境变异量贡献最大的两组。粗砂代表任一站位中的粗砂含量百分比, 中砂、黏土、砂砾、极粗砂、粉砂、细砂、极细砂同上 |
研究证实, 浊度高的海域不适合藻类生长(蔡文倩等, 2016b)。渤海湾较高的浊度限制了藻类的生长, 从而使得栖息于此的大型底栖动物较少营植食生活方式, 而本研究中3个航次均未采集到植食者, 也一定程度上验证了先前的研究结果。如上文所示, 4个摄食功能群中均有多毛类, 且在碎屑食者和肉食者中占据绝对优势地位, 如2016年5月航次海河口东沽航道内8号站的碎屑食者小头虫(Capitella capitata)密度高达1967 ind./m2, 2015年9月航次航道内9号站的碎屑食者毛鳃虫(Cirriformia filigera)密度高达600 ind./m2, 远高于其他摄食群。此外, 浮游生物食者中软体动物占据绝对优势地位, 尤其是小型双壳类如内肋蛤。这些均说明与以往的研究结果相比, 天津近岸海域大型底栖动物群落小型化、低质化的趋势并未改善(蔡文倩等, 2012, 2013; Zhou et al, 2012; 张壮壮等, 2015), 且多毛类及小型双壳类的丰度显著增加(如2015年9月航次中, 两者所占比例为67.16%, 2016年5月航次为66.23%)。
海河口接收大量的陆源污染物, 是渤海湾污染最严重的区域之一(Cai et al, 2014), 其污染效应对大型底栖动物群落的影响呈现沿河口向海逐渐减弱的空间分布趋势(秦延文等, 2012; Cai et al, 2014; 孙欢等, 2017), 营碎屑生活的小型多毛类和双壳类丰度和生物量会呈现沿河口向外海逐渐增加的空间分布趋势(蔡文倩等, 2016a, b), 这与本研究中DE摄食群丰度和生物量的空间分布模式一致。此外, 东沽航道频繁的船舶活动带来的生活污水以及海河口输入的生活污水使得该区域及邻近海域污染物浓度较高(孙欢等, 2017), 营养盐丰富, 导致耐有机污染的丝异蚓虫(Heteromastus filiformis)、小头虫、内肋蛤(Endopleura lubrica)等在该区域采样站位占据绝对优势地位, 由此导致这些站的大型底栖动物摄食功能群与其他站之间存在显著差异, 如本研究聚类分析结果所示, 而这也与以往的群落结构研究结果一致(蔡文倩等, 2012, 2013)。
3.2 摄食功能群组成与环境因子的关系由上文可知, 渤海湾天津近岸海域每年接收其沿岸入海河流和排污口输入的大量污染物(Duan et al, 2010; Wang et al, 2019, 2020), 水体和沉积物中的营养盐浓度居高不下(Peng, 2015; 宋兵魁等, 2019)。此外, 围海造陆等活动导致近岸海域纳潮能力降低(聂红涛等, 2008), 水流速度变缓慢(靳玉丹等, 2017), 污染物大量堆积在近岸海域(Cai et al, 2014), 进而造成栖息于此的敏感型生物大量死亡(Peng et al, 2013), 而一些厌氧型的耐污种则得到迅速增殖的机会。本研究中, 海河口东沽航道内的8-10号站的营养盐、叶绿素a等环境因子含量基本高于其他站位, 而聚类分析也表明3个航次的大型底栖动物摄食功能群与其他站位也存在明显差异。此外, 天津港及其南部的东沽航道内运行或停靠的船舶, 向上述区域排放上万吨的氮氧化物(Chen et al, 2016; Wan et al, 2020)。频繁的航运和污染物的大量排放使得该区域大型底栖动物群落以耐污型的碎屑食者和肉食者为主(Zheng et al, 2011), 而多为敏感种的杂食者棘刺锚参、小型蟹类等则较少。
尽管DIN作为渤海湾主要的污染物, 其春季和夏季的DIN浓度自2000年起就逐渐增加(Qiao et al, 2017; Liu et al, 2019), 但这并未直接促进水体中叶绿素浓度的增加(Qiao et al, 2017)。研究证实, 不同的浮游植物需要的营养盐也不同(Schlüter et al, 2000), 而本研究中RDA分析结果中各摄食群与营养盐的显著相关关系也在一定程度上验证了此结论。研究证实, 水动力条件、活性磷酸盐和硅酸盐是决定浮游植物分布的主要环境因子, 如Peng等(2012)研究发现渤海湾营养盐浓度呈现北高南低的空间分布, 相应地, 浮游植物丰度呈现出相同的空间分布模式。作为营浮游生物食者的大型底栖动物来说, 丰富的食物来源一定程度上刺激了其丰度剧增而成为绝对优势种(蔡文倩等, 2013, 2016b), 如蔡文倩等(2013)研究发现营浮游生物食性的凸壳肌蛤在渤海湾北部海域的丰度较高, 而本研究中凸壳肌蛤的丰度在北部海域也较高, 验证了此结论。事实上, MANOVA分析结果也说明东沽航道内站位的环境与其他站位明显不同, 这也与摄食功能群时空变化分析结果一致。由此可见, 营养盐是驱动天津近岸海域大型底栖动物摄食功能群组成变化的主要环境因子。
3.3 大型底栖动物摄食功能群的变化以往的研究证实, 渤海湾大型底栖动物摄食功能群物种数均以碎屑食者和肉食者占优, 而杂食者和浮游生物食者的最低(Peng et al, 2013; 蔡文倩等, 2016b)。同时, 海河口内大型底栖动物摄食功能群结构组成与其他区域具有显著的差异(蔡文倩等, 2016a), 这与本研究海河口东沽航道大型底栖动物摄食群与其他区域差异显著的结果一致。与2011年5、9月航次的研究结果相比, 最显著的差异在于这两个航次浮游生物食者的生物量和丰度占优。事实上, 2011年2个航次采集到大量的凸壳肌蛤和长偏顶蛤, 仅这二者的丰度和生物量整体占比均超过50%, 从而导致浮游生物食者的丰度和生物量最高。但是, 本研究2016年9月航次未采集到上述2种双壳类, 2016年5月和2015年9月航次采集量也较少, 这主要可能与5个航次的站位设置差异有关。如2011年两个航次凸壳肌蛤单站密度高达124050 ind./m2, 而其密度较高的站位并未在本研究3个航次中设置(蔡文倩等, 2013)。
由上文可知, 2011年2个航次的绝对优势种为小型双壳类凸壳肌蛤, 而本研究的大型底栖动物摄食群组成均以碎屑和肉食者为主, 结合摄食多样性指数分析结果, 一定程度上说明了渤海湾天津近岸海域大型底栖动物群落摄食功能群组成状况仍未发生明显变化。此外, 研究表明, 短时间(6年)对生态系统的保护, 长江口潮间带营浮游生物食性和植食者的大型底栖动物丰度显著增加, 而长时间(18年)的保护后, 大型底栖动物物种数、丰度和生物量均显著增加(Lv et al, 2018)。然而, 由上文可知, 在渤海湾天津近岸海域, 2011年和2015-2016年的大型底栖动物摄食功能群结构较为相似, 这在一定程度上也反映了急需进一步加强天津近岸海域海洋生态保护修复工作, 提升该区域生态系统健康水平, 为京津冀一体化经济发展打下坚实的基础。
4 结论(1) 天津近岸海域大型底栖动物摄食功能群以碎屑食者为主, 且碎屑食者结构指标呈现北高南低的空间分布模式;
(2) 聚类分析和多元方差分析结果表明, 海河口东沽航道内的摄食功能群结构组成与其他区域明显不同;
(3) 营养盐是驱动天津近岸海域大型底栖动物摄食功能群结构变化的环境因子;
(4) 与2011年相比, 天津近岸海域大型底栖动物群落摄食功能群组成状况未发生明显变化。
孙欢, 张秋丰, 屠建波, 等. 2017. 渤海湾天津近岸海域化学需氧量的分布特征、影响因素及与富营养化的关系. 海洋环境科学, 36(3): 343-348, 371 |
李俊龙, 郑丙辉, 张铃松, 等. 2016. 中国主要河口海湾富营养化特征及差异分析. 中国环境科学, 36(2): 506-516 DOI:10.3969/j.issn.1000-6923.2016.02.030 |
宋兵魁, 齐树亭, 李斯, 等. 2019. 渤海湾氮、磷营养盐在水体和沉积物中的分布特征及其相互关系. 海洋学研究, 37(1): 83-90 |
张萍, 缴建华, 孙万胜, 等. 2016. 渤海湾天津近岸海域大型底栖动物群落结构及次级生产力的初步研究. 大连海洋大学学报, 31(3): 324-330 |
张壮壮, 王琳, 杨文波, 等. 2015. 渤海湾近海栖息地变化对大型底栖动物群落结构的影响. 中国水产科学, 22(1): 106-112 |
陈琳琳, 王全超, 李晓静, 等. 2016. 渤海南部海域大型底栖动物群落演变特征及原因探讨. 中国科学:生命科学, 46(9): 1121-1134 |
国家环保总局. 2002. 水和废水监测分析方法. 4版. 北京: 中国环境科学出版社
|
周然, 覃雪波, 彭士涛, 等. 2014. 海湾大型底栖动物调查及与环境因子的相关性. 生态学报, 34(1): 50-58 |
胡桂坤, 秦璐璐, 李郁郁, 等. 2019. 基于ABC曲线的天津潮间带生物群落受扰动的分析. 天津科技大学学报, 34(5): 57-62 |
秦延文, 张雷, 郑丙辉, 等. 2012. 渤海湾岸线变化(2003-2011年)对近岸海域水质的影响. 环境科学学报, 32(9): 2149-2159 |
聂红涛, 陶建华. 2008. 渤海湾海岸带开发对近海水环境影响分析. 海洋工程, 26(3): 44-50 |
靳玉丹, 张秋丰, 李希彬, 等. 2017. 天津海域围填海工程对渤海湾水交换的影响研究. 海洋通报, 36(5): 578-584 |
蔡文倩, 刘静, 周娟, 等. 2016a. 基于生物量的大型底栖动物功能摄食群结构及生态质量评价. 生物多样性, 24(9): 1045-1055 |
蔡文倩, 刘录三, 乔飞, 等. 2012. 渤海湾大型底栖生物群落结构变化及原因探讨. 环境科学, 33(9): 3104-3109 |
蔡文倩, 林岿璇, 朱延忠, 等. 2016b. 基于大型底栖动物摄食群上的生态质量评价. 中国环境科学, 36(9): 2865-2873 |
蔡文倩, 孟伟, 刘录三, 等. 2013. 渤海湾大型底栖动物群落优势种长期变化研究. 环境科学学报, 33(8): 2332-2340 |
Borja Á, 2014. Grand challenges in marine ecosystems ecology. Frontiers in Marine Science, 1: 1 |
Borja Á, Miles A, Occhipinti-Ambrogi A et al, 2009. Current status of macroinvertebrate methods used for assessing the quality of European marine waters:implementing the Water Framework Directive. Hydrobiologia, 633(1): 181-196 DOI:10.1007/s10750-009-9881-y |
Buchanan J B, 1984. Sediment analysis. In: Holme N A, McIntyre A D eds. Methods for the Study of Marine Benthos. Oxford: Blackwell Scientific Publications, 41-65
|
Cai W Q, Borja Á, Liu L S et al, 2014. Assessing benthic health under multiple human pressures in Bohai Bay (China), using density and biomass in calculating AMBI and M-AMBI. Marine Ecology, 35(2): 180-192 DOI:10.1111/maec.12067 |
Chen D S, Zhao Y H, Nelson P et al, 2016. Estimating ship emissions based on AIS data for port of Tianjin, China. Atmospheric Environment, 145: 10-18 DOI:10.1016/j.atmosenv.2016.08.086 |
Chen M Q, Gan Z W, Qu B et al, 2019. Temporal and seasonal variation and ecological risk evaluation of flame retardants in seawater and sediments from Bohai Bay near Tianjin, China during 2014 to 2017. Marine Pollution Bulletin, 146: 874-883 DOI:10.1016/j.marpolbul.2019.07.049 |
Duan L P, Song J M, Li X G et al, 2010. Distribution of selenium and its relationship to the eco-environment in Bohai Bay seawater. Marine Chemistry, 121(1-4): 87-99 DOI:10.1016/j.marchem.2010.03.007 |
Gao X L, Zhou F X, Chen C T A, 2014. Pollution status of the Bohai Sea:an overview of the environmental quality assessment related trace metals. Environment International, 62: 12-30 DOI:10.1016/j.envint.2013.09.019 |
Han Q X, Jiang X M, Wang X B, 2016. The Polychaete feeding guild composition in the Sishili Bay, the northern Yellow Sea, China. Journal of the Marine Biological Association of the United Kingdom, 96(5): 1083-1092 DOI:10.1017/S0025315415001873 |
Hu C Y, Dong J Y, Gao L J et al, 2019. Macrobenthos functional trait responses to heavy metal pollution gradients in a temperate lagoon. Environmental Pollution, 253: 1107-1116 DOI:10.1016/j.envpol.2019.06.117 |
Liu X H, Liu D Y, Wang Y J et al, 2019. Temporal and spatial variations and impact factors of nutrients in Bohai Bay, China. Marine Pollution Bulletin, 140: 549-562 DOI:10.1016/j.marpolbul.2019.02.011 |
Lv W W, Zhou W Z, Zhao Y L, 2018. Macrobenthos functional groups as indicators of ecological restoration in reclaimed intertidal wetlands of China's Yangtze Estuary. Regional Studies in Marine Science, 22: 93-100 DOI:10.1016/j.rsma.2018.06.003 |
Peng S T, 2015. The nutrient, total petroleum hydrocarbon and heavy metal contents in the seawater of Bohai Bay, China:temporal-spatial variations, sources, pollution statuses, and ecological risks. Marine Pollution Bulletin, 95(1): 445-451 DOI:10.1016/j.marpolbul.2015.03.032 |
Peng S T, Qin X B, Shi H H et al, 2012. Distribution and controlling factors of phytoplankton assemblages in a semi-enclosed bay during spring and summer. Marine Pollution Bulletin, 64(5): 941-948 DOI:10.1016/j.marpolbul.2012.03.004 |
Peng S T, Zhou R, Qin X B et al, 2013. Application of macrobenthos functional groups to estimate the ecosystem health in a semi-enclosed bay. Marine Pollution Bulletin, 74(1): 302-310 DOI:10.1016/j.marpolbul.2013.06.037 |
Qiao Y H, Feng J F, Cui S F et al, 2017. Long-term changes in nutrients, chlorophyll a and their relationships in a semi-enclosed eutrophic ecosystem, Bohai Bay, China. Marine Pollution Bulletin, 117(1-2): 222-228 DOI:10.1016/j.marpolbul.2017.02.002 |
Schlüter L, Møhlenberg F, Havskum H et al, 2000. The use of phytoplankton pigments for identifying and quantifying phytoplankton groups in coastal areas:testing the influence of light and nutrients on pigment/chlorophyll a ratios. Marine Ecology Progress Series, 192: 49-63 DOI:10.3354/meps192049 |
Tong Y F, Chen L, Liu Y et al, 2019. Distribution, sources and ecological risk assessment of PAHs in surface seawater from coastal Bohai Bay, China. Marine Pollution Bulletin, 142: 520-524 DOI:10.1016/j.marpolbul.2019.04.004 |
Van Hoey G, Permuy D C, Vandendriessche S et al, 2013. An ecological quality status assessment procedure for soft-sediment benthic habitats:weighing alternative approaches. Ecological Indicators, 25: 266-278 DOI:10.1016/j.ecolind.2012.09.024 |
Wan Z, Ji S J, Liu Y T et al, 2020. Shipping emission inventories in China's Bohai Bay, Yangtze River Delta, and Pearl River Delta in 2018. Marine Pollution Bulletin, 151: 110882 DOI:10.1016/j.marpolbul.2019.110882 |
Wang Q Q, Li H L, Zhang Y et al, 2019. Evaluations of submarine groundwater discharge and associated heavy metal fluxes in Bohai Bay, China. Science of the Total Environment, 695: 133873 DOI:10.1016/j.scitotenv.2019.133873 |
Wang Q Q, Li H L, Zhang Y et al, 2020. Submarine groundwater discharge and its implication for nutrient budgets in the western Bohai Bay, China. Journal of Environmental Radioactivity, 212: 106132 DOI:10.1016/j.jenvrad.2019.106132 |
Xin M, Wang B D, Xie L P et al, 2019. Long-term changes in nutrient regimes and their ecological effects in the Bohai Sea, China. Marine Pollution Bulletin, 146: 562-573 DOI:10.1016/j.marpolbul.2019.07.011 |
Yang W, Li X X, Sun T et al, 2017. Macrobenthos functional groups as indicators of ecological restoration in the northern part of China's Yellow River Delta Wetlands. Ecological Indicators, 82: 381-391 DOI:10.1016/j.ecolind.2017.06.057 |
Zheng B H, Zhao X R, Liu L S et al, 2011. Effects of hydrodynamics on the distribution of trace persistent organic pollutants and macrobenthic communities in Bohai Bay. Chemosphere, 84(3): 336-341 DOI:10.1016/j.chemosphere.2011.04.006 |
Zhou H, Zhang Z N, Liu X S et al, 2012. Decadal change in sublittoral macrofaunal biodiversity in the Bohai Sea, China. Marine Pollution Bulletin, 64(11): 2364-2373 DOI:10.1016/j.marpolbul.2012.08.014 |