 2021, Vol. 52
2021, Vol. 52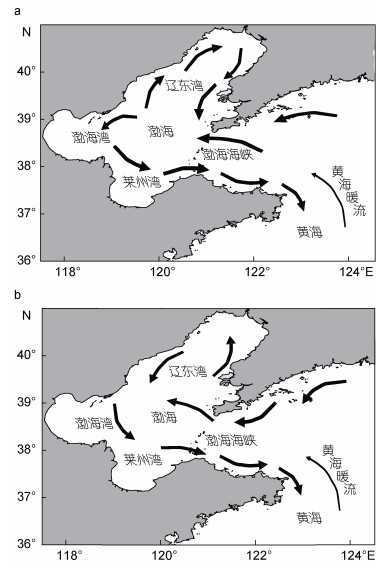

中国海洋浮游植物和赤潮物种的生物多样性研究进展(一): 渤海

http://dx.doi.org/10.11693/hyhz20200900245
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 陈楠生, 黄海龙. 2021.
- CHEN Nan-Sheng, HUANG Hai-Long. 2021.
- 中国海洋浮游植物和赤潮物种的生物多样性研究进展(一): 渤海
- ADVANCES IN THE STUDY OF BIODIVERSITY OF PHYTOPLANKTON AND RED TIDE SPECIES IN CHINA (Ⅰ): THE BOHAI SEA
- 海洋与湖沼, 52(2): 346-362
- Oceanologia et Limnologia Sinica, 52(2): 346-362.
- http://dx.doi.org/10.11693/hyhz20200900245
文章历史
-
收稿日期:2020-09-03
收修改稿日期:2020-10-21
引用本文
陈楠生, 黄海龙. 2021. 中国海洋浮游植物和赤潮物种的生物多样性研究进展(一): 渤海[J]. 海洋与湖沼, 52(2): 346-362

CHEN Nan-Sheng, HUANG Hai-Long. 2021. ADVANCES IN THE STUDY OF BIODIVERSITY OF PHYTOPLANKTON AND RED TIDE SPECIES IN CHINA (Ⅰ): THE BOHAI SEA[J]. Oceanologia et Limnologia Sinica, 52(2): 346-362.
中国海洋浮游植物和赤潮物种的生物多样性研究进展(一): 渤海
1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室 青岛 266071;
2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室 青岛 266237;
3. 中国科学院海洋大科学研究中心 青岛 266071
2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室 青岛 266237;
3. 中国科学院海洋大科学研究中心 青岛 266071
摘要:渤海是我国最大的半封闭型内海,也是重要的海洋生物产卵场、育幼场和索饵场。其陆源径流丰富,环境异质性高,具有很高的浮游植物物种多样性,其中包括大量的赤潮物种。该海域赤潮频发对水产养殖、生态环境、旅游业等造成了很大影响。因此,针对该海域浮游植物物种多样性的研究具有极其重要的意义。本文整理了1957—2019年间46个主要航次的调查资料,比较分析了这些航次中鉴定的浮游植物组成以及相对丰度,并跟踪分析了近20年来渤海的主要赤潮事件,重点描述了赤潮物种多样性,并详细阐述了常见赤潮物种。在航次鉴定的140个赤潮物种中,包括硅藻78种,甲藻52种,定鞭藻5种,绿藻4种和蓝藻1种。本文为比较研究包括渤海海域在内的不同海域浮游植物和赤潮物种的多样性,探索海洋生物多样性的起源和演化规律,解读宏基因组学及宏条形码分析结果提供了基础和参考。
关键词:渤海 生物多样性 航次调查 赤潮物种 浮游植物
ADVANCES IN THE STUDY OF BIODIVERSITY OF PHYTOPLANKTON AND RED TIDE SPECIES IN CHINA (Ⅰ): THE BOHAI SEA
1. CAS Key Laboratory of Marine Ecology and Environmental Science, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China;
2. Functional Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China;
3. Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
2. Functional Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China;
3. Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
Abstract: The Bohai Sea is the largest semi-closed inland sea in China. It is an important spawning, nursery, and feeding ground for marine organisms. It features rich runoffs from land sources, high environmental heterogeneity, and high phytoplankton species diversity. Among them, there are a large number of red tide species, which promote frequent occurrence of blooms and have a large negative impact on aquaculture, ecological environment, and tourism. It is therefore of great significance to study phytoplankton and red tide species in this area. In this review, we examined and compared the composition and relative abundance of phytoplankton reported from the marine surveys carried out between 1957 and 2019 in the Bohai Sea, with a focus on the red tide species. We also analyzed and reviewed major red tide events in Bohai Sea in the past 20 years. Altogether 140 species were identified as red tide species in the Bohai Sea in all marine surveys, which included 78 species in Ochrophyta, 52 species in Dinoflagellata, 5 species in Haptophyta, 4 species in Chlorophyta, and 1 species in Cyanophyta. We also provided detailed descriptions of common red tide species in the Bohai Sea. This review provides a reference for studying biodiversity of phytoplankton and red tide species in the Bohai Sea as well as in other sea areas, and for facilitating the interpretation of metagenomics and macrobarcoding results.
Key words:
Bohai Sea biodiversity marine survey red tide species phytoplankton
海洋中微藻或大型藻类在一定环境条件下快速增长或聚集, 使海水中生物量暴发性增加, 从而导致海水的颜色变化, 这种异常的生态现象通常被称为藻华(俞志明等, 2019)。国际上把造成危害的藻华称为有害藻华(harmful algal blooms, HABs)。我国海域常见的有害藻华包括由微藻或微微藻形成的“赤潮” (red tide)或“褐潮” (brown tide), 由大型绿藻浒苔形成的“绿潮” (green tide), 以及由大型褐藻铜藻形成的“金潮” (golden tide)(于仁成等, 2020)。近年来, 人类活动的急剧增加和全球气候持续变化的双重压力导致赤潮暴发的规模和频率均呈逐步上升趋势, 对海洋生态系统、水产养殖业和旅游观光业等造成负面影响, 这方面的研究已成为环境生态学领域的热点(郭皓等, 2015; 林森杰等, 2019)。
渤海(37°07′-41°0′N, 116°35′-122°15′E)是我国唯一的半封闭型内海(鹿琳, 2012), 总面积为77000 km2, 东西向的宽度约为346 km, 东北至西南的长约555 km, 由北部的辽东湾、西部的渤海湾、南部的莱州湾、中部的中央浅海盆地和东部的渤海海峡五个部分组成(杨琳, 2012) (图 1)。渤海三面环陆, 仅通过口宽约109 km的渤海海峡与黄海相通, 海区平均水深仅18 m, 最大水深也只有83 m (张莹, 2016)。渤海陆源径流丰富, 包括四十余条河流, 如在莱州湾入海的黄河、小清河, 辽东湾的辽河, 渤海湾的海河等, 为渤海带来大量淡水、泥沙以及丰富的营养物质, 导致渤海浮游生物种类繁多, 生物多样性高, 是众多经济鱼虾类的主要产卵场和索饵育肥场, 具有十分重要的生态价值(郭术津等, 2014; 张莹, 2016)。近几十年来, 渤海周边地区城市化发展非常迅速, 然而人口和经济的迅猛增长, 导致城市生活污水和工业废水排放量显著增加(Wu et al, 2013)。由于污染物排放和富营养化等因素, 渤海赤潮暴发频率和强度均呈上升趋势(宋南奇等, 2018)。频繁发生的赤潮灾害严重降低了渤海海域的物种多样性, 打破了生态系统平衡, 也影响了渤海海域的养殖和旅游业, 已经成为渤海最主要的海洋灾害之一(林凤翱等, 2008; 张青田, 2013)。
赤潮已经成为我国沿海地区海洋环境问题中最突出的一类海洋生态灾害问题(周名江等, 2007)。我国针对渤海赤潮物种研究可以追溯到1952年费鸿年对渤海黄河口沿岸水域夜光藻赤潮的研究(费鸿年, 1952)。1958-1959年通过全国海洋综合调查首次对渤海浮游植物的系统分析(徐渡, 2010)。1978-1980年完成的国家重大科研项目第145专题“渤海赤潮的发生机制及预测预报方法研究”, 为我国渤海赤潮的全面和深入研究打下了良好基础。20世纪90年代的“七五”和“九五”赤潮重大项目进一步推动了我国赤潮科学的发展。今年来针对国家的迫切需求和国际赤潮研究的发展趋势启动的赤潮973、赤潮863和赤潮公益专项等使我国的赤潮研究进入新纪元。
随着渤海沿岸经济圈的迅速发展, 大量城市污水和工业废水排入导致了渤海水污染的加重, 富营养化和赤潮问题也日益突出(郭术津等, 2014)。因此, 20世纪末到21世纪初对渤海浮游植物的研究重点也逐步转移到浮游植物种群动力学的预测和赤潮方面(康元德, 1991; 俞建銮等, 1993; 王俊等, 1998; 孙军等, 2004c)。
渤海浮游植物早期研究工作主要采用走航调查, 侧重于针对浮游植物的分类和生态分布习性, 其后通过网采或水样采集, 通过显微镜观察根据浮游植物的形态学特征进行鉴定和计数等方法对渤海海域浮游植物的种类组成和分布情况进行研究(孙军等, 2005; 张秋丰等, 2007; 王毅波等, 2019)。虽然光学显微镜计数法观测面积大且能较全面地反映浮游植物的种类组成信息(Hallegraeff et al, 2003), 但也存在明显的缺点。由于很多浮游植物的细胞粒径较小, 尤其是微微型浮游植物细胞(直径2-3 μm)容易被遗漏, 从而会造成统计数据的不准确, 不能准确分析有害藻华物种在藻华暴发过程中的动态变化(钱奎梅, 2015), 导致不同项目的分析结果之间往往难以相互比较, 不利于比较研究(许歆, 2017)。还有, 有一些藻类细胞比较脆弱, 难以很好地采集和固定, 导致它们在分析中出现偏好和对结果的误读(于杰, 2014)。另外, 基于形态学的分析方法不能准确、充分区分隐存种(cryptic species), 降低了对不同赤潮物种的分辨率。比如, 细胞形态高度相似的塔玛亚历山大藻复合种被利用分子生物方法鉴定为多个具有不同地理分布格局、不同生理适应特征的隐存种, 可以区分为Alexandrium tamarense、Alexandrium catenella、Alexandrium australiense、Alexandrium mediterraneum和Alexandrium pacificum (John et al, 2014)。
分子生物技术的应用大大促进了藻类分类鉴定和系统发生学研究(Ebenezer et al, 2012)。利用分子生物学技术来定性和定量分析浮游植物已经成为一种趋势, 是对传统的浮游植物分析方法的重要补充。近几年来, 基于分子标记扩增和高通量测序的DNA宏条形码(metabarcoding)技术的应用极大地推动了浮游植物研究, 具有准确、快速、高效和高通量等优势(张宛宛等, 2017; 陈楠生, 2020)。Xu等(2017)利用18S rDNA的V4分子标记对秦皇岛近岸海域2013年春季和秋季样品进行宏条形码分析, 鉴别出包括抑食金球藻在内的71种藻, 其中有34种浮游植物在此之前未曾在之前的渤海报道过。Song等(2019b)通过对18S rDNA V4的宏条形码高通量测序分析了2014年渤海辽东湾水域浮游植物群落组成和特征, 共鉴定出93种藻类, 包括56个(约占60%)在中国海域未发现或报告的藻类物种。这些研究补充了秦皇岛近岸海域真核浮游植物, 尤其是微微型浮游植物(< 3 μm)的物种库信息(Song et al, 2019b)。因此, 宏条形码分析可以用于准确鉴定和跟踪包括抑食金球藻等赤潮物种在内的很多微型和微微型藻类物种, 显示了宏条形码技术在研究赤潮物种研究中的重要性, 可以与基于形态观测的分析方法联合使用。本文整理了46个主要渤海航次调查资料, 比较分析了浮游植物组成以及相对丰度, 详细阐述了渤海常见赤潮物种, 可为今后进一步比较研究不同海域浮游植物和赤潮生物物种的多样性, 探索海洋生物多样性的起源和演化规律, 解读宏基因组学及宏条形码分析结果提供基础和参考。
1 渤海环境基本特征渤海海域具有比较复杂和多变的海洋环流系统, 主要包括潮汐余流(tide-induced residual current)、热盐环流(thermohaline circulation)和风生环流(wind-driven circulation), 主要水系包括高盐的黄海暖流余脉(Yellow Sea Warm Current)和低盐的渤海沿岸流(Bohai Coastal Current)(张莹, 2016)(图 1)。余流是一种非周期性的海流, 渤海余流较弱。渤海环流的变化受制于气候条件, 冬季受风的影响较大, 夏季则密度流占主导, 受风影响较小。进入冬季(图 1a), 黑潮支流(The Kuroshio Branch)-北黄海暖流余脉, 通过渤海海峡北部进入渤海, 一直往西, 直到渤海西岸, 遇海岸受阻, 分为南北两个分支, 北支沿渤海西岸进入辽东湾, 构成右旋(顺时针)环流。进入夏季(图 1b)后则流向发生反转, 进入的海流在渤海海峡西北口便进行分支, 一支继续西行, 一支北上, 在辽东湾构成左旋(逆时针)环流。南支进入渤海湾, 与近岸河流相接构成左旋(逆时针)环流, 最后在南部通过渤海海峡流出渤海(万修全, 2003; 毕聪聪, 2013; 韩雅琼等, 2013)。
渤海环境要素呈现出长期变化规律。从1982-2017年, 渤海年均海表温度(sea surface temperature, SST)整体呈现上升趋势, 升温速率为0.255℃/10a, 并且渤海SST年内空间分布, 随着纬度的下降而升高, 由东北向西南递增变化分布, 地表净太阳辐射主要影响了这种分布特征(刘伊格, 2019)。渤海的盐度自1965年以来在整体上呈现持续升高的趋势, 并伴有明显的年际变化。渤海的30 a间的平均盐度升高了约1.2, 年变率为0.04 a–1, 而导致渤海平均盐度升高的主要原因是黄河入海流量持续的锐减(吴德星等, 2004; 马超等, 2010)。叶绿素a浓度是反映浮游植物生物量的重要指标(Blondeau-Patissier et al, 2014)。渤海浮游植物可能受陆源径流营养物质输入影响较大。从1998-2017年渤海全年叶绿素a浓度线性趋势增加显著, 年增长率为0.0522 mg/(m3·a), 增加了14.10%(田洪阵等, 2019)。
渤海受黄河等径流和人类活动的影响非常显著, 营养盐的变化波动较大。近年来渤海海域内营养盐结构失衡, 富营养化严重, 给浮游植物群落结构以及生态结构的稳定带来显著的影响, 驱动了该海域赤潮的形成。渤海中部海域的营养盐结构发生显著变化, 表现为氮(N)/磷(P)比值的升高和硅(Si)/氮(N)比值的下降(于志刚等, 2000)。渤海各个海区内营养盐还具有明显的季节变化, 主要表现为春季向夏季降低秋季快速升高的趋势, 其中辽东湾春季营养盐浓度最高(张海波等, 2019)。渤海由早期的氮限制转化变为磷限制和硅限制(于志刚等, 2000; 魏皓等, 2003; 蒋红等, 2005; 郭术津等, 2014)。这种转化可能与城市污水的排放、农用化肥的流失和黄河断流等多种因素有关(于志刚等, 2000; 郭术津等, 2014; 张海波等, 2019)。
2 渤海的赤潮及其影响渤海属于北温带的半封闭内海。由于受人类活动和陆源径流影响较大, 同时受全球气候变化影响, 渤海赤潮频发(宋南奇等, 2018)。在渤海海域, 辽东湾、莱州湾和渤海湾的赤潮问题最为严重, 历史上曾多次暴发过大规模赤潮灾害, 对渤海的海水养殖业造成了巨大破坏(于仁成等, 2016)。统计表明, 近20年期间渤海发生主要藻华事件多达151次, 累积藻华发生面积超过了36209 km2, 造成3.41亿元直接经济损失。其中由夜光藻引发的赤潮和抑食金球藻引发的褐潮次数最多。根据藻华记录发现, 早期大多数藻华属于单相型藻华, 而近年来多种优势藻同时引起藻华的事件增多。渤海藻华发生最频繁的区域为渤海湾及邻近海域。藻华主要发生在5-11月份。
早期基于显微观察的形态鉴定以及近年来基于分子分析和形态关系相结合的研究逐步解析了渤海海域赤潮的原因种, 包括球形棕囊藻和抑食金球藻。2004年6月在黄河口附近海域暴发球形棕囊藻(Phaeocystis globosa)赤潮, 覆盖面积约1850 km2。其后, 球形棕囊藻赤潮分别又在黄河口(2005年5-6月)、天津近岸海域(2006年10-11月)及河北黄骅附近海域(2007年10月)暴发, 严重影响生态环境和养殖业(屠建波等, 2011; 沈萍萍等, 2018)。自2009年起, 褐潮几乎每年都在渤海秦皇岛近岸海域暴发, 导致养殖区内的扇贝大量死亡。造成了严重经济损失。色素组成分析确认褐潮原因致灾种为含有岩藻黄素的一类海金藻(Kong et al, 2012), 分子标记分析确定发生在秦皇岛近海的褐潮原因致灾种为抑食金球藻(Aureococcus anophagefferens) (Zhang et al, 2012)。对沉积物样本的18S rDNA V9区扩增测序分析分析抑食金球藻可能是在包括渤海在内的全世界范围内广泛分布的赤潮物种(Tang et al, 2019)。Qiao等(2019)通过对渤海海域褐潮样品中微型浮游生物的18S rDNA的V9可变区进行测序分析, 获得了微型浮游生物的结构组成和生物多样性, 特别在水样中发现褐潮藻抑食金球藻和多形微眼藻(Minutocellus polymorphus)的大量存在。通过扩增测序18S rDNA V4测序区在辽东湾发现了隶属甲藻纲甲球球藻科的Stoeckeria algicida (宋伦等, 2017a)。这个曾在韩国海域引发赤潮的物种可以侵噬鱼类细胞引起鱼类死亡(Jeong et al, 2005)。这些研究结果表明渤海海域还有很多浮游植物和赤潮物种等待鉴定, 基于分子生物学的分析方法与基于形态学的分析方法联合使用可以逐步完成鉴定。
3 渤海浮游植物调查文献检索发现1957-2019年间渤海海域针对浮游植物的46个航次调查结果报告(附表 1)。这些渤海海域的调查涉及的调查范围分布较广, 包括渤海中部海域、渤海湾西北海域、莱州湾靠近黄河口海域、河北省秦皇岛近岸海域和辽东湾东北海域(图 2)。
 |
| 图 2 渤海航次站位的分布范围 Fig. 2 Sampling stationsin the marine surveys in the Bohai Sea |
在迄今报道的46个渤海航次调查中, 共鉴定出包括6个门的380个浮游植物物种(附表 2), 其中硅藻门(Bacillariophyta)藻种数量最多共244个, 其次是甲藻门(Dinoflagellata)共89个, 绿藻门(Chlorophyta)共22个, 定鞭藻门(Haptophyta)共13个, 隐藻门(Cryptophyta)共10个和蓝藻门(Cyanophyta)2个。历次渤海航次报导的380个浮游植物种中, 在NCBI数据库中只查到了212个(55.8%)物种的全长18S rDNA序列(>1600 bp), 说明数据库中的全长18S rDNA序列数据还很不全面, 也说明渤海水域中的很多浮游植物尚未得到系统的分子分析。利用数据库中查找到的全长18S rDNA序列构建系统进化树(图 3a)发现渤海水域的浮游植物聚类到5个门, 与航次鉴定的结果基本相符。其中以硅藻物种种类最多(119个), 其次是甲藻(51个)。在渤海水域历年航次调查中出现频率最高的优势浮游植物包括中肋骨条藻(Skeletonema costatum)、具槽帕拉藻(Paralia sulcata)、星脐圆筛藻(Coscinodiscus asteromphalus)、夜光藻(Noctiluca scintillans)、虹彩圆筛藻(Coscinodiscus oculusiridis)、尖刺伪菱形藻(Pseudo-nitzschia pungens)、尖刺菱形藻(Nitzschia pungens)、格氏圆筛藻(Coscinodiscus granii)、浮动弯角藻(Eucampia zodiacus)和抑食金球藻(图 3b, 3c)。
 |
| 图 3 渤海浮游植物物种组成 Fig. 3 The composition of phytoplankton in the Bohai Sea 注: a. 渤海历史文献中报道的浮游植物18S rDNA序列(>1600 bp)构建的系统进化树(ML), 红色代表硅藻, 蓝色代表甲藻; b. 渤海历史文献中报道的优势浮游植物(出现频次>1)词云图; c. 长江口历史文献中报道的优势浮游植物(出现频次>1)柱状图 |
不同航次鉴定报道的浮游植物物种数目相差较大, 不同航次报道的物种组成相似性也很低, 没有任何浮游植物物种是46个航次中都报道过的(图 4a)。宏条形码分析能发现更多的特有物种(宋伦等, 2016; Xu et al, 2017; Song et al, 2019a)。造成不同航次调查结果之间的差异的原因很多, 包括不同航次可能调查的目的不同(比如有的航次以甲藻为主要调查对象), 航次的季节不同, 站位数的设置不同等。
 |
| 图 4 渤海浮游植物组成的航次调查结果比较分析 Fig. 4 Comparative analysis of phytoplankton identified in the marine surveys in the Bohai Sea 注: a. 航次调查结果统计到的浮游植物物种(出现频次≥2)Venn图; b. 历史航次统计的浮游植物中硅藻和甲藻的比例变化 |
文献报道表明, 浮游植物群落经历了由硅藻绝对控制转向硅藻和甲藻共同控制, 渤海的硅藻组成逐步下降, 而甲藻组成逐步上升(孙军等, 2002, 2005; 郭术津等, 2014; 孙雪梅等, 2016; 栾青杉等, 2018)。为了进一步评估渤海海域浮游植物种类组成的变化趋势, 我们系统整理了渤海海域46个航次中每个航次报道的甲藻和硅藻数目, 对文献里报道的甲藻和硅藻的实际数据进行逐年比较分析。我们的分析结果并没有显示硅藻组成逐步下降和甲藻组成逐步上升趋势(图 4b)。值得强调的是, 由于这些航次的季节、采样方法、采样位点、物种的分类标准和分析手段都可能不同, 都对分析结果会产生影响。除此之外, 主观原因比如人为因素(比如不同研究人员的分类能力)也可能导致对物种的判断不同。因此, 上面的分析结果可能不全面。随着基于分子标记(比较宏条形码分析)等定量分析的广泛使用(陈楠生, 2020), 渤海海域浮游植物组成应该能够得到更加准确的研究和判断。
4 渤海常见赤潮物种1957-2019年间的46个渤海航次调查共鉴定了包括5个门的140个赤潮物种(附表 3), 包括硅藻78种、甲藻52种、定鞭藻5种、绿藻4种和蓝藻1种。航次调查中出现频率较高(≥7), 比较常见的赤潮物种有48个(表 1)。不同航次报道的赤潮物种的种类和数目差异均较大, 并且没有一个赤潮物种在所有航次中都被发现(图 5a), 也表明渤海海域生态系统的多样性和复杂时空变化。根据文献综述, 渤海海域最常见的10个赤潮优势物种包括夜光藻、中肋骨条藻、星脐圆筛藻、具槽帕拉藻、抑食金球藻、尖刺菱形藻、格氏圆筛藻、尖刺拟菱形藻、三角角藻(Ceratium fusus同Tripos fusus)和叉状角藻(Ceratium furca同Tripos furca)(图 5b, 5c)。
表 1 渤海历史航次赤潮物种名录 Tab. 1 Common red tide species identified in the marine surveys in the Bohai Sea
 |
| 图 5 渤海赤潮物种组成(出现频次>1) Fig. 5 The composition of red tide species in the Bohai Sea 注: a. 各历史航次统计到的赤潮物种Venn图; b. 渤海历史文献中报道的优势赤潮物种词云图; c. 渤海历史文献中报道的优势赤潮物种柱状图 |
夜光藻是一种较大型的单细胞甲藻, 细胞形态近于球形, 直径约为200 μm-2.0 mm。夜光藻在我国海域分布广泛, 具有很强的环境适应能力, 在水温7.4-28.6℃, 盐度14.8-32.9的范围内都能生长, 是一种耐温耐盐上限较高的赤潮生物(赵冬至, 2000)。吴玉霖等(1994)报道的食性实验研究结果表明, 夜光藻对不同微藻具有不同的摄食强度(扁藻>等鞭金藻>三角褐指藻)。夜光藻不仅摄食浮游藻类, 还摄食其他有机碎屑和小型浮游动物等, 摄食活动和营养繁殖具有昼夜节律性(陈汉辉等, 1991; 吴玉霖等, 1994; 周成旭等, 1994)。在渤海海域, 夜光藻主要分布于渤海湾(王瑜等, 2016)、莱州湾(张继民等, 2010)、辽东湾(Song et al, 2019a)等水域, 而辽东湾是渤海夜光藻赤潮的高发海域(周遵春等, 2002)。在不同的季节里, 夜光藻也常以浮游植物的优势类群出现(王俊, 2003; 苑明莉等, 2014; 孙雪梅等, 2016; 张彭如雁等, 2016)。近20年间, 渤海海域经常发生以夜光藻为优势种的赤潮, 其中, 2005年6月16-18日发生在辽宁营口鲅鱼圈附近海域大面积的夜光藻赤潮, 影响面积多达2000 km2, 主要受灾种类包括鱼、虾、贝、藻类, 造成了严重的经济损失。
4.2 中肋骨条藻中肋骨条藻是世界性广布型海洋硅藻(Tada et al, 2001)。细胞体透镜形或圆柱形, 直径为6-22 μm, 壳面圆而鼓起, 着生一圈细长的刺, 与相临细胞的对应刺组成长链(陈锤, 2005)。从春季到秋季, 中肋骨条藻可在高温(25℃)和高盐度(18-25)下大量繁殖, 是引起中国沿海水域有害赤潮的关键物种(Yan et al, 2002; Liu et al, 2005)。中肋骨条藻也被认为是大多数富营养化水域赤潮的主要组成部分(霍文毅等, 2001)。虽然中肋骨条藻细胞个体比较小, 但其数量比较多, 生物量较高, 对整个渤海生态系统的影响较大(尹翠玲等, 2013)。中肋骨条藻作为优势种在渤海海域共出现过17次, 主要分布于渤海湾(杨世民等, 2007; 徐玉山等, 2009)、莱州湾(冷宇等, 2013)、辽东湾(孙萍等, 2008)、渤海中部(孙雪梅等, 2016)等水域。2013年7月5-8日在天津市临港经济区东部海域发生中肋骨条藻赤潮, 赤潮影响面积超过154 km2。虽然中肋骨条藻不产生毒素, 但会繁殖产生大量的生物量, 对水的物理和化学特性都有不利影响, 并可引起海水变色并产生反应性醛, 对桡足类的孵化有负面影响(Miralto et al, 1999)。中肋骨条藻赤潮严重影响海洋生物, 威胁到生态系统的稳定性(王朝晖等, 2010), 也给水产养殖业带来巨大的经济损失(Ou et al, 2008)。
4.3 星脐圆筛藻星脐圆筛藻属于硅藻, 细胞单体, 圆盘状, 壳环轴长20-35 μm, 生态类型属于广温种, 广泛的分布于我国各海域(栾青杉等, 2018)。星脐圆筛藻是渤海辽东湾海区中毛虾的主要饵料, 并且星脐圆筛藻在渤海海域全年都能作为优势种出现(张秋丰等, 2007; 杨阳等, 2016)。星脐圆筛藻物种属于假性浮游物种, 个体体积较大, 而大体积藻细胞具有较高的碳含量, 对整个碳库影响较大, 在海洋生态系统中能够起到关键性的作用(孙军等, 2002, 2004c)。星脐圆筛藻群落对胶州湾浮游植物生物量的贡献极其显著(Shen et al, 2017)。渤海近20年间未发生星脐圆筛藻赤潮现象, 但由于近年该藻在渤海出现的频率愈来愈高, 在渤海湾(王瑜等, 2016)和天津近岸(曹春晖等, 2006; 张彭如雁等, 2016; 张雪等, 2018)等海区暴发赤潮的可能性很高。
4.4 具槽帕拉藻具槽帕拉藻属于硅藻门, 直链藻属, 同种异名: 具槽直链藻(Melosira sulcata)。细胞短圆柱, 通常10-20个相连形成直链, 两细胞间有凹纹。细胞直径一般为12-40 μm, 直径大于壳环轴, 高5-16 μm。具槽帕拉藻生态类型属于温带近岸种。具槽帕拉藻的最适生长条件是温度为25℃, 光照强度为3000 lx, 盐度为24, 并且在温度20℃、光照4500 lx、盐度32的培养条件下最有利于孢子的生成(朱明等, 2007)。随着水温的变化, 渤海海域具槽帕拉藻群落的优势度也会随之发生改变(张彭如雁等, 2016)。虽然近20年来渤海未发生具槽帕拉藻赤潮现象, 但是具槽帕拉藻近年在渤海海域出现的频率和数量愈来愈高(孙军等, 2004b; 杨世民等, 2007; 孙萍等, 2008; 苑明莉等, 2014; 郭术津等, 2014; 孙雪梅等, 2016; 王毅波等, 2019), 值得关注。
4.5 抑食金球藻抑食金球藻属于海金藻纲(Pelagophyceae), 是一种个体微小(2-3 μm), 没有运动能力, 球形的微微型浮游藻类。抑食金球藻可以在温度范围内(0-25℃)生长, 但最适合生长的温度约为20℃ (Gobler et al, 2002)。抑食金球藻通常在降雨量较少和盐度较高(≥24)的海湾暴发褐潮(Laroche et al, 1997)。研究显示硝酸盐和氨氮浓度在抑食金球藻暴发之前明显降低, 无机氮和无机磷浓度在暴发时也非常低(古彬等, 2015)。研究表明抑食金球藻在DON和DIN共同作为氮源的环境中更能适应低光环境(Pustizzi et al, 2004)。色素分析和分子分析确定渤海褐潮的原因种为抑食金球藻(Kong et al, 2012; Zhang et al, 2012)。我国是继美国和南非之后第三个暴发抑食金球藻褐潮的国家(于杰, 2014)。抑食金球藻暴发褐潮时海水呈褐色, 持续时间久, 影响面积广, 褐潮优势藻浓度可高达109 ind./L (于杰, 2014; 乔玲等, 2016)。抑食金球藻暴发褐潮期间养殖的贝类摄食受到抑制, 使海湾扇贝出现滞长和部分死亡, 不仅使海洋生态系统遭到破坏, 还给旅游业和水产养殖业造成巨大的经济损失(宋伦等, 2016, 2017b; Xu et al, 2017; Chen et al, 2019; Song et al, 2019a)。自2009年, 渤海共发生4次较大规模由抑食金球藻为主要优势种的褐潮。其中, 2012年6-8月, 在河北省唐山市至辽宁省绥中近岸海域, 发生了约为3400 km2超大面积的褐潮。12个航次调查中抑食金球藻作为优势物种出现, 包括秦皇岛近岸海域的调查(Zhang et al, 2012; 于杰等, 2015; Xu et al, 2017; Chen et al, 2019; Qiao et al, 2019; Yao et al, 2019)及辽东湾海域的调查(宋伦等, 2016)。
4.6 尖刺菱形藻尖刺菱形藻属于硅藻门, 菱形藻属。细胞细长, 呈梭形, 末端尖。长80-134 μm, 宽3.7-9.0 μm。生态类型属于广温性近岸种, 分布于世界各地, 如美洲和欧洲等, 我国近海也均有报道。尖刺菱形藻是硅藻类中能引起毒性(健忘性贝毒素)赤潮的最常见的一种赤潮生物(赵冬至, 2000)。邹景忠等在国内首次研究了尖刺菱形藻在我国近海的分布及其与环境因子的关系等毒素生态特征(Zou et al, 1993)。尖刺菱形藻群落发展与盐度的变化是密切相关的(曹春晖等, 2006)。近20年间, 渤海未发生尖刺菱形藻赤潮现象, 但是尖刺菱形藻在多个渤海航次中被鉴定为优势物种, 包括渤海湾(张秋丰等, 2007)、莱州湾(张继民等, 2010)和天津近岸等航次调查(曹春晖等, 2006; 张彭如雁等, 2016; 张雪等, 2018), 表明该物种具有较高的丰度, 有可能暴发赤潮。
4.7 格氏圆筛藻格氏圆筛藻又称偏环圆筛藻。藻体细胞直径60-215 μm。属于世界广布性藻类, 我国各海域均有分布, 其中以黄海和渤海数量较多(张秋丰等, 2007)。格氏圆筛藻是渤海毛虾和挠足类、糠虾等的主要饵料, 圆筛藻的种群数量变动和分布与毛虾的摄食强度密切相关(康元德, 1991; 王俊等, 1998)。格氏圆筛藻与硝酸盐、亚硝酸盐和氨氮具有显著正相关性(杨阳等, 2016)。周然等通过渤海湾样品的RDA分析表明, 格氏圆筛藻与水温成正相关, 喜欢较高温度的水域, 虽然这与认为硅藻一般喜欢较低温水域的结论不一样(da Silva et al, 2005), 但这与格氏圆筛藻生态习性相关, 属于广温性藻类, 可以在不同水温下生长(周然等, 2013)。格氏圆筛藻为假性浮游物种, 细胞个体较大, 碳含量较高, 在渤海生态系统中起到关键性的作用, 因此也被列为渤海浮游植物的关键种(孙军等, 1999; 张秋丰等, 2007; 尹翠玲等, 2013)。格氏圆筛藻作为优势赤潮物种在渤海海域共出现在了9个航次中, 包括渤海湾(张秋丰等, 2007; 徐玉山等, 2009; 周然等, 2013)和天津近岸海域等航次调查(张彭如雁等, 2016; 张雪等, 2018)。近20年间, 渤海未发生格氏圆筛藻赤潮现象, 但在不同的季节里, 该藻也常以浮游植物的优势类群出现(尹翠玲等, 2013), 暴发赤潮的潜在性大, 今后的工作中应将格氏圆筛藻列为渤海重点关注的浮游植物关键种。
4.8 尖刺拟菱形藻尖刺拟菱形藻是一种在全球近岸海域分布极广能够产毒的硅藻, 属于广温广盐物种, 也是我国沿海许多海域的优势种(吕颂辉等, 1992)。尖刺拟菱形藻细胞为线形或披针形, 环面呈纺锤状, 群体具有运动能力。我国学者对东海及我国沿海的尖刺拟菱形藻进行了毒素检测, 但并没有检测到多莫酸(domoic acid)(董焕嫦等, 2018)。尖刺拟菱形藻对氮和磷的依赖性较强, 属于营养依赖型藻类, 最适生长的氮磷比在10-32之间(吕颂辉等, 1992)。渤海湾浮游植物中的尖刺拟菱形藻长期属于优势种, 在渤海湾生态系统中起着关键性的作用, 因此尖刺拟菱形藻被列为渤海湾本地种中的关键种(杨世民等, 2007)。渤海海域的尖刺拟菱形藻与水温、活性磷酸盐、硅酸盐、亚硝酸盐和氨氮具有良好的正相关性(卞少伟等, 2015)。近20年间, 渤海未发生以尖刺拟菱形藻为优势种的赤潮现象, 但在不同的季节里, 经常以优势种类群出现(孙军等, 2005; 张彭如雁等, 2016), 潜在暴发赤潮的机会大, 值得关注。尖刺拟菱形藻作为优势赤潮物种在渤海航次调查中共出现过8次, 主要包括渤海湾(杨世民等, 2007)、辽东湾(孙萍等, 2008)、渤海中部(孙军等, 2005)、天津近岸(卞少伟等, 2015; 张彭如雁等, 2016; 张雪等, 2018)和渤海海峡(王毅波等, 2019)等海域航次调查。
4.9 三角角藻三角角藻属于单细胞甲藻, 有时几个细胞会连成链状。生态类型属广温广盐的世界性种, 三大洋均有分布。三角角藻会随着黑潮进入渤海水域并影响渤海浮游植物群落结构, 属于暖海大洋性的外源性物种, 出现频度和优势度都非常高, 极易形成赤潮并造成灾害(孙军等, 2005)。三角角藻在渤海的繁殖盛期一般出现在夏季和秋季, 往往在8月会成为该海域浮游植物的优势种, 在冬季尽管三角角藻的优势度很低, 但经常出现, 出现频率达到52%, 后续研究值得关注(俞建銮等, 1993; 孙军等, 2004b)。
三角角藻作为优势藻种, 分布极不均匀, 最高细胞丰度值出现在莱州湾湾口(苑明莉等, 2014)。与历史资料相比, 渤海湾近岸海域的浮游植物群落中硅藻仍占有较大优势, 但甲藻门占比有所上升, 特别是赤潮藻三角角藻等的数量有所增加, 在秋季出现频率达到93.33% (王瑜等, 2016)。近20年间, 渤海未发生三角角藻赤潮现象, 但在不同的季节里, 该藻也常以浮游植物的优势类群出现, 暴发三角角藻赤潮的潜在性大, 后续需密切监视。三角角藻作为优势赤潮物种在渤海海域航次共出现过7次, 主要出现在渤海湾(张秋丰等, 2007; 王瑜等, 2016)、渤海中部水域(孙军等, 2005)和莱州湾(苑明莉等, 2014)等水域调查航次。
4.10 叉状角藻叉状角藻, 多甲藻目, 角藻科, 为温带偏冷水种, 分布较广, 数量多, 适宜生长温度为6.6-29.4℃, 具有运动和捕食能力, 经常在内湾水域形成赤潮(周遵春等, 2002)。在对1998年秋季渤海中部和渤海海峡及其邻近海域进行浮游植物群落调查期间, 在渤海湾发现一次梭状角藻和叉状角藻的混合型赤潮(孙军等, 2004a)。2006年渤海湾赤潮重点监控区的监测结果显示, 在浮游植物群落中, 硅藻为主要优势种, 而甲藻门中叉状角藻在个别站位也成为了优势种(张秋丰等, 2007)。曹春晖等对天津近岸海域调查也表明, 调查区的浮游植物优势种大部分为硅藻, 但叉状角藻在个别站位形成优势(曹春晖等, 2006)。2008-2012年渤海浮游植物优势种多为硅藻, 但叉状角藻在2008年浮游植物群落中优势度达0.640, 优势明显极易引发有害甲藻赤潮, 后续应对其进行重点关注(尹翠玲等, 2013)。近20年间, 渤海发生2次叉状角藻赤潮现象, 其中, 2014年8月26日-9月25日, 在天津滨海旅游区海域发生了离心列海链藻, 多环旋沟藻和叉状角藻的多种优势种混合型的赤潮, 影响面积为300 km2。叉状角藻作为优势赤潮物种在渤海海域调查中共出现过7次, 包括渤海湾(张秋丰等, 2007)、渤海中部(孙军等, 2004a)和天津近岸(曹春晖等, 2006)等海域调查航次。
5 主要结论与展望本文通过对渤海历史文献资料的整理汇总, 主要得到如下几点结论:
(1) 在我国, 渤海是除东海长江口邻近海域和南海近岸海域之外的典型的赤潮高发区。通过分析整理《中国海洋灾害公报》中报导的在渤海水域暴发的赤潮事件, 我们发现近20年期间渤海发生主要藻华事件多达151次, 累积藻华发生面积超过了36209 km2, 造成很高的经济损失。其中由夜光藻引发的赤潮和抑食金球藻引发的褐潮次数最多。渤海藻华发生最频繁的区域为渤海湾及邻近海域。
(2) 渤海由于特殊的地理位置, 具有很高的环境异质性, 浮游植物的生物多样性也很高, 其中硅藻种类物种最多, 其次是甲藻。在渤海海域所有浮游植物中, 中肋骨条藻是文献报道中出现次数最多的优势物种, 其次是具槽帕拉藻、星脐圆筛藻和夜光藻。大多数报道认为在近几十年里, 渤海海域的硅藻组成在逐步下降, 而甲藻组成逐步上升。不过, 我们通过对相关文献报导的结果进行比较分析, 并没有发现浮游植物组成中硅藻和甲藻的相对组成显示出显著的变化。
(3) 渤海赤潮物种具有较高的生物多样性。通过整理统计了1957-2019年间46个渤海航次调查数据, 我们发现了包括5个门的140个赤潮物种, 包括硅藻78种, 甲藻52种, 定鞭藻5种, 绿藻4种和蓝藻1种。
(4) 渤海浮游植物和赤潮物种都很丰富, 但是目前代表性分子标记(比如18S rDNA, 28S rDNA, ITS, cox1, rbcL)数据库中代表渤海浮游植物和赤潮物种的数据有限, 加上每个物种基因拷贝数可能不同, 比如一些甲藻18S rDNA具有极高的拷贝数, 不同拷贝之间还存在序列差异(胡晓坤等, 2019), 这些都严重限制了组学分析方法的充分利用。需要对渤海浮游植物特别是赤潮物种进行系统性的分析, 并获得重要分子标记(比如18S rDNA)序列信息和拷贝数信息和拷贝序列信息, 丰富数据库, 促进宏条形码分析在研究渤海赤潮中的利用。
今后的工作中, 我们应更加关注渤海浮游植物的物种组成和特征, 尤其是赤潮物种, 通过采用具有高分辨率和高特异性的分子生物学方法, 并结合传统的镜检方法, 对数据进行深入和准确的分析, 这不仅有助于我们掌握渤海赤潮物种的变化规律, 而且能够对渤海赤潮暴发作出预测, 及时防范赤潮风险, 降低由赤潮带来的经济损失。
电子附件材料:
附表 1、附表 2、附表 3见http://dx.doi.org/10.11693/hyhz20200900245
参考文献
于杰, 2014. 浮游生物多样性高效检测技术的建立及其在渤海褐潮研究中的应用. 青岛: 中国海洋大学硕士学位论文, 1-64
|
于杰, 张玲玲, 孙妍, 等. 2015. 渤海海域褐潮期微型浮游生物多样性的初步研究. 中国海洋大学学报, 45(3): 73-78 |
于仁成, 吕颂辉, 齐雨藻, 等. 2020. 中国近海有害藻华研究现状与展望. 海洋与湖沼, 51(4): 768-788 |
于仁成, 刘东艳. 2016. 我国近海藻华灾害现状、演变趋势与应对策略. 中国科学院院刊, 31(10): 1167-1174 |
于志刚, 米铁柱, 谢宝东, 等. 2000. 二十年来渤海生态环境参数的演化和相互关系. 海洋环境科学, 19(1): 16-19 |
万修全, 2003. 渤海冬夏季环流特征及变异的初步研究. 青岛: 中国海洋大学硕士学位论文, 3-101
|
马超, 鞠霞, 吴德星, 等. 2010. 黄、渤海断面及海洋站的盐度分布特征与变化趋势. 海洋科学, 34(9): 70-75 |
王俊. 2003. 渤海近岸浮游植物种类组成及其数量变动的研究. 海洋水产研究, 24(4): 44-50 |
王俊, 康元德. 1998. 渤海浮游植物种群动态的研究. 海洋水产研究, 19(1): 43-52 |
王艳, 黄琳, 顾海峰, 等. 2012. 中国渤海海域甲藻孢囊的种类多样性和生态地理分布. 植物学报, 47(2): 125-132 |
王瑜, 刘录三, 朱延忠, 等. 2016. 渤海湾近岸海域春秋季网采浮游植物群落特征. 海洋环境科学, 35(4): 565-570 |
王朝晖, 袁美玲, 骆育敏, 等. 2010. 海洋卡盾藻与中肋骨条藻和锥状斯氏藻种间竞争研究. 水生生物学报, 34(2): 336-344 |
王毅波, 孙延瑜, 王彩霞, 等. 2019. 夏季渤海网采浮游植物群落和叶绿素a分布特征及其对渔业资源的影响. 渔业科学进展, 40(5): 42-51 |
卞少伟, 孙韧, 梅鹏蔚, 等. 2015. 2013年春夏季天津近岸海域浮游植物的群落结构. 水生态学杂志, 36(1): 47-52 |
尹翠玲, 张秋丰, 崔健, 等. 2013. 2008-2012年渤海湾天津近岸海域夏季浮游植物组成. 海洋科学进展, 31(4): 527-537 DOI:10.3969/j.issn.1671-6647.2013.04.011 |
古彬, 甄毓, 米铁柱. 2015. 秦皇岛扇贝养殖区褐潮暴发期浮游微藻群落结构特点研究. 中国海洋大学学报, 45(7): 64-72 |
田洪阵, 刘沁萍, Goes J I, 等. 2019. 近20年渤海叶绿素a浓度时空变化. 海洋学报, 41(8): 131-140 |
毕聪聪, 2013. 渤海环流季节变化及机制分析研究. 青岛: 中国海洋大学硕士学位论文, 1-63
|
吕颂辉, 齐雨藻. 1992. 南海大鹏湾的主要赤潮生物. 暨南大学学报(自然科学), 13(3): 130-133 |
朱明, 阎斌伦, 2007. 具槽直链藻生长条件和生活史的研究. 见: 中国海洋湖沼学会藻类学分会第七届会员大会暨第十四次学术讨论会论文摘要集. 呼和浩特: 中国海洋湖沼学会
|
朱树屏, 郭玉潔. 1957. 烟台、威海鲐鱼渔场及其附近海区角毛硅藻属的研究Ⅰ. 分类的研究. 海洋与湖沼, 1(1): 27-87 |
乔玲, 甄毓, 米铁柱. 2016. 抑食金球藻(Aureococcus anophagefferens)褐潮研究概述. 海洋环境科学, 35(3): 473-480 |
刘伊格, 2019. 渤海和黄海海表温度变化及成因分析. 南京: 南京大学硕士学位论文, 1-48
|
刘晓彤, 2011. 夏、秋季黄河口及其邻近海域浮游植物群落结构和粒级结构的研究. 青岛: 中国海洋大学硕士学位论文, 1-295
|
许歆, 2017. 秦皇岛近海浮游植物群落结构变化及其组学研究. 青岛: 中国科学院大学博士学位论文, 1-143
|
孙军, 刘东艳. 2005. 2000年秋季渤海的网采浮游植物群落. 海洋学报, 27(3): 124-132 DOI:10.3321/j.issn:0253-4193.2005.03.018 |
孙军, 刘东艳, 王威, 等. 2004a. 1998年秋季渤海中部及其邻近海域的网采浮游植物群落. 生态学报, 24(8): 1643-1655 |
孙军, 刘东艳, 白洁, 等. 2004b. 2001年冬季渤海的浮游植物群落结构特征. 中国海洋大学学报, 34(3): 413-422 |
孙军, 刘东艳, 杨世民, 等. 2002. 渤海中部和渤海海峡及邻近海域浮游植物群落结构的初步研究. 海洋与湖沼, 33(5): 461-471 DOI:10.3321/j.issn:0029-814X.2002.05.002 |
孙军, 刘东艳, 钱树本. 1999. 浮游植物生物量研究Ⅰ.浮游植物生物量细胞体积转化法. 海洋学报, 21(2): 75-85 DOI:10.3321/j.issn:0253-4193.1999.02.010 |
孙军, 刘东艳, 徐俊, 等. 2004c. 1999年春季渤海中部及其邻近海域的网采浮游植物群落. 生态学报, 24(9): 2003-2016 |
孙萍, 李瑞香, 李艳, 等. 2008. 2005年夏末渤海网采浮游植物群落结构. 海洋科学进展, 26(3): 354-363 DOI:10.3969/j.issn.1671-6647.2008.03.011 |
孙雪梅, 徐东会, 夏斌, 等. 2016. 渤海中部网采浮游植物种类组成和季节变化. 渔业科学进展, 37(4): 19-27 |
纪昱彤, 王宁, 陈洪举, 等. 2018. 2013年秋季渤黄海浮游植物的群落特征. 中国海洋大学学报, 48(S2): 31-41 |
杨阳, 孙军, 关翔宇, 等. 2016. 渤海网采浮游植物群集的季节变化. 海洋通报, 35(2): 121-131 |
杨琳, 2012. 渤海微微型浮游植物的时空分布. 青岛: 中国海洋大学硕士学位论文, 1-74
|
杨世民, 董树刚, 李锋, 等. 2007. 渤海湾海域生态环境的研究Ⅰ.浮游植物种类组成和数量变化. 海洋环境科学, 26(5): 442-445 DOI:10.3969/j.issn.1007-6336.2007.05.010 |
吴玉霖, 周成旭, 张永山. 1994. 夜光藻的室内培养. 海洋与湖沼, 25(2): 165-167 DOI:10.3321/j.issn:0029-814X.1994.02.009 |
吴德星, 万修全, 鲍献文, 等. 2004. 渤海1958年和2000年夏季温盐场及环流结构的比较. 科学通报, 49(3): 287-292 DOI:10.3321/j.issn:0023-074X.2004.03.017 |
冷宇, 赵升, 刘霜, 等. 2013. 黄河口海域夏季浮游植物的分布特征. 水生态学杂志, 34(6): 41-46 DOI:10.3969/j.issn.1674-3075.2013.06.007 |
冷春梅, 曹振杰, 张金路, 等. 2014. 黄河口浮游生物群落结构特征及环境质量评价. 海洋环境科学, 33(3): 360-365 |
沈萍萍, 齐雨藻, 欧林坚. 2018. 中国沿海球形棕囊藻(Phaeocystis globosa)的分类、分布及其藻华. 海洋科学, 42(10): 146-162 DOI:10.11759/hykx20171225004 |
宋伦, 刘卫东, 吴景, 等. 2017a. 有害甲藻Stoeckeria algicida在辽东湾的时空分布. 生态学报, 37(4): 1339-1345 |
宋伦, 吴景, 刘卫东, 等. 2016. 渤海长兴岛海域微型和微微型浮游植物多样性. 环境科学研究, 29(11): 1635-1642 |
宋伦, 吴景, 宋永刚, 等. 2017b. 褐潮致灾种抑食金球藻在辽东湾的分布. 环境科学研究, 30(4): 537-544 |
宋南奇, 王诺, 吴暖, 等. 2018. 基于GIS的我国渤海1952~2016年赤潮时空分布. 中国环境科学, 38(3): 1142-1148 DOI:10.3969/j.issn.1000-6923.2018.03.042 |
张莹, 2016. 渤海中部浮游植物与环境因子的空间关系及季节差异分析. 烟台: 中国科学院大学烟台海岸带研究所硕士学位论文, 1-59
|
张雪, 徐晓甫, 戴媛媛, 等. 2018. 天津近岸人工鱼礁海域浮游植物群落及其变化特征. 渔业科学进展, 39(6): 1-10 |
张青田. 2013. 中国海域赤潮发生趋势的年际变化. 中国环境监测, 29(5): 98-102 DOI:10.3969/j.issn.1002-6002.2013.05.019 |
张宛宛, 谢玉为, 杨江华, 等. 2017. DNA宏条形码(metabarcoding)技术在浮游植物群落监测研究中的应用. 生态毒理学报, 12(1): 15-24 |
张秋丰, 尹翠玲, 徐玉山, 等. 2007. 2006年夏季渤海湾赤潮重点监控区的网采浮游植物群落. 天津科技大学学报, 22(3): 19-23 DOI:10.3969/j.issn.1672-6510.2007.03.006 |
张海波, 王丽莎, 石晓勇, 等. 2019. 渤海中部营养盐季节变化及限制统计分析. 中国环境科学, 39(4): 1579-1588 DOI:10.3969/j.issn.1000-6923.2019.04.028 |
张继民, 刘霜, 张琦, 等. 2010. 黄河口附近海域浮游植物种群变化. 海洋环境科学, 29(6): 834-837 DOI:10.3969/j.issn.1007-6336.2010.06.013 |
张彭如雁, 李德亮, 戴媛媛, 等. 2016. 天津近岸海域浮游植物群落结构特征. 海洋湖沼通报, (6): 53-59 |
陈锤. 2005. 主要水产经济生物开发技术手册. 北京: 中国农业出版社
|
陈汉辉, 杞桑. 1991. 夜光藻摄食和营养繁殖的昼夜节律. 暨南大学学报, 12(3): 104-107, 111 |
陈楠生. 2020. 有害藻华的宏条形码分析: 机会与挑战. 海洋科学, 44(7): 116-134 |
苑明莉, 孙军, 翟惟东. 2014. 2012年秋季渤海和北黄海浮游植物群落. 天津科技大学学报, 29(6): 56-64 |
林凤翱, 卢兴旺, 洛昊, 等. 2008. 渤海赤潮的历史、现状及其特点. 海洋环境科学, 27(S2): 1-5 |
林森杰, 姬南京, 罗昊. 2019. 海洋有害藻华研究进展. 海洋与湖沼, 50(3): 495-510 |
周然, 彭士涛, 覃雪波, 等. 2013. 渤海湾浮游植物与环境因子关系的多元分析. 环境科学, 34(3): 864-873 |
周成旭, 吴玉霖, 邹景忠. 1994. 夜光藻的营养动力. 海洋与湖沼, 25(2): 152-157 DOI:10.3321/j.issn:0029-814X.1994.02.006 |
周名江, 于仁成. 2007. 有害赤潮的形成机制、危害效应与防治对策. 自然杂志, 29(2): 72-77 DOI:10.3969/j.issn.0253-9608.2007.02.003 |
周遵春, 马志强, 薛克, 等. 2002. 对辽东湾夜光藻赤潮和叉状角藻赤潮成因的初步研究. 水产科学, 21(2): 9-12 DOI:10.3969/j.issn.1003-1111.2002.02.003 |
赵文, 魏洪祥, 郭凯. 2011. 2006-2007年大连大窑湾海区浮游植物的群落结构及其季节变化. 大连海洋大学学报, 26(4): 291-298 DOI:10.3969/j.issn.1000-9957.2011.04.002 |
赵冬至. 2000. 渤海赤潮灾害监测与评估研究文集. 北京: 海洋出版社, 52-66
|
胡晓坤, 张清春, 陈振帆, 等. 2019. 北部湾海域球形棕囊藻遗传多样性分析. 海洋与湖沼, 50(3): 601-610 |
俞志明, 陈楠生. 2019. 国内外赤潮的发展趋势与研究热点. 海洋与湖沼, 50(3): 474-486 |
俞建銮, 李瑞香. 1993. 渤海、黄海浮游植物生态的研究. 黄渤海海洋, 11(3): 52-59 |
费鸿年. 1952. 发生赤潮的原因. 学艺, 22(1): 1-3 |
钱奎梅, 刘霞, 陈宇炜. 2015. 淡水浮游植物计数与定量方法. 湖泊科学, 27(5): 767-775 |
徐渡. 2010. 1958~1960年: 全国海洋综合调查. 海洋科学, 34(4): 109-110 |
徐玉山, 刘宪斌, 张秋丰. 2009. 渤海湾近岸海域浮游植物多样性研究. 盐业与化工, 38(6): 11-14 |
栾青杉, 康元德, 王俊. 2018. 渤海浮游植物群落的长期变化(1959~2015). 渔业科学进展, 39(4): 9-18 |
郭皓, 丁德文, 林凤翱, 等. 2015. 近20a我国近海赤潮特点与发生规律. 海洋科学进展, 33(4): 547-558 DOI:10.3969/j.issn.1671-6647.2015.04.013 |
郭术津, 李彦翘, 张翠霞, 等. 2014. 渤海浮游植物群落结构及与环境因子的相关性分析. 海洋通报, 33(1): 95-105 |
黄文祥, 沈亮夫, 朱琳. 1989. 大连湾赤潮生物——中肋骨条藻(Skeletonema costatum)生态初探. 海洋环境科学, 8(2): 35-39 |
曹春晖, 孙之南, 王学魁, 等. 2006. 渤海天津海域的网采浮游植物群落结构与赤潮植物的初步研究. 天津科技大学学报, 21(3): 34-37 DOI:10.3969/j.issn.1672-6510.2006.03.010 |
康元德. 1991. 渤海浮游植物的数量分布和季节变化. 海洋水产研究, (12): 31-54 |
鹿琳, 2012. 黄渤海浮游植物种多样性及部分种分子鉴定. 青岛: 中国海洋大学硕士学位论文, 1-74
|
屠建波, 张秋丰, 徐玉山, 等. 2011. 渤海湾天津近岸海域首次棕囊藻赤潮初探. 海洋通报, 30(3): 334-337 DOI:10.3969/j.issn.1001-6392.2011.03.016 |
董焕嫦, 黄春秀, 徐国双, 等. 2018. 中国沿海尖刺拟菱形藻的种下分类学研究. 热带海洋学报, 37(1): 12-19 |
蒋红, 崔毅, 陈碧鹃, 等. 2005. 渤海近20年来营养盐变化趋势研究. 海洋水产研究, 26(6): 61-67 |
韩雅琼, 沈永明. 2013. 基于EFDC的渤海冬夏季环流及其影响因素的数值模拟研究. 水动力学研究与进展A辑, 28(6): 733-744 |
喻龙, 郝彦菊. 2009. 烟台四十里湾一次血红哈卡藻赤潮过程的分析. 海洋科学进展, 27(4): 516-522 |
霍文毅, 俞志明, 邹景忠, 等. 2001. 胶州湾中肋骨条藻赤潮与环境因子的关系. 海洋与湖沼, 32(3): 311-318 |
魏皓, 赵亮, 于志刚, 等. 2003. 渤海浮游植物生物量时空变化初析. 青岛海洋大学学报, 33(2): 173-179 |
Blondeau-Patissier D, Gower J F R, Dekker A G et al, 2014. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans. Progress in Oceanography, 123: 123-144 |
Chen Z F, Zhang Q C, Kong F Z et al, 2019. Resolving phytoplankton taxa based on high-throughput sequencing during brown tides in the Bohai Sea, China. Harmful Algae, 84: 127-138 |
da Silva C A, Train S, Rodrigues L C, 2005. Phytoplankton assemblages in a Brazilian subtropical cascading reservoir system. Hydrobiologia, 537(1-3): 99-109 DOI:10.1007/s10750-004-2552-0 |
Ebenezer V, Medlin L K, Ki J S, 2012. Molecular detection, quantification, and diversity evaluation of microalgae. Marine Biotechnology, 14(2): 129-142 |
Gobler C J, Renaghan M J, Buck N J, 2002. Impacts of nutrients and grazing mortality on the abundance of Aureococcus anophagefferens during a New York brown tide bloom. Limnology and Oceanography, 47(1): 129-141 |
Hallegraeff G M, Anderson D M, Cembella A D et al, 2003. Manual on Harmful Marine Microalgae. Paris: Unesco
|
Jeong H J, Kim J S, Park J Y et al, 2005. Stoeckeria algicida n. gen., n. sp. (Dinophyceae) from the coastal waters off Southern Korea: morphology and small subunit ribosomal DNA gene sequence. Journal of Eukaryotic Microbiology, 52(4): 382-390 |
John U, Litaker R W, Montresor M et al, 2014. Formal revision of the Alexandrium tamarense species complex (dinophyceae) taxonomy: the introduction of five species with emphasis on molecular-based (rDNA) classification. Protist, 165(6): 779-804 |
Kong F Z, Yu R C, Zhang Q C et al, 2012. Pigment characterization for the 2011 bloom in Qinhuangdao implicated "brown tide" events in China. Chinese Journal of Oceanology and Limnology, 30(3): 361-370 |
Laroche J, Nuzzi R, Waters R et al, 1997. Brown tide blooms in Long Island's coastal waters linked to interannual variability in groundwater flow. Global Change Biology, 3(5): 397-410 |
Liu D Y, Sun J, Zou J Z et al, 2005. Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Marine Pollution Bulletin, 50(1): 91-94 |
Miralto A, Barone G, Romano G et al, 1999. The insidious effect of diatoms on copepod reproduction. Nature, 402: 173-176 |
Ou L J, Wang D, Huang B Q et al, 2008. Comparative study of phosphorus strategies of three typical harmful algae in Chinese coastal waters. Journal of Plankton Research, 30(9): 1007-1017 |
Pustizzi F, MacIntyre H, Warner M E et al, 2004. Interaction of nitrogen source and light intensity on the growth and photosynthesis of the brown tide alga Aureococcus anophagefferens. Harmful Algae, 3(4): 343-360 |
Qiao L, Yu J, Li Y et al, 2019. Amplicon-based illumina sequencing and quantitative pcr reveals nanoplankton diversity and biomass in surface water of Qinhuangdao Coastal Area, China. Journal of Ocean University of China, 18(4): 962-976 |
Shen Z L, Shi Q, Zheng S et al, 2017. Chemical composition and biomass of Coscinodiscus asteromphalus in Jiaozhou Bay, China. Environmental Monitoring and Assessment, 189(3): 94 |
Song L, Wu J, Du J et al, 2019a. The characteristics and distribution of eukaryotic phytoplankton community in Liaodong Bay, China. Ocean Science Journal, 54(2): 183-203 DOI:10.1007/s12601-019-0007-9 |
Song L, Wu J, Du J et al, 2019b. Comparison of two methods to assess the size structure of phytoplankton community assemblages, in Liaodong Bay, China. Journal of Ocean University of China, 18(5): 1207-1215 DOI:10.1007/s11802-019-3960-0 |
Tada K, Morishita M, Hamada K I et al, 2001. Standing stock and production rate of phytoplankton and a red tide outbreak in a heavily eutrophic embayment, Dokai Bay, Japan. Marine Pollution Bulletin, 42(11): 1177-1186 |
Tang Y Z, Ma Z P, Hu Z X et al, 2019. 3, 000 km and 1, 500-year presence of Aureococcus anophagefferens reveals indigenous origin of brown tides in China. Molecular Ecology, 28(17): 4065-4076 |
Wu Z X, Yu Z M, Song X X et al, 2013. The spatial and temporal characteristics of harmful algal blooms in the southwest Bohai Sea. Continental Shelf Research, 59: 10-17 |
Xu X, Yu Z M, Cheng F J et al, 2017. Molecular diversity and ecological characteristics of the eukaryotic phytoplankton community in the coastal waters of the Bohai Sea, China. Harmful Algae, 61: 13-22 |
Yan T, Zhou M J, Qian P Y, 2002. Combined effects of temperature, irradiance and salinity on growth of diatom Skeletonema costatum. Chinese Journal of Oceanology and Limnology, 20(3): 237-243 |
Yao P, Lei L, Zhao B et al, 2019. Spatial-temporal variation of Aureococcus anophagefferens blooms in relation to environmental factors in the coastal waters of Qinhuangdao, China. Harmful Algae, 86: 106-118 |
Zhang Q C, Qiu L M, Yu R C et al, 2012. Emergence of brown tides caused by Aureococcus anophagefferens Hargraves et Sieburth in China. Harmful Algae, 19: 117-124 |
Zhen Y, Qiao L, Gu B et al, 2016. Characteristics of eukaryotic microalgal community and its abiotic influencing factors during brown tide blooms near Qinhuangdao, China. Harmful Algae, 57: 1-12 |
Zou J Z, Zhou M J, Zhang C, 1993. Ecological features of toxic Nitzschia pungens Grunow in Chinese coastal waters. In: Smayda T J, Shimizu Y eds. Toxic Phytoplankton Blooms in the Sea. Amsterdam: Elsevier Science Publishers, 651-657
|