 2021, Vol. 52
2021, Vol. 52

条斑紫菜Tubulin和Kinesin基因家族鉴定及失水胁迫表达模式分析

http://dx.doi.org/10.11693/hyhz20200600174
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 崔正彩, 孔凡娜, 茅云翔, 孙斌, 王俊皓, 董道英. 2021.
- CUI Zheng-Cai, KONG Fan-Na, MAO Yun-Xiang, SUN Bin, WANG Jun-Hao, DONG Dao-Ying. 2021.
- 条斑紫菜Tubulin和Kinesin基因家族鉴定及失水胁迫表达模式分析
- IDENTIFICATION AND EXPRESSION PATTERN ANALYSIS OF TUBULIN AND KINESIN GENE FAMILY IN PYROPIA YEZOENSIS TO DEHYDRATION STRESSES
- 海洋与湖沼, 52(3): 673-684
- Oceanologia et Limnologia Sinica, 52(3): 673-684.
- http://dx.doi.org/10.11693/hyhz20200600174
文章历史
-
收稿日期:2020-06-21
收修改稿日期:2020-07-19
引用本文
崔正彩, 孔凡娜, 茅云翔, 孙斌, 王俊皓, 董道英. 2021. 条斑紫菜Tubulin和Kinesin基因家族鉴定及失水胁迫表达模式分析[J]. 海洋与湖沼, 52(3): 673-684

CUI Zheng-Cai, KONG Fan-Na, MAO Yun-Xiang, SUN Bin, WANG Jun-Hao, DONG Dao-Ying. 2021. IDENTIFICATION AND EXPRESSION PATTERN ANALYSIS OF TUBULIN AND KINESIN GENE FAMILY IN PYROPIA YEZOENSIS TO DEHYDRATION STRESSES[J]. Oceanologia et Limnologia Sinica, 52(3): 673-684.
条斑紫菜Tubulin和Kinesin基因家族鉴定及失水胁迫表达模式分析
1. 中国海洋大学海洋生命学院 海洋生物遗传学与育种教育部重点实验室 青岛 266003;
2. 海南热带海洋学院水产与生命学院 热带海洋生物资源利用与保护教育部重点实验室 三亚 572022
2. 海南热带海洋学院水产与生命学院 热带海洋生物资源利用与保护教育部重点实验室 三亚 572022
摘要:微管是细胞骨架的主要成分,其结构及动力学机制对提高生物体耐受性具有重要作用。微管网络如何通过结构的动态变化调控适应环境变化已成为胁迫生物学领域的研究热点之一。条斑紫菜(Pyropia yezoensis)能够适应潮间带复杂多变的环境,是研究潮间带大型海藻抗逆机制的良好材料。目前对紫菜微管相关基因家族构成及其生物学功能的研究较少。本研究利用生物信息学方法,在条斑紫菜基因组中共鉴定出4个Tubulin基因(Pyα-tubulin 1、Pyα-tubulin 2、Pyβ-tubulin和Pyγ-tubulin)和11个Kinesin基因(PyKinesin 1—PyKinesin 11),并对其理化性质、基因结构、蛋白特征、染色体定位、系统进化和失水胁迫下的表达模式进行了系统分析。结果显示:条斑紫菜中有3种微管蛋白(Tubulin)亚型;该家族成员散布于1号和2号染色体上,Pyα-tubulin 1和Pyα-tubulin 2为串联重复基因;PyTubulin家族成员在基因结构和蛋白特征方面均较为保守,且在转录水平对失水胁迫不敏感。条斑紫菜中有5种驱动蛋白(Kinesin)亚型,亚家族种类和基因数量均少于高等植物;该家族基因散布于1号、2号和3号染色体上,无串联重复基因;PyKinesin家族成员在基因结构和蛋白特征方面存在一定差异,PyKinesin 1在中高度失水胁迫下表达量显著上调。本研究为进一步理解Tubulin和Kinesin基因家族的进化和功能、解析微管在条斑紫菜响应失水胁迫中的作用奠定了基础。
关键词:条斑紫菜 Tubulin Kinesin 基因家族 失水胁迫
IDENTIFICATION AND EXPRESSION PATTERN ANALYSIS OF TUBULIN AND KINESIN GENE FAMILY IN PYROPIA YEZOENSIS TO DEHYDRATION STRESSES
1. The Key Laboratory of Marine Genetics and Breeding, Ministry of Education, Ocean University of China, Qingdao 266003, China;
2. Key Laboratory of Utilization and Conservation of Tropical Marine Bioresource (Ministry of Education), College of Fisheries and Life Science, Hainan Tropical Ocean University, Sanya 572022, China
2. Key Laboratory of Utilization and Conservation of Tropical Marine Bioresource (Ministry of Education), College of Fisheries and Life Science, Hainan Tropical Ocean University, Sanya 572022, China
Abstract: Microtubules, the main components of the cytoskeleton, play vital important roles in improving the tolerance of organisms by altering their structures in dynamic mechanisms; and they have become one of the research hotspots in the field of stress biology. Pyropia yezoensis is an ideal candidate macroalgae for the investigation of the mechanism of stress resistance because of its fast-changing living conditions in intertidal zone. At present, few studies reported the compositions and biological functions of microtubule-related gene families of P. yezoensis. In this study, we identified four Tubulin genes (Pyα-tubulin 1, Pyα-tubulin 2, Pyβ-tubulin, and Pyγ-tubulin) and 11 Kinesin genes (PyKinesin 1-PyKinesin 11) in P. yezoensis genome using bioinformatics methods. The physicochemical properties, gene structures, protein characteristics, chromosomal locations, phylogenetic evolution, and expression patterns of the two gene families in P. yezoensis under dehydration stress were systematically analyzed. The results show that four Tubulin genes could be divided into three subclusters of Tubulin gene families and they were scattered on chromosome 1 and chromosome 2 of P. yezoensis. Pyα-tubulin 1 and Pyα-tubulin 2 were tandem repeat genes. The PyTubulin family was conserved in gene and protein structures and insensitive to dehydration stress in transcription level. Five subclusters of Kinesin gene families were identified among the 11 Kinesin genes in P. yezoensis, which was less than that of higher plants. PyKinesins were scattered on all the three chromosomes of P. yezoensis and there were no tandem repeat genes. The PyKinesin families had certain differences in gene and protein structures. The expression of PyKinesin 1 was significantly enhanced under the medium and high dehydration stress. This study shall laid foundation for further analysis of microtubules in respond to dehydration stress of P. yezoensis.
Key words:
Pyropia yezoensis Tubulin Kinesin gene family dehydration stress
微管作为细胞骨架的重要组成部分, 是多种非生物胁迫的感受器, 在植物抗逆性中发挥重要作用(Krasylenko et al, 2012), 微管网络结构的排列影响着微管对环境信号的敏感性(Mirabet et al, 2018)。微管响应环境信号变化方式包括介导细胞内物质运输、调节胞内信号的转导、调控细胞壁纤维丝的组装以及调节自身结构动态重排等多种生物学过程(Oakley, 1999; 陈志玲等, 2003; Paradez et al, 2006; Mineyuki, 2007)。α-和β-tubulin是构成微管蛋白亚基的基本结构成分(Nogales et al, 1998), 其构象变化能够调节微管的动态稳定性(Shaw et al, 2003), 使微管骨架通过不断重构响应各种生物和非生物刺激(王朝凤等, 2019), 对微管发挥生物学功能具有重要意义。驱动蛋白Kinesin是沿微管轨道进行长距离运输的动力蛋白超家族(Lee et al, 2004), 具有保守的微管结合位点和运动结构域(Reddy, 2001)。Kinesin家族蛋白不仅能够通过调节微管结合蛋白的运输参与微管结构的调控, 还能够直接结合在微管末端促进微管的解聚(周浩等, 2014), 对微管的动态稳定性及结构重排都具有十分重要的作用。
植物微管通过微管蛋白及其相关蛋白的调节参与多种生物学过程从而适应环境胁迫。拟南芥通过磷脂酶调节微管的稳定性以实现对其耐热性的负调控(Zhang et al, 2017), 通过微管相关蛋白调节皮脂微管的重组以提高其耐盐性(Zhou et al, 2017)。玉米微管通过调节细胞壁结构和胞内膨压以增强对干旱环境的耐受性(Baskin et al, 1999); 杨树叶片通过微管结构的变化调节诱导气孔关闭以减少干旱条件下的水分流失(Swamy et al, 2015); 仙鹤藓(Atrichum undulatum)通过叶片微管重排以迅速对失水胁迫做出响应(陈志玲等, 2003); 石南星绿藻(Asterochloris erici)的Tubulin基因在转录水平显著响应于失水胁迫(Gasulla et al, 2013); 拟南芥(Arabidopsis thaliana) Kinesin-13A基因缺失突变体在干旱条件下比野生型更早出现萎蔫死亡现象(魏丽勤, 2005)。
条斑紫菜生长于潮间带地区, 受潮汐影响, 潮间带地区海水周期性涨落, 条斑紫菜每天都要经历干湿度、温度以及渗透压等各种环境因子的急剧变化, 是研究潮间带大型海藻非生物胁迫响应机制的理想模式生物。目前对潮间带海藻细胞骨架在其适应潮间带环境中的作用机制研究甚少。本研究首次在全基因组水平分析了条斑紫菜微管相关基因Tubulin和Kinesin基因家族的组成及其特点, 并通过实时荧光定量PCR技术探究了叶状体中表达丰度高的3个PyTubulin和2个PyKinesin在失水胁迫处理下的表达模式, 为探究微管在条斑紫菜响应失水胁迫中的作用奠定基础, 也为进一步解析条斑紫菜响应逆境胁迫的调控机制提供支持。
1 材料与方法 1.1 材料 1.1.1 实验材料条斑紫菜(Pyropia yezoensis) RZ纯系, 从海洋生物遗传与育种教育部重点实验室紫菜种质库获取。
1.1.2 实验试剂RNA提取试剂盒(购自OMEGA); DNA酶(购自OMEGA); 反转录试剂盒(购自TAKARA); 琼脂糖凝胶回收试剂盒(购自全式金); 快速质粒小提试剂盒(购自天根); 2×Taq Plus Master Mix (购自南京诺唯赞); ChamQTM SYBR Color qPCR Master Mix (购自南京诺唯赞)。
1.2 方法 1.2.1 基因家族鉴定本研究采用隐马尔可夫模型(Hidden Markov Model, HMM)对条斑紫菜Tubulin和Kinesin基因家族进行鉴定: 从Pfam数据库(http://pfam.xfam.org/)下载Tubulin和Kinesin家族保守结构域HMM模型(PF00022), 通过hmmsearch检索条斑紫菜Tubulin和Kinesin蛋白种子序列, E-value设置为1×e-20; 根据提取的蛋白种子序列构建条斑紫菜特异性HMM模型, 利用所构建模型通过hmmsearch再次检索并提取条斑紫菜Tubulin和Kinesin蛋白序列, 设置E-value < 0.01; 将提取到的蛋白序列去重复后, 根据蛋白ID号提取对应基因序列, 获得PyTubulin和PyKinesin基因家族序列信息。通过软件SnapGene分析蛋白等电点(pI)和分子量(Mw), 通过在线网站Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)预测蛋白亚细胞定位。
1.2.2 基因结构及蛋白序列分析为探究PyTubulin和PyKinesin家族基因结构和蛋白序列特征, 本研究对基因内含子-外显子分布、蛋白保守基序和蛋白保守结构域进行预测分析。根据PyTubulin和PyKinesin家族基因组信息和转录本ID进行基因结构注释, 从Gff注释文件中获得基因位置信息, 通过网站GSDS (http://gsds.cbi.pku.edu.cn/)分析内含子-外显子分布。通过在线网站MEME (http://meme-suite.org/index.html)预测蛋白保守基序, 分布模型选择anr, 预测数量设置为10, 最短motif设置为6 aa, 最长motif设置为50 aa。将蛋白序列提交至在线网站SMART(http://smart.embl.de/)预测蛋白结构域, 预测模型为Normal。
1.2.3 染色体定位及基因复制分析本研究通过SAMtools (http://samtools.sourceforge.net/)分析获得条斑紫菜的染色体长度信息, 根据PyTubulin和PyKinesin基因ID在Gff文件中提取相应的基因位置信息, 将提取到的染色体长度信息和基因位置信息提交至网站MapGene2 Chrom (http://mg2c.iask.in/mg2c_v2.1/), 绘制基因在染色体上的分布图。为探究2个家族中基因复制情况, 本研究以两条序列一致度 > 75%, 比对区域序列长度 > 较长序列的75%, 基因距离 < 100 kb为标准进行串联重复基因筛选, 通过ClustalW比对串联重复基因的CDS序列, 根据输出文件计算得到串联重复基因的非同义替换率/同义替换率(Ka/Ks)值, 并分析串联重复基因的进化模式。
1.2.4 基因家族系统进化分析系统进化分析所用脐形紫菜(Porphyra umbilicalis)、紫球藻(Porphyridium purpureum)、角叉菜(Chondrus crispus)、莱茵衣藻(Chlamydomonas reinhardtii)、团藻(Volvox carteri)、长囊水云(Ectocarpus siliculosus)、拟南芥(Arabidopsis thaliana)以及水稻(Oryza sativa) 蛋白序列均下载自NCBI数据库(https://www.ncbi. nlm.nih.gov/), 条斑紫菜(P. yezoensis)和坛紫菜(P. haitanensis)相关序列均来自本实验室通过二代测序和三代测序获得的高质量基因组。利用本研究中基因家族鉴定方法, 从NCBI下载的蛋白序列信息中提取各物种Tubulin和Kinesin家族成员蛋白序列, 利用ClustalW对提取的蛋白序列进行多序列比对, 使用Mega7构建系统进化树, 建树方法为最大似然法(Maximum Likelihood), 建树模型为泊松模型(Poisson Model), 迭代1000次。
1.2.5 失水胁迫材料处理参考Kim等人(2009)对紫菜叶状体失水的处理方法, 挑选同批培养的生长状态良好且日龄为45 d的条斑紫菜叶状体进行不同程度失水胁迫处理, 处理条件分别设置为失水10%、30%、50%、60%、70%、80%, 失水50%后复水30 min, 失水80%后复水30 min和失水80%后复水60 min, 每组设置三个生物学平行。将藻体置于(10±1) ℃培养箱中恒温失水, 光照为35—45 μmol photons/(m2·s), 样品失水过程需将藻体平整展开以保证失水均匀, 通过二氧化硅促进失水以缩短中高度失水处理组的失水时间, 将失水时间控制在0.5 h以内。藻体失水率按公式(1)计算, 其中藻体鲜重为处理前去除藻体表面残留的水分后测得的重量, 藻体干重为60 ℃下将新鲜藻体烘干至藻体重量不再变化时测得的重量。
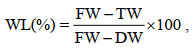 (1)
(1)式中, WL: 藻体失水率; FW: 藻体鲜重; TW: 失水后藻体重量; DW: 藻体干重。
1.2.6 基因表达水平检测通过对实验室前期转录组数据分析, 选取在条斑紫菜叶状体中表达丰度较高的Pyα-tubulin 2、Pyβ-tubulin、Pyγ-tubulin、PyKinesin 1和PyKinesin 10, 探究PyTubulin和PyKinesin在不同失水胁迫下的表达模式。以未经失水处理的条斑紫菜RZ纯系叶状体为对照组, 不同失水和复水条件处理的条斑紫菜RZ叶状体为实验组, 每组设置3个生物学平行和3个技术重复, 通过qRT-PCR技术检测目的基因在不同程度失水胁迫下的表达变化, 引物序列如表 1所示。内参基因为不同失水条件下在条斑紫菜中表达稳定的PyUBC基因(Gao et al, 2018), 在内参基因的矫正下, 用2-ΔΔCt法计算得到目的基因的相对表达量, 并通过数据统计软件SPSS (Statistical Product and Service Solutions)对不同失水胁迫条件下基因的相对表达量进行独立样本T检验, 置信区间百分比为95%。
表 1 qRT-PCR所用引物序列 Tab. 1 Primer sequences used in qRT-PCR
| 基因名称 | 引物序列(5′—3′) |
| PyUBC | F: CGCTGACCGTTTCCAAG |
| R: CGACTGCGGTTGGACTT | |
| Pyα-tubulin 2 | F: GGTTTCACCGTCTACCCTTCG |
| R: CGCCTCATTGTCCAGCATCA | |
| Pyβ-tubulin | F: TGATTGGTTACGCCCCGCT |
| R: CCGCTGTTCTTCGTCTGGA | |
| Pyγ-tubulin | F: ATTGGGTGATGGGGGAGTCT |
| R: GCTTGGCACTTGGTACAGGG | |
| PyKinesin 1 | F: CGGCTCGACCTGTCCAAGTA |
| R: CGCAAAGACGGCTTCCAC | |
| PyKinesin 10 | F: CGCAGGCCATCAATAAGTCG |
| R: CGCCCAATCTCACACGCAT |
经过hmmsearch检索和比对筛选, 最终确定条斑紫菜基因组中Tubulin基因家族有4个成员, 均含有Tubulin家族保守结构域, 分别命名为Pyα-tubulin 1、Pyα-tubulin 2、Pyβ-tubulin和Pyγ-tubulin。cDNA序列最短为1377 bp, 最长为2529 bp, 编码多肽的氨基酸数量为454—465, 分子量从49.51— 51.42 kDa, 等电点从4.33到4.97不等。对蛋白进行亚细胞定位预测, 预测结果显示Pyα-tubulin 1和Pyα-tubulin 2定位于线粒体, Pyβ-tubulin和Pyγ-tubulin定位于细胞质。条斑紫菜Kinesin家族共鉴定出11个成员, 均含有驱动蛋白保守运动域, 将11个PyKinesin分别命名为PyKinesin 1—PyKinesin 11, cDNA序列最短为1272 bp, 最长为4101 bp, 编码多肽的氨基酸数量为423—1366, 分子量从41.91—128.26 kDa, 等电点从4.79到10.96不等。亚细胞定位预测结果显示有8个蛋白定位于细胞核, 3个蛋白定位于细胞质。PyTubulin和PyKinesin基因家族蛋白特征如表 2所示。
表 2 条斑紫菜Tubulin和Kinesin基因家族蛋白特征 Tab. 2 Protein characteristics of Tubulin and Kinesin gene family in P. yezoensis
| 基因名称 | 基因ID | 编码序列 | 氨基酸 | 分子量(kDa) | 等电点 | 亚细胞定位 |
| Pyα-tubulin 1 | py10637.t1 | 1977 | 455 | 49.51 | 4.97 | 线粒体 |
| Pyα-tubulin 2 | py10654.t1 | 1997 | 454 | 49.53 | 4.9 | 线粒体 |
| Pyβ-tubulin | py04248.t1 | 1377 | 458 | 50.18 | 4.33 | 细胞质 |
| Pyγ-tubulin | py03680.t1 | 2529 | 465 | 51.42 | 4.92 | 细胞质 |
| PyKinesin 1 | py03095.t1 | 4101 | 1366 | 128.26 | 10.74 | 细胞核 |
| PyKinesin 2 | py03345.t1 | 2703 | 900 | 85.95 | 6.55 | 细胞核 |
| PyKinesin 3 | py04096.t1 | 3048 | 1015 | 95.05 | 10.79 | 细胞核 |
| PyKinesin 4 | py04806.t1 | 1272 | 423 | 41.91 | 10.15 | 细胞质 |
| PyKinesin 5 | py06213.t1 | 2985 | 994 | 90.96 | 4.85 | 细胞核 |
| PyKinesin 6 | py06963.t1 | 3783 | 1260 | 120.10 | 5.07 | 细胞质 |
| PyKinesin 7 | py07128.t1 | 2346 | 781 | 73.72 | 10.96 | 细胞核 |
| PyKinesin 8 | py07129.t1 | 3969 | 1323 | 123.99 | 4.79 | 细胞核 |
| PyKinesin 9 | py08263.t1 | 1818 | 605 | 54.64 | 6.62 | 细胞质 |
| PyKinesin 10 | py10378.t1 | 3400 | 712 | 75.26 | 6.34 | 细胞核 |
| PyKinesin 11 | py10448.t1 | 3879 | 1292 | 120.30 | 5.94 | 细胞核 |
条斑紫菜Tubulin家族基因结构和蛋白结构域分析结果表明, PyTubulin基因结构简单, 4个基因均不含内含子(图 1a); 蛋白保守基序相似性较高, 所有家族成员均包含motif1-motif 8, Pyγ-tubulin缺少motif 9和motif 10, Pyβ-tubulin缺少motif 10 (图 1b); 蛋白结构域保守, 4个Py-tubulin蛋白氨基端均包含一个保守的Tubulin蛋白GTP酶结构域, 羧基端均含有一个保守的Tubulin羧基端结构域(图 1c)。
 |
| 图 1 PyTubulin基因家族基因结构和蛋白结构域 Fig. 1 Gene structure and protein domain of PyTubulin gene family 注: a: PyTubulin家族内含子-外显子分布; b: PyTubulin家族motif(基序)分布; c: PyTubulin家族蛋白结构域分布; d: PyTubulin家族motif(基序)序列信息 |
条斑紫菜Kinesin家族基因结构和蛋白结构域分析结果表明, PyKinesin基因结构较为简单, 其中PyKinesin 2、PyKinesin 3、PyKinesin 5和PyKinesin 9均含有1个内含子, 其余7个基因均不含内含子(图 2a); 不同蛋白保守基序的数量和分布差异较大, 同一蛋白中某一特定motif重复出现频率高, 如motif 5在PyKinesin 3中出现了10次, 在PyKinesin 10中没有出现, motif 9在PyKinesin 8中连续出现8次(图 2b); 蛋白结构域保守, 11个蛋白均含有典型的驱动蛋白运动域, 其中有6个Py-Kinesin的运动域在氨基端, 3个Py-Kinesin的运动域在羧基端, 另外2个Py-Kinesin的运动域位于蛋白序列的中间位置(图 2c)。
 |
| 图 2 PyKinesin基因家族基因结构和蛋白结构域 Fig. 2 Gene structure and protein domain of PyKinesin gene family 注: a: PyKinesin家族内含子-外显子分布; b: PyKinesin家族motif(基序)分布; c: PyKinesin家族蛋白结构域分布; d: PyKinesin家族motif(基序)序列信息 |
PyTubulin基因家族染色体定位分析结果表明Pyα-tubulin 1、Pyα-tubulin 2和Pyβ-tubulin均定位于2号染色体, Pyγ-tubulin定位于1号染色体(图 3)。基因复制分析结果表明Pyα-tubulin 1和Pyα-tubulin 2为串联重复基因, 其非同义替换率Ka=0.0166, 同义替换率Ks=0.8126, Ka/Ks= 0.0204。根据Ka/Ks值估算串联重复基因的进化选择模式, Ka/Ks=1表示中性选择, Ka/Ks < 1表示纯化选择, Ka/Ks > 1表示正选择(李晓翠等, 2020), 故Pyα-tubulin 1和Pyα-tubulin 2的进化选择模式为纯化选择。
 |
| 图 3 PyTubulin基因家族染色体定位 Fig. 3 Chromosomal location of PyTubulin gene family 注: PY-1: 条斑紫菜1号染色体; PY-2: 条斑紫菜2号染色体 |
PyKinesin家族染色体定位结果显示, PyKinesin 2、PyKinesin 5、PyKinesin 6和PyKinesin 11分布于1号染色体, PyKinesin 3、PyKinesin 7、PyKinesin 8、PyKinesin 9和PyKinesin 10分布于2号染色体, PyKinesin 1位于3号染色体(图 4)。PyKinesin 4由于基因组Gff文件中缺少注释信息, 无法定位。基因复制分析结果表明PyKinesin基因家族没有串联重复基因。
 |
| 图 4 PyKinesin基因家族染色体定位 Fig. 4 Chromosomal location of PyKinesin gene family 注: PY-1: 条斑紫菜1号染色体; PY-2: 条斑紫菜2号染色体; PY-3: 条斑紫菜3号染色体 |
对来自条斑紫菜(Py)、坛紫菜(Ph)、脐形紫菜(Pu)、紫球藻(Pp)、角叉菜(Cc)、莱茵衣藻(Cr)、团藻(Vc)、长囊水云(Es)、拟南芥(At)以及水稻(Os)的72条Tubulin蛋白序列构建系统进化树(图 5)。结果表明, 10个物种的Tubulin蛋白均包含α-tubulin、β-tubulin和γ-tubulin 3大亚家族, 在莱茵衣藻和团藻两种绿藻中还存在δ-tubulin、ε-tubulin和η-tubulin 3种微管蛋白亚型。在α-tubulin、β-tubulin和γ-tubulin 3大亚家族分支中, 红藻、褐藻和绿色植物的Tubulin分别形成了独立的姐妹枝, 条斑紫菜的Tubulin基因均与其他4种红藻的Tubulin聚在一起, 莱茵衣藻和团藻两种绿藻与拟南芥和水稻的Tubulin聚在一簇, 长囊水云单独成一簇。
 |
| 图 5 PyTubulin基因家族系统进化分析 Fig. 5 Phylogenetic analysis of PyTubulin gene family |
对条斑紫菜(Py)、坛紫菜(Ph)、脐形紫菜(Pu)、角叉菜(Cc)、莱茵衣藻(Cr)、团藻(Vc)、长囊水云(Es)以及拟南芥(At) Kinesin家族的系统进化分析显示(图 6), 152个Kinesin家族蛋白共分为11个Group, 其中Group Ⅰ和Group Ⅺ为藻类中特有的亚型。红藻分布于其中6个Group, 拟南芥和绿藻均分布于其中9个Group, 褐藻长囊水云在11个Group中均有分布。PyKinesin家族的11个蛋白分布在其中5个Group中, PyKinesin4单独聚在Group Ⅱ的一个分支中, 其余10个PyKinesin均和其他红藻Kinesin聚在一起。系统进化分析结果表明, 4个红藻的Kinesin基因亚家族种类和基因数量均明显少于高等植物和其他藻类, 这可能是由于Kinesin家族在进化过程中发生了基因家族扩张。
 |
| 图 6 PyKinesin基因家族系统进化分析 Fig. 6 Phylogenetic analysis of PyKinesin gene family |
选取条斑紫菜叶状体中表达丰度高的Pyα-tubulin 2、Pyβ-tubulin、Pyγ-tubulin、PyKinesin 1和PyKinesin 10作为目的基因, 利用qRT-PCR技术检测分析5个基因在不同失水和复水条件下的转录变化, 检测结果如图 7—10所示。Pyα-tubulin 2、Pyβ-tubulin和Pyγ-tubulin基因在不同失水和复水条件下表现出不同的表达模式, 但表达量变化均不显著, 对失水胁迫没有明显的响应。2个PyKinesin基因对失水胁迫呈现出不同的响应模式, PyKinesin 1对失水胁迫敏感, 其表达量随着失水程度的增加显著上调, 在失水50%、60%、70%以及80%时达到对照组的2倍以上, 复水后表达量下调, 在失水80%复水60 min后下调显著, PyKinesin 10在不同失水和复水胁迫下表达量均无显著变化。
 |
| 图 7 不同失水条件下PyTubulin基因表达量 Fig. 7 Expression of PyTubulin under different dehydration conditions 注: 10%: 失水10%处理组; 30%: 失水30%处理组; 50%: 失水50%处理组; 60%: 失水60%处理组; 70%: 失水70%处理组; 80%: 失水80%处理组; 相对表达量=表达量平均值±标准差(n=3) |
 |
| 图 8 不同复水条件下PyTubulin基因表达量 Fig. 8 Expression of PyTubulin under different rehydration conditions 注: a: 失水50%复水组; b: 失水80%复水组; 50%: 失水50%处理组; 80%: 失水80%处理组; 50%-30 min: 失水50%后复水30 min处理组; 80%-30 min: 失水80%后复水30 min处理组; 80%-60 min: 失水80%后复水60 min处理组; 相对表达量=表达量平均值±标准差(n=3) |
 |
| 图 9 不同失水条件下PyKinesin基因表达量 Fig. 9 Expression of PyKinesin under different dehydration conditions 注: 10%: 失水10%处理组; 30%: 失水30%处理组; 50%: 失水50%处理组; 60%: 失水60%处理组; 70%: 失水70%处理组; 80%: 失水80%处理组; 相对表达量=表达量平均值±标准差(n=3); **: P < 0.01 |
 |
| 图 10 不同复水条件下PyKinesin基因表达量 Fig. 10 Expression of PyKinesin under different rehydration conditions 注: a: 失水50%复水组; b: 失水80%复水组; 50%: 失水50%处理组; 50%-30 min: 失水50%后复水30 min处理组; 80%: 失水80%处理组; 80%-30 min: 失水80%后复水30 min处理组; 80%-60 min: 失水80%后复水60 min处理组; 相对表达量=表达量平均值±标准差(n=3); **: P < 0.01 |
微管是细胞骨架的重要组成部分, 目前在真核生物中共鉴定出7种微管蛋白, 分别是α、γ、β、δ、ε、ζ和η微管蛋白, α、β、γ微管蛋白基因存在于所有真核生物中, 是微管聚合的必需基因, δ、ε、ζ和η微管蛋白基因只在少数真核生物中存在(Dutcher, 2001)。在高等植物中通常有多个α/β-tubulin基因, 棉花中至少有13个α-tubulin, 水稻中至少有8个β-tubulin (Radchuk, 2008), 而白杨中至少存在20个β-tubulin (Oakley et al, 2007)。本研究发现, 所分析的5种红藻中只有α、β和γ 3种微管蛋白基因, 其中条斑紫菜中含有2个α-tubulin、1个β-tubulin和1个γ-tubulin, 基因数量明显少于高等植物。莱茵衣藻和团藻两种绿藻中除了α、β和γ三种微管蛋白外, 还存在δ-tubulin、ε-tubulin和η-tubulin。高等植物拟南芥基因组拥有61种驱动蛋白的编码基因, 是植物基因组中最大的驱动蛋白库, 硅藻中有25种驱动蛋白, 与绿藻驱动蛋白的数量相近, 而红藻驱动蛋白(Kinesin)种类明显少于高等植物和其他藻类, Cyanidioschyzon merolae中只有5种驱动蛋白(Richardson et al, 2006), 脐形紫菜只鉴定出4种驱动蛋白(Brawley et al, 2017), 本研究在条斑紫菜中鉴定出了11个驱动蛋白基因, 分为5个亚家族。条斑紫菜隶属于红藻门, 后者作为一种古老的物种, 其微管及其相关蛋白家族成员数量之少, 可能与其在进化过程中缺乏薄壁组织且无法形成复杂的多细胞结构有关(Brawley et al, 2017)。
高等植物Tubulin基因中通常含有多个内含子, 拟南芥α1-tubulin中包含4个内含子(Ludwig et al, 1988), 毛白杨α-tubulin基因中也均包含3—4个内含子(饶国栋等, 2015), 而条斑紫菜Tubulin家族基因结构比高等植物简单, 4个Pytubulin均不含内含子。衣藻和被子植物所有的驱动蛋白(Kinesin)都含有内含子, 而红藻驱动蛋白普遍缺乏内含子, 条斑紫菜驱动蛋白大多不含内含子, 少数驱动蛋白基因只有1个内含子, 这与其他红藻驱动蛋白内含子的数量相近, 也支持了红藻基因在进化过程中发生了内含子丢失这一观点(Hoef et al, 2005)。
微管骨架是植物细胞响应胁迫信号的传感器, 胁迫会引起微管骨架的解体和重新聚合(Ma et al, 2019), 但不同物种中微管结构对失水胁迫的响应方式不同。玉米微管在失水胁迫下参与调控气孔形态和细胞壁结构, 并通过控制纤维素微纤丝的形成来调节细胞壁的形成和膨压以增强对干旱的耐受性(Baskin et al, 1999), 而细胞膨压与微管的排列方向有关, 螺旋藻微管随着膨压的升高从纵向/斜向排列向横向排列转换(Iwata et al, 2001)。仙鹤藓(Atrichum undulatum)微管骨架在失水胁迫影响下从有规律排列的细丝状重排为无规律排列的较粗的微管束(陈志玲等, 2003)。油菜幼苗细胞的快速失水会导致微管的解聚, 但在低失水胁迫压力下, 微管结构并没有明显的变化(Bagniewska-Zadworna, 2008)。研究也发现, 失水胁迫下Tubulin基因在转录水平上的表达量也呈现显著变化, 如石南星绿藻(Asterochloris erici) 2个β-tubulin基因的转录产物在失水胁迫后期显著积累(Gasulla et al, 2013), 表明Tubulin基因转录产物的积累与微管骨架形态的变化及结构的重排有密切关联。驱动蛋白(Kinesin)利用ATP水解释放的能量沿微管运动, 具有保守的运动蛋白结构域, 能够牵动分子货物沿着微管作长距离运输(陈珊珊, 2019), 直接参与到细胞骨架在多种生物过程中的排列调节。
项目组前期转录组研究表明, 条斑紫菜的Tubulin和Kinesin家族在叶状体和丝状体中的表达具有显著的世代特异性。本研究进一步采用qRT-PCR方法检测了在叶状体中高表达的Pyα-tubulin 2、Pyβ-tubulin、Pyγ-tubulin、PyKinesin 1和PyKinesin 10在失水胁迫下的表达模式, 结果显示, 不同成员在失水、复水胁迫下的表达模式不同。Pyα-tubulin 2、Pyβ-tubulin和Pyγ-tubulin三个基因在不同失水胁迫条件下表达量虽出现浮动, 但并无显著变化, 在转录水平对失水胁迫响应不显著, 与之前转录组结果一致。转录组分析发现, Pyα-tubulin 1在不同失水和复水条件下表达丰度变化不显著, 因此推测该基因在转录水平对失水胁迫也不敏感。PyKinesin 1在中高度失水条件下表达量显著上调, 在复水后表达水平又迅速降低, PyKinesin 10在失水胁迫下表达量没有显著变化。转录组分析表明, PyKinesin 1在高度失水时表达丰度显著增加, 复水后表达丰度降低, PyKinesin 10在不同失水和复水条件下表达丰度变化不显著, 与本实验结果一致。转录组分析发现, PyKinesin家族其余基因在失水和复水过程中表达丰度均较低, 且没有发生显著变化, 推测该家族其余基因在转录水平对失水胁迫不敏感。对PyKinesin基因家族的系统进化分析发现PyKinesin 1属于Kinesin 13家族, 有研究表明, Kinesin 13家族成员具有解聚酶催化活性, 能够结合在微管末端促进微管的解聚, 在细胞发育、细胞分离以及细胞形态调节等过程均发挥了重要作用(周浩等, 2014)。据此推测, PyKinesin 1在条斑紫菜细胞响应失水胁迫时, 可能在微管结构的动态调节中发挥重要作用, 但该基因在微管结构的动态调节中如何发挥作用还需要进一步研究。
4 结语本研究利用生物信息学方法鉴定了条斑紫菜Tubulin和Kinesin基因家族, 解析了PyTubulin和PyKinesin家族成员的组成、基因结构、蛋白结构和进化特点, 探究了在叶状体中高表达的PyTubulin和PyKinesin基因家族成员在不同失水胁迫条件下的表达变化, 为进一步了解微管相关基因在条斑紫菜响应失水胁迫中的作用奠定了基础。
参考文献
王朝凤, 孔照胜. 2019. 植物细胞微管研究进展. 中国细胞生物学学报, 41(3): 372-380 |
李晓翠, 康凯程, 黄先忠, 等. 2020. 小拟南芥MKK基因家族全基因组鉴定及进化和表达分析. 遗传, 42(4): 403-421 |
陈志玲, 欧阳浩淼, 刘祥林, 等. 2003. 微管骨架在苔藓植物适应干旱胁迫应答中的功能研究. 生物工程学报, 19(3): 317-321 DOI:10.3321/j.issn:1000-3061.2003.03.012 |
陈珊珊, 2019. 番茄表皮毛形态和内部细胞骨架排布的研究. 杭州: 浙江大学硕士学位论文
|
周浩, 沈涛. 2014. 微管解聚酶驱动蛋白家族成员KIF2A研究进展. 生命科学, 26(8): 866-873 |
饶国栋, 睢金凯, 张建国. 2015. 毛白杨α-微管蛋白基因家族的克隆与序列分析. 林业科学研究, 28(1): 44-49 |
魏丽勤, 2005. 动蛋白Kinesin-13A和类锚蛋白AtANK1在植物细胞中的分布及功能. 北京: 中国农业大学博士学位论文
|
Bagniewska-Zadworna A, 2008. The root microtubule cytoskeleton and cell cycle analysis through desiccation of Brassica napus seedlings. Protoplasma, 233(3-4): 177-185 DOI:10.1007/s00709-008-0001-z |
Baskin T I, Meekes H T H N, Liang B M et al, 1999. Regulation of growth anisotropy in well-watered and water-stressed maize roots. Ⅱ. Role of cortical microtubules and cellulose microfibrils. Plant Physiology, 119(2): 681-692 DOI:10.1104/pp.119.2.681 |
Brawley S H, Blouin N A, Ficko-Blean E et al, 2017. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proceedings of the National Academy of Sciences of the United States of America, 114(31): E6361-E6370 DOI:10.1073/pnas.1703088114 |
Dutcher S K, 2001. The tubulin fraternity: alpha to eta. Current Opinion in Cell Biology, 13(1): 49-54 DOI:10.1016/S0955-0674(00)00173-3 |
Gao D, Kong F N, Sun P P et al, 2018. Transcriptome-wide identification of optimal reference genes for expression analysis of Pyropia yezoensis responses to abiotic stress. BMC Genomics, 19(1): 251 DOI:10.1186/s12864-018-4643-8 |
Gasulla F, Jain R, Barreno E et al, 2013. The response of Asterochloris erici (Ahmadjian) Skaloud et Peksa to desiccation: a proteomic approach. Plant, Cell & Environment, 36(7): 1363-1378 |
Hoef E K, Shrestha R P, Lapidot M et al, 2005. Actin phylogeny and intron distribution in bangiophyte red algae (rhodoplantae). Journal of Molecular Evolution, 61(3): 360-371 DOI:10.1007/s00239-004-0366-3 |
Iwata K, Tazawa M, Itoh T, 2001. Turgor pressure regulation and the orientation of cortical microtubules in Spirogyra cells. Plant and Cell Physiology, 42(6): 594-598 DOI:10.1093/pcp/pce073 |
Kim J K, Kraemer G P, Yarish C, 2009. Research note: comparison of growth and nitrate uptake by New England Porphyra species from different tidal elevations in relation to desiccation. Phycological Research, 57(2): 152-157 DOI:10.1111/j.1440-1835.2009.00533.x |
Krasylenko Y A, Yemets A I, Blume Y B, 2012. Cytoskeleton-mediated signalling pathways in UV-B perception by plant cell. Emirates Journal of Food and Agriculture, 24(6): 557-564 DOI:10.9755/ejfa.v24i6.557564 |
Lee Y R J, Liu B, 2004. Cytoskeletal motors in Arabidopsis. Sixty-one kinesins and seventeen myosins. Plant Physiology, 136(4): 3877-3883 DOI:10.1104/pp.104.052621 |
Ludwig S R, Oppenheimer D G, Silflow C D et al, 1988. The α1-tubulin gene of Arabidopsis thaliana: primary structure and preferential expression in flowers. Plant Molecular Biology, 10(4): 311-321 DOI:10.1007/BF00029881 |
Ma H X, Liu M, 2019. The microtubule cytoskeleton acts as a sensor for stress response signaling in plants. Molecular Biology Reports, 46(5): 5603-5608 DOI:10.1007/s11033-019-04872-x |
Mineyuki Y, 2007. Plant microtubule studies: past and present. Journal of Plant Research, 120(1): 45-51 DOI:10.1007/s10265-006-0063-y |
Mirabet V, Krupinski P, Hamant O et al, 2018. The self-organization of plant microtubules inside the cell volume yields their cortical localization, stable alignment, and sensitivity to external cues. PLoS Computational Biology, 14(2): e1006011 DOI:10.1371/journal.pcbi.1006011 |
Nogales E, Wolf S G, Downing K H, 1998. Structure of the αβ tubulin dimer by electron crystallography. Nature, 391(6663): 199-203 DOI:10.1038/34465 |
Oakley B R, 1999. γ-Tubulin. Current Topics in Developmental Biology, 49: 27-54 |
Oakley R V, Wang Y S, Ramakrishna W et al, 2007. Differential expansion and expression of α-and β-tubulin gene families in Populus. Plant Physiology, 145(3): 961-973 DOI:10.1104/pp.107.107086 |
Paradez A, Wright A, Ehrhardt D W, 2006. Microtubule cortical array organization and plant cell morphogenesis. Current Opinion in Plant Biology, 9(6): 571-578 DOI:10.1016/j.pbi.2006.09.005 |
Radchuk V V, 2008. The transcriptome of the tubulin gene family in plants. In: Blume, Y B, Baird, W V eds. The Plant Cytoskeleton: A Key Tool For Agro-biotechnology. New York, NY: Springer Dordrecht, 219-241
|
Reddy A S N, 2001. Molecular motors and their functions in plants. International Review of Cytology, 204: 97-178 |
Richardson D N, Simmons M P, Reddy A S, 2006. Comprehensive comparative analysis of kinesins in photosynthetic eukaryotes. BMC Genomics, 7: 18 DOI:10.1186/1471-2164-7-18 |
Shaw S L, Kamyar R, Ehrhardt D W, 2003. Sustained microtubule treadmilling in Arabidopsis cortical arrays. Science, 300(5626): 1715-1718 DOI:10.1126/science.1083529 |
Swamy P S, Hu H, Pattathil S et al, 2015. Tubulin perturbation leads to unexpected cell wall modifications and affects stomatal behaviour in Populus. Journal of Experimental Botany, 66(20): 6507-6518 DOI:10.1093/jxb/erv383 |
Zhang Q, Song P, Qu Y N et al, 2017. Phospholipase Dδ negatively regulates plant thermotolerance by destabilizing cortical microtubules in Arabidopsis. Plant, Cell & Environment, 40(10): 2220-2235 |
Zhou S, Chen Q H, Li X Y et al, 2017. MAP65-1 is required for the depolymerization and reorganization of cortical microtubules in the response to salt stress in Arabidopsis. Plant Science, 264: 112-121 DOI:10.1016/j.plantsci.2017.09.004 |