 2021, Vol. 52
2021, Vol. 52

基于大涡模拟与被动示踪物模型的人工鱼礁数值研究

http://dx.doi.org/10.11693/hyhz20210300075
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 王者也, 李爽. 2021.
- WANG Zhe-Ye, LI Shuang. 2021.
- 基于大涡模拟与被动示踪物模型的人工鱼礁数值研究
- NUMERICAL SIMULATION OF CUBIC ARTIFICIAL REEF UNDER LARGE EDDY IN PASSIVE SCALAR MODEL
- 海洋与湖沼, 52(6): 1376-1387
- Oceanologia et Limnologia Sinica, 52(6): 1376-1387.
- http://dx.doi.org/10.11693/hyhz20210300075
文章历史
-
收稿日期:2021-03-30
收修改稿日期:2021-07-28
引用本文
王者也, 李爽. 2021. 基于大涡模拟与被动示踪物模型的人工鱼礁数值研究[J]. 海洋与湖沼, 52(6): 1376-1387

WANG Zhe-Ye, LI Shuang. 2021. NUMERICAL SIMULATION OF CUBIC ARTIFICIAL REEF UNDER LARGE EDDY IN PASSIVE SCALAR MODEL[J]. Oceanologia et Limnologia Sinica, 52(6): 1376-1387.
基于大涡模拟与被动示踪物模型的人工鱼礁数值研究
浙江大学海洋学院 舟山 316021
摘要:投放人工鱼礁是解决海洋生态环境问题的重要措施之一。通过使用并行大涡模拟模式(the parallelizeda large-eddy simulation model,PALM)及被动示踪物模型模块,研究了不同流速条件(0.1,0.2,0.4和0.6 m/s)下方型人工鱼礁对流场形态、营养盐的抬升作用、和湍流动能收支的影响。研究表明,鱼礁的存在使得其附近垂向速度增大,产生上升流。受到上升流的抬升作用,鱼礁底部的示踪物迅速进入海洋上层,之后遇到鱼礁后方的背涡流,示踪物的抬升受阻,高度逐渐降低。在上升流区域以及背涡流区域的共同影响下,示踪物抬升区域的最大高度与来流流速无关。不同上升流定义对应的上升流区域的高度与来流流速均不相关;上升流区域的最大速度、平均速度与来流流速都成线性增加的关系;然而随着来流流速的增大,不同上升流定义对应的上升流区域面积的变化趋势却完全不同。投放鱼礁后,鱼礁区域底部的混合增强,区域底部的能量被输运至上层。这说明,鱼礁的存在不仅对营养物质具有抬升作用,还能将能量向上输运。
关键词:大涡模拟 人工鱼礁 被动示踪物 湍流动能收支
NUMERICAL SIMULATION OF CUBIC ARTIFICIAL REEF UNDER LARGE EDDY IN PASSIVE SCALAR MODEL
Ocean College, Zhejiang University, Zhoushan 316021, China
Abstract: Artificial reef is a widely used tool for marine ecosystem improvement. A numerical simulation was conducted in the parallel large eddy simulation model (PALM) in passive scalar mode to explore the effects of solid cubic artificial reef on flow field, turbulent kinetic energy (TKE) budget, and nutrient uplift under four inflow velocities (0.1, 0.2, 0.4, and 0.6 m/s). Results show that artificial reef significantly increases the vertical velocities near reef, and underpin inflow into upwelling. Scalars at the reef bottom are pushed up to the ocean surface quickly and then sunk by vortex at the reef rear. Under the joint effects of upwelling and vortex, the maximum height of uplift of passive scalar show no relation with inflow velocities. In addition, different definitions of upwelling present no influences on the heights and velocities of upwelling. Both average and maximum velocities of upwelling are increased linearly with inflow velocities. However, the area of upwelling change demonstrates a totally different trend. After reef is placed, mixing at the bottom is strengthened, dissipation and shear are remarkably enlarged, and energy at the bottom is transported to the upper layers. Therefore, artificial reef can not only uplift nutrients, but also transport energy upward.
Key words:
large eddy simulation artificial reef passive scalar TKE (turbulent kinetic energy)
随着海洋生态环境逐步恶化, 许多养殖场不再适合养殖鱼类、软体动物和甲壳类动物。事实上, 因为大多数渔业资源已经被过度开发, 当前渔业系统的可持续性正受到质疑(Garcia et al, 2001)。联合国粮食及农业组织调查(FAO, 2020)显示, 尽管水产养殖业依旧在发展, 海洋捕鱼业似乎已经到达上限。就资源开发的角度而言, 全球生态系统压力过大, 生物多样性减少, 并面临崩溃的危险。因为资源的逐渐枯竭, 人造建筑物被用来增加鱼类丰度以及多样性, 改善栖息环境以及修复受损的鱼礁(Lan et al, 2004)。目前, 存在着很多关于人工鱼礁的定义。Seaman (2000)将人工鱼礁定义为放置在海底的自然或人造物体, 其作用是调控海洋环境中的生物和物理参数。Grace (2001)研究了影响人工鱼礁寿命的环境因素。大量具有不同材料成分和结构的人工鱼礁在世界各地被广泛利用(Walker et al, 2002)。海洋是由众多物种组成的生态系统, 从简单的浮游植物到更高营养层的鱼类, 形成一个相互影响和相互依赖的海洋食物网。人工鱼礁的投放可以改变流场、颗粒物分布, 有利于营养盐的积累(Davis et al, 1982; Ambrose et al, 1990)。因此, 鱼礁可以形成适宜鱼类生存、繁殖, 躲避天敌的区域。鱼群会被鱼礁吸引, 然后逐渐形成人工生态系统。
在开放的环境中投放鱼礁将会改变原来稳定的流场, 进而促进水体交换。鱼礁的尺寸和结构的多样性导致不同地区的鱼礁附近的流场不同。Vicente等(2008)对部署在不同地区的人工鱼礁系统进行营养循环和海底有机物富集方面的环境评估。事实上水体交换不仅可以为鱼类提供不同的流体环境, 还可以促进营养盐的输运。因此, 从修复海洋环境到丰富沿海生态系统, 人工鱼礁都是岩礁鱼类的理想栖息地(Haro et al, 2004; Lan et al, 2004)。
投放人工鱼礁会影响鱼礁区域的流速分布, 并产生不容忽视的湍流。研究人工鱼礁对鱼礁区域影响的方法主要分为两种: 物理模型试验和数值模拟。早期研究人员使用物理模型试验对人工鱼礁附近流场形态变化进行初步分析。此后, 许多研究采用复杂的流体动力学模型, 通过数值模拟来探究流速的变化和涡流效应。在投放人工鱼礁前, 通过数值模拟对鱼礁投放后产生的流场效应进行分析, 可以节省费用以及时间, 但模拟结果的可靠性需要验证。Liu等(2006)利用有限体积法(finite volume method, FVM)对鱼礁附近流场进行了数值研究。Su等(2007)利用粒子成像测速仪(particle image velocimetry, PIV)对鱼礁流场进行测量与分析, 并对Liu等(2006)的数值结果进行了进一步的比较和验证。Su等(2008)还研究了海底地形对人工鱼礁内外流场的影响。Jiang (2010)利用(renormalization group theory, RNG) k-ε湍流模型和水槽实验模型对单体鱼礁产生的流场进行分析, 得出模型结果可以代替试验结果。
经鱼礁提高的渔业资源的生态效率在一定程度上取决于人工鱼礁的流场效应, 因此, 流场分析是研究人工鱼礁生态效率的关键。上升流和背涡流的规模成为众多研究中衡量鱼礁流场效应的主要标准。湍流的存在对于海洋上混合层的物理过程有着非常重要的影响。然而多数研究(Su et al, 2008; Liu et al, 2013; Li et al, 2017)并未分析湍流对于鱼礁区域流场形态的影响。湍流在不同尺度上对浮游生物产生影响(Kiørboe, 1993)。在近岸海域湍流能够增加营养盐通量(Dade, 1993)。一些研究数据表明湍流(即混合)与浮游植物群落组成之间存在联系(Prairie et al, 2012; Acevedo-Trejos et al, 2015)。湍流可能弥补了它们缺乏自我推进的器官的不足, 有利于它们遇到营养物质, 以及在真光层的持续存在(Margalef, 1978)。混合能够增强底部营养盐的扩散, 同时可能改变浮游植物的光照条件。本文将通过使用并行大涡模拟模式(the parallelized large-eddy simulation model, PALM)对鱼礁区域的湍流动能收支、流场形态进行分析。此外, 文章引入营养盐模型, 以探究投放鱼礁后的示踪物分布变化。
1 模型设计 1.1 模式介绍本文使用的PALM基于Boussinesq近似, 经过滤波的不可压缩的非流体静力学Navier-Stokes方程进行求解。其基本方程如下:
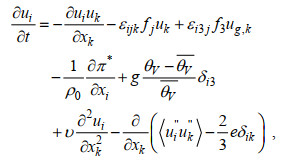 (1)
(1)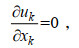 (2)
(2)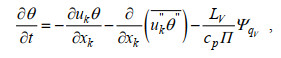 (3)
(3)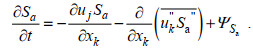 (4)
(4)由于营养盐与被动示踪物在水体中的运动特性相似, 故将PALM中的被动示踪物视为研究区域中的营养盐, 其控制方程如下:
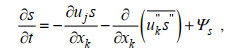 (5)
(5)其中, 上划线表示滤波后去除次网格项的值; 双撇号表示次网格尺度(subgrid-scale, SGS)变量; 尖括号表示SGS通量; 表 1详细列举了方程的参数说明。上述方程通过离散笛卡尔网格上的控制方程对变量求平均值, 可以实现分辨尺度和SGS湍流模式的隐式分离, 其中采用Deardorff(1980)提出的方法建立SGS, 为方便起见, 本文使用了方程的连续形式。模式的详细介绍见Huq等(2018)与Maronga等(2015)。
表 1 参数列表 Tab. 1 List of parameters
| 符号 | 单位 | 说明 |
| t | s | 时间 |
| i, j, k | / | i, j, k ∈ {1, 2, 3} |
| ui | m/s | 速度分量(u1 = u, u2 = v, u3= w) |
| xi | m | 笛卡尔网格坐标xi(x1= x, x2= y, x3= z) |
| ε | / | 列维-奇维塔符号 |
| fi | s–1 | 科氏力参数 |
| ρ0 | kg/m3 | 海水密度 |
| π* | hPa | 修改后的扰动压力 |
| θ | k | 位势温度 |
| θV | k | 虚拟位势温度 |
| υ | / | 运动黏度系数 |
| δi3 | / | 克罗内克符号 |
| e | m2/s2 | 次网格湍流动能 |
| qv | kg/kg | 湿度 |
| Sa | 盐度 | |
| S | kg/m3 | 示踪物浓度 |
| Ψqv | kg/(kg·s) | 湿度的源/汇项 |
| Ψs | kg/(m3·s) | 示踪物(营养盐)的源/汇项 |
| Π | / | 用于温度与位势温度之间的转换 |
本文通过计算鱼礁上方各深度的上升流通量(Jiang et al, 2020)来分析不同来流流速下鱼礁的抬升效果。
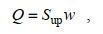 (6)
(6)其中, Sup表示上升流区域的面积; w表示上升流区域的垂向速度。若流场中垂向速度w大于或等于来流流速Uin的5%, 即w≥0.05Uin, 则可称该区域为上升流区域(Jiang et al, 2020)。
1.2 礁体模型与计算区域人工鱼礁结构繁多, 如箱体型、三角型、组合型等(杨吝等, 2005)。在我国的东南部沿海海域, 鱼礁多以米字型、立方体型为主(姜昭阳等, 2019)。在我国东部海域, 方型鱼礁已被广泛使用。李磊等(2018)通过研究箱体礁、三角形礁、框架礁对黑棘鲷的诱集作用, 发现箱体礁的诱集效果最佳。在近海区域, 付东伟等(2014)研究发现, 开口小的单体鱼礁产生的流场效应更好。林军等(2006)研究表明, 对于方型单礁来说, 其缓流区域体积与通透系数成反比。因此, 本次模拟选取边长L为3.0 m的实心方型人工鱼礁作为研究目标。赵海涛等(2006)认为, 由于波浪对鱼礁的冲击作用, 鱼礁应投放在水深20—30 m的海域。结合人工鱼礁实际案例(赵荣荣, 2019; 冯英明等, 2020), 本文的模拟区域水深为20 m。
为避免人工鱼礁流场遭受干扰(Ong et al, 2010), 本文将计算域的长度设置为100 m, 宽度设置为100 m。模型网格设置为200×200×80 (x×y×z), 网格大小设置为0.5×0.5×0.25 (m3), 即100 m×100 m×20 m (图 1)。参考Jiang等(2020)、Li等(2017)以及Liu等(2013)研究中, 模型的来流流速设置。本文设置四组理想实验, 其来流流速为0.1、0.2、0.4和0.6 m/s; 并设置对照组, 即区域不设置鱼礁, 其余的条件与实验组一致。入流边界条件为非循环边界条件; 表层、底层边界采用自由滑移边界条件。
 |
| 图 1 计算域示意图 Fig. 1 Computational domain |
曹欣中(1983)研究中的观测数据详细地展示了我国东部沿海海域表层营养盐各深度的分布情况。为使此次研究的模拟结果更具代表性, 本文参考曹欣中(1983)的观测结果, 设置了模拟区域各深度被动示踪物的浓度分布。被动示踪物浓度的分布见图 2。为与模型初始场使用的被动示踪物浓度观测数据的来源区域相对应, 此次研究设置的纬度为30°N。
 |
| 图 2 初始被动示踪物剖面 Fig. 2 Concentration of the passive scalar at the beginning time 注: S表示被动示踪物; Z表示深度; L代表鱼礁高度 |
鱼礁对营养盐的抬升作用依赖于鱼礁区域的垂向流场。在模式稳定时间内, 通过将鱼礁区的垂向速度进行水平(y)方向平均并与对照组相减得到图 3, 以研究鱼礁区域垂向速度与上升流区域的垂向分布。从图 3可以看出, 鱼礁的存在可以增大鱼礁前方的垂向速度。在垂向上, 垂向速度增大区域的面积随着来流流速的增大而增大。当来流流速达到0.6 m/s时, 该区域的高度可达到水体表面, 实现了对全水深流场的影响。不同来流流速对应的上升流部分在垂向上大体呈扇形且面积相近。当来流流速为0.2 m/s时, 鱼礁后方出现微弱的上升流, 其面积随来流流速的增大而增大。在鱼礁后方, 各来流流速对应的垂向流速方向均被迫向下, 形成背涡流区域。
 |
| 图 3 来流流速为0.1 (a)、0.2 (b)、0.4 (c)、和0.6 m/s (d)时, 实验组与对照组的垂向速度w在x-z方向上的差值 Fig. 3 Difference between vertical velocity w component of the experimental grip and that of the control group in the x-z profile, under different inflow velocities of 0.1 (a), 0.2 (b), 0.4 (c), and 0.6 m/s (d) 注: 灰线包裹区域为上升流区域; 黑色方块表示鱼礁; X代表模拟区域长度; Z表示深度; L代表鱼礁高度 |
图 4展示了来流流速为0.2 m/s时, 不同深度下(-6.00L、-5.67L、-5.33L、-5.00L、-4.67L、-4.33L), 鱼礁区的垂向速度与上升流区域在水平方向的分布。从图 4a可以看出, 上升流区域位于鱼礁侧后方且面积较小。对比图 4a—4f可知, 随着深度的变浅, 上升流区域的水平分布面积在垂向上先增大后减小。鱼礁后方垂向速度增大的部分与减小的部分相互脱离。在水平方向上, 鱼礁上方的上升流部分形状为圆形, 中心处的垂向速度最大, 越靠近外围, 垂向速度越小。
 |
| 图 4 来流流速为0.2 m/s时, 实验组与对照组在深度为-6.00L (a)、-5.67L (b)、-5.33L (c)、-5.00L (d)、-4.67L (e)、-4.33L (f)处, 垂向速度w在x-y方向上的差值 Fig. 4 Difference between vertical velocity w component of the experimental group and that of the control group in the x-y profile at different depths [-6.33L (a), -5.67L (b)、-5.33L (c)、-5.00L (d)、-4.67L (e)、-4.33L (f)] under the inflow velocity 0.2 m/s 注: 灰线包裹区域为上升流区域; 黑色方块表示鱼礁; Y代表模拟区域宽度; X代表模拟区域长度; L代表鱼礁高度 |
为进一步探究上升流区域的规模与强度, 文章通过使用参数Wa、Wmax、Hup、Sup对上升流区域的规模和强度进行描述。上升流最大速度(Wmax)、上升流平均速度(Wa)被用来形容上升流的强度; 上升流高度(Hup)、上升流面积(Sup)被用来形容上升流的规模。
为了使结论具有代表性, 增加两组来流流速(0.3、0.5 m/s)。从图 5a中可以看出, 上升流的最大速度、平均速度均与来流流速(Uin)成线性关系, 其线性关系式分别为Wmax=0.71Uin、Wa=0.08Uin, 相关系数均为1.00。从图 5b中可以看出, 随着来流流速的增大, 上升流的高度基本不变, 均在2.42L (7.25 m)附近。因此, Hup与Uin相关性不大。上升流面积与来流流速的关系可以用指数回归多项式进行表示。表达式为Sup=0.0013e14.1Um+32.92, 相关系数为0.97。
 |
| 图 5 上升流平均速度Wa及最大速度Wmax, 上升流高度Hup及面积Sup与来流流速Uin之间的关系 Fig. 5 The relationships of inflow velocities with average and maximum upwelling velocities, height and area of upwelling current region |
除了关注上升流区域与垂向速度在垂向以及水平方向上的分布之外, 文章还关注了鱼礁产生的上升流通量。本文选取鱼礁上方不同高度来分析各来流流速对应的上升流通量Q变化。
从图 6中可以看出, 鱼礁上方, 各来流流速对应上升流通量在垂向上的变化趋势相近, 即随着深度的增大, Q先增大后减少, 与(Jiang et al, 2020)研究结果类似。随着来流流速的增大, 鱼礁附近的上升流通量Q逐渐增大。图 4中的上升流区域垂向面积的变化趋势(先增大后减小)解释了上升流通量Q的在垂向的变化趋势。
 |
| 图 6 来流流速为0.1、0.2、0.4和0.6 m/s时, 上升流通量Q在不同深度处的分布 Fig. 6 Upwelling flux Q at different depths, under different inflow velocities of 0.1, 0.2, 0.4 and 0.6 m/s 注: Z表示深度; L代表鱼礁高度 |
根据公式(1)可得水平平均的SGS-TKE(turbulent kinetic energy)方程(Skyllingstad et al, 2000)为:
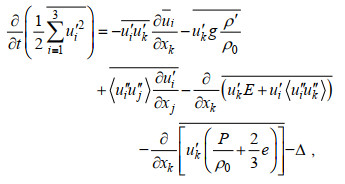 (7)
(7)其中,
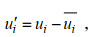 (8)
(8)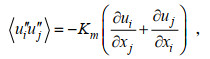 (9)
(9)P表示压强, Km表示次网格的涡黏系数, g表示重力加速度, E表示湍动能, Δ表示耗散项。公式(7)的等号右边, 第一项表示速度剪切项、第二项表示浮力项、第三项表示次网格耗散项、第四项和第五项分别表示湍流动能传输项和压力项、第六项表示耗散项。下文中湍流动能的耗散项包含次网格耗散, 即两项之和。
人工鱼礁的投放不仅会影响鱼礁区域的流场分布, 还会产生不容忽视的湍流。图 7展示了模型的湍流动能收支情况。根据公式(8), 本文用红线表示速度剪切项, 绿线表示湍流动能传输项, 蓝线表示压力项, 黑线表示耗散项。左图为四种来流流速下的海洋湍流动能收支情况, 右图均为左图底部区域的局部放大图。从图 7a、7c、7e、7g中可以发现, 海洋湍流动能收支各项的数值在深度为-5.5L处迅速增大, 然后迅速减小, 尤其是剪切项与耗散项的变化最为剧烈。此现象说明, 当来流遇到鱼礁后会产生强烈的湍流, 进而增强混合。在图 7b中, 压力项与湍流动能传输项的值在近海底处差别不大, 耗散项的最小值可以达到-1.14×10–6 m2/s3; 在图 7d中, 剪切项与耗散项绝对值的最大值位置接近海底且耗散项的数值较大, 约为4.52×10–6 m2/s3。在图 7f中, 耗散项在-6.33L处达到最小值, 最小值为-1.89×10–5 m2/s3, 在图 7h中, 对比剪切项和耗散项绝对值的最大值可知, 耗散项约为剪切项的1.5倍, 即4.46×10–5 m2/s3。
 |
| 图 7 来流流速为0.1 (a, b)、0.2 (c, d)、0.4 (e, f)、和0.6 m/s (g, h)时水体底部的湍流动能收支 Fig. 7 The distribution of TKE (turbulent kinetic energy) budget at the bottom of the research domain under different inflow velocities of 0.1 (a, b), 0.2 (c, d), 0.4 (e, f), and 0.6 m/s (g, h) 注: 右图均为左图底部区域的局部放大图; Z表示深度; L代表鱼礁高度 |
随着来流流速的增加, 在近海底处的剪切、传输与压力项必然增大, 导致向上传递的距离越远, 所产生的耗散也相应增大, 且随着深度减小逐渐减弱。
2.3 被动示踪物 2.3.1 被动示踪物分布为验证人工鱼礁产生的上升流对被动示踪物是否有抬升作用, 将实验组减去对照组的被动示踪物浓度得到图 8, 以探究鱼礁被动示踪物在垂向上的抬升效果。图 8a—8d中人工鱼礁后方, 接近底部的被动示踪物浓度为负值, 高于人工鱼礁的部分被动示踪物浓度为正值, 说明相较于未放置人工鱼礁的对照组, 在人工鱼礁的抬升作用下, 底部更多的被动示踪物进入上层水体。在人工鱼礁上部, 被动示踪物的抬升区域与上升流区域在垂向上均成扇形。
 |
| 图 8 来流流速为0.1 (a)、0.2 (b)、0.4 (c)、和0.6 m/s (d)时, 实验组与对照组在x-z方向的被动示踪物浓度S的差值 Fig. 8 Difference between the concentration of passive scalar S of the experimental group and that of the control group in the x-z profile, under different inflow velocities of 0.1 (a), 0.2 (b), 0.4 (c), and 0.6 m/s (d) 注: 灰线包裹区域为上升流区域; 黑色方块表示鱼礁; X代表模拟区域长度; Z表示深度; L代表鱼礁高度 |
各来流流速对应的被动示踪物抬升区域, 在人工鱼礁后方的不同位置处连续性明显减弱且抬升高度降低。造成此现象的原因为, 在距人工鱼礁一段距离后, 流场更加紊乱且垂向速度为负。
图 9展示了来流流速为0.2 m/s时, 不同深度下(-6.00L、-5.67L、-5.33L、-5.00L、-4.67L、-4.33L), 被动示踪物的水平分布。从图 9a可以看出, 受到在鱼礁的抬升作用, 接近鱼礁底部的营养盐被水体裹挟进入上层水体, 使得鱼礁后方的营养盐浓度明显降低。从图 9b—9f中可以看出距离中心越远的位置, 营养盐的抬升效果越弱。在水平方向上, 相较于图 4中上升流面积的变化趋势, 鱼礁上方营养盐的抬升面积不随深度的变化而变化。
 |
| 图 9 来流流速为0.2 m/s时, 实验组与对照组在深度为-6.00L (a)、-5.67L (b)、-5.33L (c)、-5.00L (d)、-4.67L (e)、-4.33L (f)处, 被动示踪物S浓度在x-y方向上的差值 Fig. 9 Difference between the concentration of passive scalar S of the experimental group and that of the control group in the x-y profile at different depths [-6.00L (a)、-5.67L (b)、-5.33L (c)、-5.00L (d)、-4.67L (e)、-4.33L (f)] under the inflow velocity 0.2 m/s 注: 灰线包裹区域为上升流区域; 黑色方块表示鱼礁; Y代表模拟区域宽度; X代表模拟区域长度; L代表鱼礁高度 |
为进一步探究鱼礁对被动示踪物的抬升效果, 在人工鱼礁上方0L以及后方5L、10L、15L处, 将实验组的被动示踪物浓度与对照组相减得到图 10。随着深度的加深, 被动示踪物的抬升效果基本呈现先缓慢增大后迅速减小的趋势。
 |
| 图 10 来流流速为0.1 (a)、0.2 (b)、0.4 (c)、和0.6 m/s (d)时, 在距人工鱼礁0L (黑线)、5L (黄线)、15L (蓝线)、20L (红线)处, 实验组与对照组的被动示踪物S浓度的差值 Fig. 10 Difference between the concentration of passive scalar S of the experimental group and that of the control group at 0L (black line), 5L (yellow line), 15L (blue line), and 20L (red line) from reef, under different inflow velocities of 0.1 (a), 0.2 (b), 0.4 (c), and 0.6 m/s (d) |
图 10a—10d中, 各来流流速对应的被动示踪物浓度分布在鱼礁上方十分接近。在距离鱼礁5L处, 随着来流流速的增大, 被动示踪物浓度有所降低。在距离鱼礁10L处, 来流流速为0.1、0.2、0.4 m/s时对应被动示踪物浓度不断降低; 当来流流速达到0.6 m/s时, 被动示踪物的抬升作用明显增强, 相较于来流流速为0.4 m/s, 抬升的被动示踪物浓度增大约2倍左右。在距离鱼礁15L处, 随着来流流速的增大, 被动示踪物的抬升作用先减弱后增强。对比图 10a—10d中的示踪物分布, 可以发现当来流流速大于0.2 m/s时, 鱼礁的抬升效果有明显的提升, 与图 8中被动示踪物抬升区域的连续性相符合。
2.3.2 被动示踪物通量文章还通过计算鱼礁上方的上升流区域对应的被动示踪物通量Fup来衡量人工鱼礁的抬升效果。
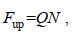 (10)
(10)其中, Q表示上升流通量; N表示上升流区域Sup所含被动示踪物浓度。研究选取鱼礁上方不同高度来分析各来流流速对应上升流区域的被动示踪物通量的Fup变化。
从图 11中可以看出, 鱼礁上方, 各来流流速对应被动示踪物通量与上升流通量变化趋势相近。随着来流流速的增大, 鱼礁附近的被动示踪物通量Fup逐渐增大。
 |
| 图 11 来流流速为0.1、0.2、0.4和0.6 m/s时, 上升流区域对应的营养盐通量Fup在不同深度处的分布 Fig. 11 Nutrient flux Fup of upwelling current region at different depths, under different inflow velocities of 0.1, 0.2, 0.4 and 0.6m/s 注: Z表示深度; L代表鱼礁高度 |
黄远东等(2012)利用计算流体力学(computational fluid dynamics, CFD)技术研究不同流速下, 方型鱼礁对应的上升流和背涡流的分布情况, 发现方型鱼礁的上升流高度和背涡流区宽度与来流流速无关; 唐衍力等(2017)研究进一步发现, 不同人工鱼礁结构的上升流及背涡流面积的大小与来流流速不相关, 最大上升流流速与来流流速呈线性关系。于定勇等(2019)发现随着鱼礁开口比的增大, 礁体产生上升流范围以及垂向速度逐渐减少。
与唐衍力等(2017)研究结果不同, 图 5中上升流面积与来流流速的关系为指数递增。由于本文将垂向速度w大于或等于来流流速Uin的5%的区域定义为上升流区域, 而唐衍力等(2017)则将垂向流速分量与来流流速之比等于或大于10%的区域定义为上升流区域。因此, 作者按照唐衍力等(2017)中的上升流的定义分别对上升流最大速度(Wmax)、上升流平均速度(Wa)及上升流高度(Hup)、上升流面积(Sup)进行统计得到图 12。图 12a中可以看出上升流的最大速度、平均速度均与来流流速成线性关系, 其线性关系式分别为Wmax=0.71Uin、Wa=0.15Uin、相关系数均为1.00。从图 12b中可以看出随着来流流速的增大, 上升流区域高度以及上升流区域面积基本不变, 均在2.42L(7.25m)以及1.44L2(13m2)附近, 与唐衍力等(2017)的研究结果一致。因此, 两次研究结果出现差异的原因为对于上升流的定义不同。
 |
| 图 12 改变上升流的定义后, 上升流平均速度Wa及最大速度Wmax, 上升流高度Hup及面积Sup与来流流速Uin之间的关系 Fig. 12 The relationships of inflow velocities with average and maximum upwelling velocities, height and area of upwelling current region at different definitions of upwelling changes |
图 3与图 4表明, 在投放鱼礁后, 鱼礁附近的垂向流速明显增大, 从而产生上升流。文章通过引用两种定义方法(唐衍力等, 2017; Jiang et al, 2020) 对上升流区域进行描述, 分别得到图 5以及图 12。由于上升流区域的最大速度与上升流区域的定义无关, 因此, 对比图 5与图 12可知, 在两种不同的上升流定义下, 除了上升流区域的最大速度(Wmax)相同之外, 上升流区域的平均速度(Wa)、高度(Hup)以及面积(Sup)的数值均不相同。值得注意的是, 两种上升流定义对应的上升流的最大流速(Wmax)及平均流速(Wa)与来流流速均成线性增加的关系, 上升流的高度(Hup)均不随来流流速的改变而改变, 随着来流流速的增大, 上升流区域面积(Sup)的变化趋势却完全不同。
3.2 被动示踪物的抬升效果在图 3中, 鱼礁后方存在明显的背涡流区域, 且随着来流流速的增大, 鱼礁产生背涡流随之增强。这就使得鱼礁对示踪物的抬升效果受到限制。当示踪物遇到鱼礁产生的上升流后, 会被迅速抬升至上层水体; 然后遇到鱼礁后方的背涡流, 示踪物的抬升受阻, 高度降低。因此, 从图 8中可以看出, 在垂向上各来流流速对示踪物的最大抬升高度基本相同, 鱼礁后方示踪物的抬升高度有所降低, 示踪物的抬升区域的面积基本相同。
在图 10中, 鱼礁底部对应各流速抬升的示踪物浓度均为负值, 即底部的示踪物被抬升到鱼礁上方。同时, 图 7中绿线代表的湍流传输项显示, 区域底部的传输项在垂向上的数值变化均为先负后正, 即底部的能量向上运输。此现象表明, 投置鱼礁后, 区域底部的动能被带往上层水体耗散。因此, 鱼礁的存在使得底部能量向上输运, 底层的示踪物也随之被带往上层, 体现了鱼礁对于投放海域底层的能量与营养物质均有抬升作用。
宁修仁等(2007)经过现场观测和取样, 得到珠江口冬季真光层深度大约为2.9±1.5 m。陈雨(2014)发现, 冬季长江口附近海域真光层深度在0.31—12.42 m之间变动, 且离岸越近, 真光层深度越小。从图 7中可以看出, 在近岸真光层范围内人工鱼礁对于营养盐均有抬升作用。
4 结论为研究不同来流流速下, 方型人工鱼礁对流场形态的影响以及对营养盐的抬升作用, 本文设置四种来流流速(0.1、0.2、0.4和0.6 m/s), 分析不同流速下的流场形态, 湍流动能收支各项的分布结果与被动示踪物的分布状态, 并对比有无人工鱼礁对被动示踪物抬升效果的差别。主要结论如下:
(1) 对于不同流速, 鱼礁上方上升流通量随着深度的变浅, 其变化趋势均为先增大后减小; 随着来流流速的增大, 鱼礁上方的上升流通量随之增大; 鱼礁产生的上升流区域均呈扇形。鱼礁的存在, 不仅使得其附近垂向速度增大, 还使其后方垂向速度减小。文章引用两种定义方法(唐衍力等, 2017; Jiang et al, 2020) 分别对上升流区域进行描述。结果发现, 两种上升流定义对应的上升流区域的高度(Hup)均不随来流流速的改变而改变, 上升流的最大流速(Wmax)及平均流速(Wa)与来流流速均成线性增加的关系, 然而随着来流流速的增大, 上升流区域面积(Sup)的变化趋势却完全不同。
(2) 受到上升流的抬升作用, 鱼礁底部的示踪物迅速进入海洋上层, 而后遇到鱼礁后方的背涡流, 示踪物的抬升受阻, 高度逐渐降低。上升流区域以及背涡流区域的共同影响下, 示踪物抬升区域的最大高度与来流流速无关; 且在鱼礁后方, 示踪物抬升区域的高度有所降低。各流速下, 鱼礁上升流区域的营养盐通量的变化趋势与上升流通量相似。随着与鱼礁的距离增大, 鱼礁的抬升作用先增强后减弱; 鱼礁上方的抬升效果最佳。在近海区域的真光层, 鱼礁可以实现对营养盐的抬升作用。
(3) 文章简略探讨了鱼礁产生湍流对海洋上层的影响。剪切项、压力项以及传输项为区域底部湍流动能的生成源, 平衡增大的耗散项主要靠剪切项。投放鱼礁后, 鱼礁区域底部的混合增强, 同时湍流剪切项与耗散项显著增大。湍流传输项的分布显示区域底部的能量被输运至上层, 这说明, 鱼礁的存在不仅对营养物质具有抬升作用, 还能将能量向上层运输; 此外, 它引起的上层混合也能使浮游生物的分布等进行重新分配, 进而形成特殊的生态区域。
本文通过使用并行大涡模拟模式PALM及其被动示踪物模块, 初步探索了鱼礁区域的流场形态与湍流过程对示踪物空间分布的影响。潮流在我国近海十分显著, 对近岸海洋环境的水动力过程影响较大, 未来工作重点便是在模式中加入周期性的潮流动力环境, 考虑潮流作用对鱼礁区域的影响。
参考文献
于定勇, 杨远航, 李宇佳. 2019. 不同开口比人工鱼礁体水动力特性及礁体稳定性研究. 中国海洋大学学报, 49(4): 128-136 |
付东伟, 陈勇, 陈衍顺, 等. 2014. 方形人工鱼礁单体流场效应的PIV试验研究. 大连海洋大学学报, 29(1): 82-85 |
冯英明, 许丙彩, 郝义, 等. 2020. 日照市海洋牧场示范区人工鱼礁选址适宜性分析. 山东国土资源, 36(1): 44-50 |
李磊, 陈栋, 彭建新, 等. 2018. 3种人工鱼礁模型对黑棘鲷的诱集效果研究. 海洋渔业, 40(5): 625-631 DOI:10.3969/j.issn.1004-2490.2018.05.013 |
杨吝, 刘同渝. 2005. 我国人工鱼礁种类的划分方法. 渔业现代化, (6): 22-23, 25 DOI:10.3969/j.issn.1007-9580.2005.06.006 |
陈雨, 2014. 长江口邻近海域冬季水体漫衰减特性及遥感反演初探. 上海: 华东师范大学硕士学位论文, 41-42
|
林军, 章守宇. 2006. 人工鱼礁物理稳定性及其生态效应的研究进展. 海洋渔业, 28(3): 257-262 DOI:10.3969/j.issn.1004-2490.2006.03.015 |
姜昭阳, 郭战胜, 朱立新, 等. 2019. 人工鱼礁结构设计原理与研究进展. 水产学报, 43(9): 1881-1889 |
刘诚刚, 宁修仁, 蔡昱明, 等. 2007. 南海北部及珠江口细菌生产力研究. 海洋学报(中文版), (2): 112-122 DOI:10.3321/j.issn:0253-4193.2007.02.014 |
赵荣荣, 2019. 长岛挡浪岛人工鱼礁生态修复效果初步评价. 舟山: 浙江海洋大学硕士学位论文, 7-8
|
赵海涛, 张亦飞, 郝春玲, 等. 2006. 人工鱼礁的投放区选址和礁体设计. 海洋学研究, 24(4): 69-76 DOI:10.3969/j.issn.1001-909X.2006.04.008 |
唐衍力, 龙翔宇, 王欣欣, 等. 2017. 中国常用人工鱼礁流场效应的比较分析. 农业工程学报, 33(8): 97-103 |
黄远东, 姜剑伟, 赵树夫. 2012. 方型人工鱼礁周围水流运动的数值模拟研究. 水资源与水工程学报, 23(3): 1-3 |
曹欣中. 1983. 影响浙江近海营养盐分布主要水动力因子的探讨. 海洋科学. 1983, 7(2): 36-38 |
Acevedo-Trejos E, Brandt G, Bruggeman J et al, 2015. Mechanisms shaping size structure and functional diversity of phytoplankton communities in the ocean. Scientific Reports, 5(1): 8918 DOI:10.1038/srep08918 |
Ambrose R F, Anderson T W, 1990. Influence of an artificial reef on the surrounding infaunal community. Marine Biology, 107(1): 41-52 DOI:10.1007/BF01313240 |
Dade W B, 1993. Near-bed turbulence and hydrodynamic control of diffusional mass transfer at the sea floor. Limnology and Oceanography, 38(1): 52-69 DOI:10.4319/lo.1993.38.1.0052 |
Davis N, VanBlaricom G R, Dayton P K, 1982. Man-made structures on marine sediments: Effects on adjacent benthic communities. Marine Biology, 70(3): 295-303 DOI:10.1007/BF00396848 |
Deardorff J W, 1980. Stratocumulus-capped mixed layers derived from a three-dimensional model. Boundary-Layer Meteorology, 18(4): 495-527 DOI:10.1007/BF00119502 |
FAO, 2020. The state of world Fisheries and Aquaculture 2020. Rome: Food and Agriculture Organization of the United Nations.
|
García S, Leiva Moreno I, 2003. Global overview of marine fisheries. Conference on Responsible Fisheries in the Marine Ecosystem, 103-123
|
Grace R A, 2001. The factors and processes that influence artificial reef longevity. Marine Technology Society Journal, 35(2): 3-13 DOI:10.4031/002533201788001910 |
Haro A, Castro-Santos T, Noreika J et al, 2004. Swimming performance of upstream migrant fishes in open-channel flow: a new approach to predicting passage through velocity barriers. Canadian Journal of Fisheries and Aquatic Sciences, 61(9): 1590-1601 DOI:10.1139/f04-093 |
Huq S, De Roo F, Raasch S et al, 2019. Vertically nested les for high-resolution simulation of the surface layer in PALM (version 5.0). Geoscientific Model Development, 12: 2523-2538 DOI:10.5194/gmd-12-2523-2019 |
Jiang Z Y, Liang Z L, Tang Y L et al, 2010. Numerical simulation and experimental study of the hydrodynamics of a modeled reef located within a current. Chinese Journal of Oceanology and Limnology, 28(2): 267-273 DOI:10.1007/s00343-010-9228-6 |
Jiang Z Y, Liang Z L, Zhu L X et al, 2020. Effect of hole diameter of rotary-shaped artificial reef on flow field. Ocean Engineering, 197: 106917 DOI:10.1016/j.oceaneng.2020.106917 |
Kiørboe T, 1993. Turbulence, phytoplankton cell size, and the structure of pelagic food webs. Advances in Marine Biology, 29: 1-72 |
Lan C H, Chen C C, Hsui C Y, 2004. An approach to design spatial configuration of artificial reef ecosystem. Ecological Engineering, 22(4-5): 217-226 DOI:10.1016/j.ecoleng.2004.04.004 |
Li J, Zheng Y X, Gong P H et al, 2017. Numerical simulation and PIV experimental study of the effect of flow fields around tube artificial reefs. Ocean Engineering, 134: 96-104 DOI:10.1016/j.oceaneng.2017.02.016 |
Liu T L, Liou S, Chen D W et al, 2006. Numerical flow simulation for the deployment analysis of artificial reefs. in Proceedings of the International Offshore and Polar Engineering Conference 533-537
|
Liu Y, Zhao Y P, Dong G H et al, 2013. A study of the flow field characteristics around star-shaped artificial reefs. Journal of Fluids and Structures, 39: 27-40 DOI:10.1016/j.jfluidstructs.2013.02.018 |
Margalef R, 1978. Life-forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta, 1(4): 493-509 |
Maronga B, Gryschka M, Heinze R et al, 2015. The parallelized large-eddy simulation model (PALM) version 4.0 for atmospheric and oceanic flows: Model formulation, recent developments, and future perspectives. Geoscientific Model Development, 8(8): 2515-2551 DOI:10.5194/gmd-8-2515-2015 |
Ong M C, Utnes T, Holmedal L E et al, 2010. Numerical simulation of flow around a circular cylinder close to a flat seabed at high Reynolds numbers using a k-ε model. Coastal Engineering, 57(10): 931-947 DOI:10.1016/j.coastaleng.2010.05.008 |
Prairie J C, Sutherland K R, Nickols K J et al, 2012. Biophysical interactions in the plankton: a cross-scale review. Limnology and Oceanography: Fluids and Environments, 2(1): 121-145 DOI:10.1215/21573689-1964713 |
Seaman W Jr, 2000. Artificial reef evaluation: with application to natural marine habitats. London: CRC Press, 1-249
|
Skyllingstad E D, Smyth W D, Crawford G B, 2000. Resonant wind-driven mixing in the ocean boundary layer. Journal of Physical Oceanography, 30(8): 1866-1890 DOI:10.1175/1520-0485(2000)030<1866:RWDMIT>2.0.CO;2 |
Su D T, Liu T L, Ou C H, 2007. A comparison of the PIV measurements and numerical predictions of the flow field patterns within an artificial reef. In: Proceedings of the 17th International Offshore and Polar Engineering Conference. Lisbon, Portugal: ISOPE, 2239-2245
|
Su D T, Liu T L, Ou C H, 2008. Numerical investigation into effects of seabed topography on flows in and around artificial reefs. Fisheries Science, 74(2): 236-254 DOI:10.1111/j.1444-2906.2008.01518.x |
Vicente M, Falcão M, Santos M N et al, 2008. Environmental assessment of two artificial reef systems off southern Portugal (Faro and Olhão): a question of location. Continental Shelf Research, 28(6): 839-847 DOI:10.1016/j.csr.2007.12.009 |
Walker B K, Henderson B, Spieler R E, 2002. Fish assemblages associated with artificial reefs of concrete aggregates or quarry stone offshore Miami Beach, Florida, USA. Aquatic Living Resources, 15(2): 95-105 DOI:10.1016/S0990-7440(02)01154-3 |