 2021, Vol. 52
2021, Vol. 52中国海洋湖沼学会主办。
文章信息
- 田园, 金燕, 陈炜, 李翠翠, 王璠, 于欣, 车宗豪, 刘括, 梁腾, 霍忠明, 闫喜武. 2021.
- TIAN Yuan, JIN Yan, CHEN Wei, LI Cui-Cui, WANG Fan, YU Xin, CHE Zong-Hao, LIU Kuo, LIANG Teng, HUO Zhong-Ming, YAN Xi-Wu. 2021.
- 菲律宾蛤仔(Ruditapes philippinarum)“斑马蛤2号”筏式和底播养殖模式比较研究
- COMPARISON IN GROWTH AND NUTRITION OF NEW MANILA CLAM BREED "ZC Ⅱ" CULTURED IN RAFT HANGING AND BOTTOM SOWING MODELS
- 海洋与湖沼, 52(6): 1496-1505
- Oceanologia et Limnologia Sinica, 52(6): 1496-1505.
- http://dx.doi.org/10.11693/hyhz20210500125
文章历史
-
收稿日期:2021-05-31
收修改稿日期:2021-09-03




2. 大连海洋大学 水产养殖学国家级实验教学示范中心 大连 116023
2. Dalian Ocean University, National Demonstration Center for Experimental Aquaculture Education, Dalian Ocean University, Dalian 116023, China
菲律宾蛤仔(Ruditapes philippinarum)属于广温、广盐性的埋栖型贝类, 原产于亚洲太平洋和印度洋沿岸, 在我国辽宁、河北、广东、福建等南北沿海均有分布。1936年由日本被引种到北美洲加拿大的不列颠哥伦比亚省, 随后又从北美相继引种到法国、英国、意大利、西班牙和土耳其等欧洲沿海地区, 现已成为世界性养殖贝类(Ghiselli et al, 2012)。菲律宾蛤仔是我国四大养殖贝类之一, 养殖产量占世界的90%以上, 具有养殖风险小、投资少、相对成本低、产量高的特点, 经济地位也越来越重要(闫喜武, 2005)。
我国菲律宾蛤仔主要养殖方式有滩涂养殖、池塘养殖和浅海底播养殖。滩涂养殖是指在近岸利用平整滩涂养殖菲律宾蛤仔。池塘养殖的主要方式为多营养级复合养殖(俗称池塘混养)模式(梁淼等, 2018), 是利用不同养殖生物间的生态位互补原理, 将滤食性(菲律宾蛤仔)、肉食性(对虾、蟹类、鱼类)和杂食性(鲻鱼、蓝子鱼)等多种营养级互补的养殖生物按合理密度放养, 通过水质调控等方法, 使人工投入物质得到综合利用, 从而提高养殖种类的存活和生长以及获得最佳产量。浅海底播养殖又称潮下带养殖, 该方式将菲律宾蛤仔的养殖水深由潮间带推进到潮下带30 m海域, 大幅度拓展了养殖空间, 缩短了养殖周期, 降低了产业风险, 提高了养殖效益, 现已广泛应用于鲍、海胆等品种的养殖(王波等, 1999; 杨红生等, 1999; 王兴章等, 2004; 林志强, 2006; 史洁等, 2010)。
随着菲律宾蛤仔养殖规模的逐渐扩大, 传统的养殖方式出现养殖成活率低, 回捕率低, 采捕破碎率高, 冬季采捕困难等问题。探索一种高效、环境友好型养殖方式是解决当前存在问题的主要途径之一。我国海岸线绵长, 港湾曲折, 饵料丰富, 环境多样化, 为贝类浅海筏式养殖提供了广阔空间。贝类筏式养殖具有管理方便、产量高、生长速度快等优点。因此, 近年来我国牡蛎、扇贝、贻贝、鲍、魁蚶等浅海筏式养殖发展迅猛(刘述锡等, 2013; 罗俊标等, 2016; 赵春暖等 2017; 欧洪来等, 2018; 谢玺等, 2020; 李华琳等, 2021), 为沿海经济发展做出了重大贡献。但目前国内外关于菲律宾蛤仔的筏式养殖尚无报道。
菲律宾蛤仔“斑马蛤2号”新品种(GS-01-007-2021)是以北方大连石河蛤仔为基础群体, 采用群体选育技术培育而成, 具有壳色美观, 生长快, 在北方滩涂和浅海越冬成活率高等特点。本研究结合当前菲律宾蛤仔养殖现状, 创新建立良种养殖推广模式, 在我国黄海北部首次开展了菲律宾蛤仔“斑马蛤2号”浅海筏式与底播养殖两种模式的对比试验, 通过两种养殖模式的生长、存活和营养成分比较, 探究菲律宾蛤仔筏式养殖的可行性, 对菲律宾蛤仔产品走向高端市场, 产业提质增效有重要意义。
1 材料与方法 1.1 样品采集于2018年秋季10月选择辽宁盘锦二界沟池塘混养培育的菲律宾蛤仔“斑马蛤2号”大规格苗种作为试验材料(图 1)。苗种平均壳长为(2.37±0.04) cm, 平均全湿重为(2.99±0.91) g。
 |
| 图 1 “斑马蛤2号” Fig. 1 "ZC Ⅱ", a new breed of R. philippinarum |
装卸避开中午高温时间, 干法运输至大连庄河王家岛, 运输途中严防暴晒、雨淋、风吹、机械损伤。运输温度为8 ℃, 运输时间为8 h以内。
1.2 方法 1.2.1 试验海区选择选择大连庄河王家岛海域进行菲律宾蛤仔“斑马蛤2号”筏式养殖和底播养殖对比试验。该海域风浪平静, 潮流畅通并有淡水注入的内湾浅海, 底质以砂泥底为主, 海流流速40— 100 cm/s, 养殖海区地势平坦、无工业污染。
1.2.2 养殖方法(1) 筏式吊养: 筏架采取垂直法设置, 延绳式吊漂法, 行距15 m。浮梗采用直径30 mm的聚乙烯绳, 每根梗绳100 m (加木橛腿), 有效挂笼长度80 m。2018年10月将菲律宾蛤仔“斑马蛤2号”大规格苗种装入多层聚乙烯网笼中, 养殖密度为150—200粒/层(m2), 每笼18层。将塑料网笼吊挂于设置好的海区浮筏, 每台筏悬挂60个网笼。将装有菲律宾蛤仔“斑马蛤2号”大规格苗种的塑料网笼挂于筏式养殖海区, 共挂养1 477笼。在菲律宾蛤仔“斑马蛤2号”筏式养殖期间进行水质指标监测与养殖管理维护, 后期随着附泥及附着物增多和苗种的体重增加, 及时增加浮漂以防筏架下沉。在7月份养殖期更换一次养殖笼, 清除网笼内外的附着物。
(2) 浅海底播养殖: 2018年10月放养苗种, 避开潮汐流速最大的时段。放养密度为530—600粒/m2。运苗船通过导航定位至放养区域, 底播养殖水深7—8 m。采用干法放苗, 均匀播撒。在菲律宾蛤仔“斑马蛤2号”浅海底播养殖期间进行水质指标监测与养殖管理, 使用拖网清除广大扁玉螺、香螺、多棘海盘车等敌害生物。
1.2.3 采样及测量分别于2019年冬季1月、春季4月、夏季7月、秋季11月采集浅海筏式养殖和底播养殖的“斑马蛤2号”样品。每个试验区随机确定3个采样点, 利用GPS进行定位, 筏式养殖3个采样点与底播养殖3个采样点见图 2与表 1。
 |
| 图 2 两种养殖方式的采样地点 Fig. 2 Sampling sites of the two breeding modes |
| 养殖方式 | 采样点 | 纬度(N) | 经度(E) |
| 筏式养殖(FC) | FC-1 | 39°30′227″ | 123°05′189″ |
| FC-2 | 39°30′241″ | 123°05′164″ | |
| FC-3 | 39°30′220″ | 123°05′143″ | |
| 底播养殖(SC) | SC-1 | 39°34′686″ | 123°21′089″ |
| SC-2 | 39°34′833″ | 123°21′162″ | |
| SC-3 | 39°34′542″ | 123°21′555″ |
采用游标卡尺(精确到0.01 mm)对两种养殖方式“斑马蛤2号”壳长、壳高、壳宽表型生长性状进行测量, 用电子天平(精确到0.01 g)称取“斑马蛤2号”全湿重及软体重。每次每个采样点选取30个样品进行测量。计算各个性状(壳长、壳宽、壳高、全湿重、软体重)每个季度的相对生长速率。相对生长速率简称RGR, 以R表示。增加一单位生物量所需整体生物量的增长速率, 即单位时间内蛤仔各性状的增长量。公式如下:
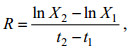 (1)
(1)式中, R代表相对生长速率(RGR), X2代表t2时刻蛤仔表型性状观测值, X1代表t1时刻蛤仔表型性状观测值。t单位为d, X单位为mm或g。
1.3 营养成分将每个季度采集到的菲律宾蛤仔“斑马蛤2号”低温运回辽宁省贝类良种繁育工程技术研究中心实验室。解剖样品后, 将软体部装入密封袋, 放置于–80 ℃保存待用。
1.3.1 一般营养成分测定(1) 水分的测定: 干燥法(国标GB 5009.3— 2010)。把培养皿放入105 ℃烘干箱中恒重时取出, 利用电子天平称取样品后放入105 ℃烘干箱中进行烘干至恒重(两次重量差不超过0.002 g即为恒重)。
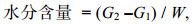 (2)
(2)式中, G1和G2分别为恒重后培养皿重量(g)和恒重后培养皿和样品总重(g), W表示样品重量(g)。
(2) 灰分的测定: 采用国标(GB 5009.4-2016)第一法。样品放置坩埚中称重, 坩埚放置马弗炉中550 ℃灼烧6 h并重复灼烧至恒重, 计算灰分含量。
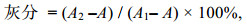 (3)
(3)式中, A1和A2分别为灼烧前坩埚和样品重(g)和灼烧完的坩埚和样品重(g), A表示坩埚重(g)。
(3) 粗蛋白的测定采用考马斯亮蓝法测定: 使用南京建成的蛋白测定试剂盒。
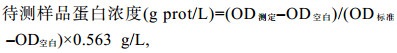 (4)
(4)样品处理: 称取4—5 g样品, 加入2% BHT-甲醇溶液, 采用氯仿-甲醇(体积分数2︰1)混合液超声震荡抽提脂质, 抽滤。先后加入0.88% KCl水溶液、水-甲醇混合液(体积分数1︰1), 充分震荡, 静置分层。用旋转蒸发仪减压蒸干氯仿, 底瓶恒重, 增重值为脂质含量。向含有脂质的蒸馏瓶中加入0.5 moL/L的KOH-无水甲醇混合液, 于70 ℃下完成回流水解。以BF3乙醚液为催化剂, 完成甲酯化。用沸程30—60 ℃的石油醚萃取脂肪酸甲酯, 挥发浓缩石油醚至适当体积, 用日本岛津公司的GC-2010型气相色谱仪测定脂肪酸。
色谱分析条件: FFAP石英交联毛细管色谱柱(30 m×0.25 mm×0.3 μm); 进样口260 ℃, 分流比100∶1; 流动相为高纯N2, 柱流量为1 mL/min, 初始柱温160 ℃, 升温速率2 ℃ /min, 终止温度230 ℃; FID检测器, 温度230 ℃。色谱峰的鉴定采用脂肪酸甲酯标准样品(SIGMA公司和上海试剂一厂)与ECL值相结合的方法, 采用面积归一化法定量。
1.3.3 糖原的测定使用苏州科铭公司生产的糖原测定试剂盒。
样品处理: 称取0.1—0.2 g组织样品, 置于10 mL试管/离心管中; 加入0.75 mL提取液, 沸水浴处理20 min (密封以防止水分散失), 每隔5 min振摇混匀1次; 待组织全部溶解后, 冷却至室温, 使用蒸馏水定容至5 mL, 取1 mL提取液在常温离心10 min, 取上清液即为待测样本。再根据试剂盒操作说明书的操作方法进行测定。
糖原含量计算公式如下:
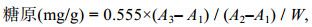 (5)
(5)测定两种养殖方式四个季度的水环境指标。采用水质分析仪(YSL proplus)现场测定温度(t, ℃)、溶氧(DO)、盐度(S)、pH。按照《海洋监测规范》测定三氮一磷及叶绿素a。
1.5 数据分析采用EXCEL2010软件对数据进行处理, 实验数据以“平均值±标准差(mean±SD)”表示; 用SPSS 16.0进行方差齐性分析, 并进行T检验与单因素方差分析(OneWay ANOVA), 显著性水平设置为P < 0.05。
2 结果 2.1 两种养殖模式四个季度生长及存活率比较浅海筏式养殖和底播养殖“斑马蛤2号”壳长、壳宽、壳高、全湿重、软体重及其相对生长速率见表 2和图 3。结果表明, 4—7月筏式养殖“斑马蛤2号”壳长、壳宽、壳高、全湿重、软体重相对生长速率比底播养殖“斑马蛤2号”快149.54%、100.61%、116.30%、152.25%、132.58%, 两者差异显著(P < 0.05)。但其他月份筏式养殖“斑马蛤2号”生长速率略慢于底播养殖“斑马蛤2号”(P < 0.05)。
| 采样日期 | 养殖方式 | 壳长(cm) | 壳宽(cm) | 壳高(cm) | 全湿重(g) | 软体重(g) |
| 冬季(1月) | SC | 2.74±0.19a | 1.14±0.11a | 1.93±0.17a | 3.94±0.82a | 1.29±0.30a |
| FC | 2.39±0.44b | 1.04±0.09b | 1.70±0.28b | 3.06±0.68b | 1.06±0.23b | |
| 春季(4月) | SC | 3.03±0.18a | 1.27±0.09a | 2.07±0.12a | 5.20±0.87a | 1.69±0.34a |
| FC | 2.45±0.26b | 1.03±0.10b | 1.69±0.17b | 2.91±0.92b | 1.08±0.37b | |
| 夏季(7月) | SC | 3.37±0.25a | 1.51±0.09a | 2.35±0.14a | 7.31±1.78aa | 2.13±0.70a |
| FC | 3.16±0.20b | 1.44±0.10b | 2.21±0.14b | 6.72±1.26bb | 1.80±0.40b | |
| 秋季(11月) | SC | 3.51±0.12a | 1.52±0.06a | 2.39±0.08a | 8.08±2.16a | 1.40±0.22a |
| FC | 3.18±0.29b | 1.51±0.10a | 2.26±0.15a | 7.61±1.61a | 1.37±0.48a | |
| 注: SC为底播养殖, FC为筏式养殖; 同一列含有不同字母表示差异显著(P < 0.05), 标有相同字母者表示组间无显著性差异(P > 0.05) | ||||||
 |
| 图 3 两种养殖方式“斑马蛤2号”壳长、壳宽、壳高、全湿重及软体重相对生长速率比较 Fig. 3 Comparisons in RGR, shell length, shell width, shell height, total wet weight, and soft body weight of the "ZC Ⅱ" in the two breeding modes 注: a. 壳长; b. 壳高; c. 壳宽; d. 全湿重; e. 软体重 |
筏式养殖“斑马蛤2号”存活率为90%以上, 且秋季和冬季显著高于底播养殖(P < 0.05; 图 4)。
 |
| 图 4 两种养殖方式“斑马蛤2号”存活率比较 Fig. 4 Comparison in survival rate of the "ZC Ⅱ" in two breeding methods 注: 同一列不同字母表示差异显著(P < 0.05), 相同字母表示无显著性差异(P > 0.05) |
两种养殖方式“斑马蛤2号”水分、灰分、粗蛋白、糖原变化如图 5。夏季7月筏式养殖“斑马蛤2号”水分显著高于底播养殖“斑马蛤2号”(P < 0.05), 其余月份两种养殖模式无显著差异(P > 0.05)。秋季11月筏式养殖“斑马蛤2号”灰分含量显著高于底播养殖“斑马蛤2号”(P < 0.05), 其余月份两种养殖模式之间无显著差异(P > 0.05)。四个季度两种养殖方式“斑马蛤2号”粗蛋白无显著差异(P > 0.05)。“斑马蛤2号”夏季7月糖原含量显著高于其他季节(P < 0.05), 但两种养殖模式间“斑马蛤2号”糖原含量差异不显著(P > 0.05)。秋季11月筏式养殖“斑马蛤2号”糖原显著高于底播养殖“斑马蛤2号”(P < 0.05)。
 |
| 图 5 两种养殖方式“斑马蛤2号”水分、灰分、糖原及蛋白含量随季节变化 Fig. 5 The seasonal variations in moisture, ash, glycogen and protein contents of the "ZC Ⅱ" in the two breeding methods 注: a. 水分含量; b. 灰分含量; c. 糖原含量; d. 蛋白含量 |
不同季节两种养殖方式下“斑马蛤2号”的脂肪酸组成和含量见表 3。所有样本中的脂肪酸种类和特点一致, 共检测出22种脂肪酸, 其中包含6种饱和脂肪酸(SFA)、3种单不饱和脂肪酸(MUFA)和13种多不饱和脂肪酸(PUFA)。含量较多的主要脂肪酸为C16:0、C16:1n-7、C18:0、C18:1、C20:0、C20:5n-3 (EPA)、C22:6n-3 (DHA), 且PUFA > SFA > MFUA。
| 月份 | 1月(冬季) | 4月(春季) | 7月(夏季) | 11月(秋季) | ||||||||
| 养殖模式 | 浮筏 | 底播 | 浮筏 | 底播 | 浮筏 | 底播 | 浮筏 | 底播 | ||||
| C14:0 | 3.52±0.02c | 4.06±0.08c | 2.20±0.22b | 1.94±0.07b | 1.90±1.23b | 1.71±0.42b | 0.76±0.13a | 0.66±0.08a | ||||
| C15:0 | 1.35±0.29bc | 1.57±0.27c | 1.01±0.11ab | 0.76±0.13a | 0.72±0.24a | 0.83±0.22a | 1.15±0.49abc | 0.86±0.03a | ||||
| C16:0 | 19.31±0.78b | 20.27±0.20b | 20.61±0.82b | 19.79±0.53b | 20.18±2.96b | 19.28±1.67b | 14.24±2.29a | 12.75±1.20a | ||||
| C16:1n-7 | 6.34±0.26d | 6.10±0.14cd | 6.03±0.32cd | 5.03±0.14bc | 4.12±1.59b | 4.07±0.45b | 2.35±0.33a | 2.58±0.15a | ||||
| C17:0 | 0.89±0.05a | 0.93±0.04ab | 0.83±0.06a | 0.84±0.05a | 1.17±0.26c | 1.12±0.11bc | 1.57±0.07d | 1.59±0.19d | ||||
| C17:1n-7 | 1.12±0.11ab | 0.98±0.09a | 0.72±0.11a | 0.82±0.06a | 2.82±1.08abc | 3.13±0.93bc | 4.23±2.03c | 6.36±2.09d | ||||
| C18:0 | 3.68±0.09a | 4.02±0.19ab | 4.28±0.25ab | 4.67±0.12b | 7.16±0.92c | 7.44±0.62cd | 7.74±0.28cd | 8.11±0.21d | ||||
| C18:1 | 6.34±1.05bc | 5.15±0.13ab | 6.76±0.57c | 6.14±0.14bc | 7.07±1.48c | 7.39±0.84c | 4.57±0.46a | 4.52±0.15a | ||||
| C18:2n-6 | 0.99±0.51bc | 0.73±0.09abc | 1.03±0.21bc | 1.07±0.24c | 0.76±0.36abc | 0.80±0.10bc | 0.51±0.27ab | 0.26±0.12a | ||||
| C18:3n-3 | 0.83±0.07c | 0.78±0.03bc | 0.84±0.06c | 1.09±0.16d | 0.74±0.30bc | 0.53±0.11b | 0.28±0.12a | 0.23±0.03a | ||||
| C18:4n-3 | 3.50±0.30b | 3.56±0.11b | 3.38±0.44b | 3.98±0.20b | 1.49±0.66a | 1.44±0.35a | 1.25±0.45a | 1.69±0.53a | ||||
| C20:0 | 3.98±0.49a | 3.68±0.17a | 4.56±0.47a | 3.91±0.73a | 7.47±0.95b | 8.29±0.57b | 8.11±0.35b | 8.07±0.07b | ||||
| C20:2n-6 | 1.05±0.05a | 1.00±0.01a | 1.25±0.07b | 1.32±0.05b | 1.86±0.23d | 1.84±0.13d | 1.52±0.09c | 1.41±0.05bc | ||||
| C20:4n-6 | 1.78±0.03a | 1.78±0.04a | 1.43±0.09a | 1.57±0.07a | 2.56±0.43b | 2.93±0.51b | 4.74±0.39c | 4.86±0.13c | ||||
| C20:4n-3 | 1.20±0.07c | 1.21±0.02c | 1.18±0.03c | 1.17±0.05c | 0.73±0.14b | 0.68±0.02b | 0.19±0.10a | 0.24±0.06a | ||||
| C20:5n-3(EPA) | 18.31±1.39f | 17.39±0.40f | 15.52±1.22e | 13.59±0.26d | 9.78±1.82c | 8.40±0.75bc | 5.93±0.36a | 6.79±0.19ab | ||||
| C22:2 | 2.06±0.05a | 2.05±0.13a | 2.31±0.38a | 2.38±0.06a | 3.84±1.54b | 4.44±0.85b | 7.91±0.61c | 6.88±0.77c | ||||
| C22:3n-6 | 1.74±0.08b | 1.72±0.07b | 1.63±0.07b | 1.67±0.04b | 1.06±0.35a | 0.98±0.10a | 1.13±0.04a | 0.90±0.07a | ||||
| C22:4n-6 | 0.38±0.01ab | 0.36±0.02ab | 0.32±0.04a | 0.34±0.03a | 0.66±0.20bc | 0.75±0.23c | 1.94±0.06d | 2.08±0.35d | ||||
| C22:5n-6 | 0.81±0.23bc | 0.59±0.04ab | 0.55±0.07a | 0.47±0.05a | 0.86±0.26c | 0.83±0.13bc | 1.82±0.03e | 1.24±0.08d | ||||
| C22:5n-3 | 1.28±0.18a | 1.25±0.04a | 1.59±0.12a | 1.57±0.07a | 1.69±0.18a | 1.82±0.20a | 1.90±0.13a | 2.87±1.47b | ||||
| C22:6n-3(DHA) | 15.20±1.21a | 16.00±0.55ab | 18.68±1.54bcd | 21.89±0.60d | 19.24±3.98bcd | 18.12±1.85abc | 19.65±0.84cd | 18.07±0.87abc | ||||
| 其他 | 4.34±0.58abc | 4.83±0.75bcd | 3.30±1.11ab | 3.98±1.64ab | 2.14±1.41a | 3.17±1.25ab | 6.52±2.12cd | 6.97±0.91d | ||||
| 总和 | 95.66±0.58bcd | 95.17±0.75abc | 96.70±1.11cd | 96.02±1.64cd | 97.86±1.41d | 96.83±1.25cd | 93.48±2.12ab | 93.03±0.91a | ||||
| ∑SFA | 32.73±0.68a | 34.52±0.44a | 33.49±0.75a | 31.92±1.37a | 38.60±2.32b | 38.67±1.65b | 33.57±2.44a | 32.04±1.62a | ||||
| ∑MUFA | 13.80±1.26ab | 12.24±0.35ab | 13.51±0.96ab | 11.99±0.18ab | 14.00±2.93ab | 14.59±1.76b | 11.14±1.75a | 13.46±2.03ab | ||||
| ∑PUFA | 49.14±2.49bcd | 48.42±0.97bcd | 49.70±2.47cd | 52.11±0.36d | 45.26±3.96ab | 43.57±2.45a | 48.77±1.46bcd | 47.53±0.55bc | ||||
| 注: SFA为饱和脂肪酸, MUFA为单不饱和脂肪酸, PUFA为多不饱和脂肪酸 | ||||||||||||
由表 3可见, 养殖模式对“斑马蛤2号”脂肪酸含量的影响较小, 除了4月份的C18:3n-3、EPA和11月份的C17:1n-7、C22:5n-6、C22:5n-3有显著差异之外(P < 0.05), 其他脂肪酸含量在两种养殖模式之间均无显著差异(P > 0.05)。
不同季节的“斑马蛤2号”脂肪酸含量存在一定差异。筏式和底播养殖“斑马蛤2号”DHA含量均在春季4月最高, 冬季1月最低, 且两者差异显著(P < 0.05); 夏季7月的SFA含量显著高于其他月份(P < 0.05); PUFA含量在春季4月最高, 且在相同养殖模式下春季4月的PUFA含量显著高于7月夏季(P < 0.05)。
2.3 两种养殖方式四个季度水环境因子特征分析对两种养殖方式四个季度的海水无机氮(亚硝酸盐、硝酸盐、氨盐)、活性磷酸盐、盐度、温度、pH和溶解氧数据监测结果见表 4。结果表明, 筏式养殖和底播养殖各水环境因子差异不显著(P > 0.05)。
| 时间 | 采样点 | 亚硝态氮(mg/L) | 氨氮(mg/L) | 硝态氮(mg/L) | 磷(mg/L) | 叶绿素a(μg/L) | 溶解氧(mg/L) | 盐度 | 温度(℃) | 酸碱度pH |
| 1月 | FC | 0.008 63 | 0.042 43 | 0.057 92 | 0.033 22 | 4.69 | 10.45 | 28.40 | 2.23 | 8.08 |
| SC | 0.006 69 | 0.040 22 | 0.058 54 | 0.035 06 | 3.64 | 10.34 | 28.20 | 1.77 | 8.08 | |
| 4月 | FC | 0.004 06 | 0.031 85 | 0.025 73 | 0.021 08 | 3.92 | 8.95 | 28.10 | 9.50 | 8.20 |
| SC | 0.003 98 | 0.035 05 | 0.027 15 | 0.022 90 | 3.59 | 9.50 | 28.13 | 8.67 | 8.16 | |
| 7月 | FC | 0.001 81 | 0.023 26 | 0.026 48 | 0.016 00 | 5.14 | 8.43 | 27.60 | 23.60 | 8.13 |
| SC | 0.002 13 | 0.019 33 | 0.022 24 | 0.012 07 | 5.62 | 9.37 | 25.93 | 27.67 | 8.21 | |
| 11月 | FC | 0.002 88 | 0.019 17 | 0.029 45 | 0.011 57 | 4.69 | 9.53 | 28.63 | 15.17 | 8.03 |
| SC | 0.003 68 | 0.016 19 | 0.027 52 | 0.009 85 | 5.32 | 10.53 | 27.23 | 14.20 | 8.12 | |
| 注: SC为底播养殖, FC为筏式养殖 | ||||||||||
海洋贝类主要有底播和筏式两种养殖方式, 其中贝类筏式养殖以其养殖种类生长速度快、产量高、管理方便等优点, 逐步发展成为海洋贝类养殖的主要生产方式, 并得到大面积推广(冯继兴等, 2016)。我国筏式养殖的贝类主要以扇贝、牡蛎、贻贝、鲍为主, 20世纪70年代贻贝养殖的规模化, 标志着我国浅海贝类养殖业的真正崛起。80年代海湾扇贝和虾夷扇贝的引进, 形成筏式养殖等方式, 有力地推动了我国海水养殖业的发展。
在本研究中, “斑马蛤2号”筏式养殖和底播养殖四个季度的海水养殖环境差异不显著的情况下, 筏式养殖的“斑马蛤2号”生长速率虽然略慢于底播养殖的“斑马蛤2号”, 但在夏季4—7月期间, 筏式养殖“斑马蛤2号”生长速度显著快于底播养殖蛤仔(P < 0.05), 筏式养殖“斑马蛤2号”夏季(4—7月)壳长、壳宽、壳高、全湿重、软体重的相对生长速率分别比底播养殖“斑马蛤2号”快149.54%、100.61%、116.30%、152.25%、132.58% (P < 0.05), 说明“斑马蛤2号”筏式养殖在夏季具有一定的优势。氮磷是养殖区浮游植物生长繁殖必要的营养要素。从表 4中可发现, 黄海海域的冬季海水中的无机氮含量最高, 夏季的叶绿素含量高于冬季, 这些海水环境为贝类提供了丰富的浮游植物, 不同季节浮游植物的分布以及丰度情况会直接影响蛤仔的生长速率。在以后的研究中, 应增加有机颗粒及水流速度等环境因子的测定, 进一步分析夏季浮筏养殖菲律宾蛤仔“斑马蛤2号”生长快的原因。
合理的养殖密度是贝类筏式养殖的关键要素。筏式养殖和底播养殖菲律宾蛤仔“斑马蛤2号”的生长、存活比较研究表明, 采用多层聚乙烯网笼(于当年10月), 按密度为150—200粒/m2放养大规格苗种(平均壳长为2.37 cm, 平均全湿重为2.99 g)的“斑马蛤2号”, 至翌年的4月、7月、11月可分别收获壳长为3.03 cm、3.36 cm、3.51 cm的“斑马蛤2号”。李华琳等(2021)在筏式养殖虾夷扇贝的调查过程中发现, 低密度养殖在壳高性状上具有优势, 王崇明等(2004)在栉孔扇贝养殖过程中也发现了养殖密度越大死亡率越高的问题。何毛贤等(2009)研究发现2 m水层筏式养殖的马氏珠母贝存活率最高, 指出养殖水层和养殖密度间具有显著的交互作用。于佐安等(2016)通过对1—2龄的虾夷扇贝筏式养殖研究分析发现春季到夏季附着生物增多, 浮筏养殖水交换受到了一定影响。双壳贝类的滤水能力非常强大(Ezgeta-Balić et al, 2012; Peharda et al, 2012; Galimany et al, 2017), 能摄食水体中多种悬浮颗粒, 养殖密度越高, 养殖区内饵料缺乏, 会导致贝类营养不足, 影响浅海生态系统物质及能量流动, 受到病原感染的机率增大, 死亡率高, 因此低密度养殖可降低贝类的死亡率(冯继兴等, 2016; 于佐安等, 2019)。在本研究中, 四个季度筏式养殖“斑马蛤2号”存活率为90%以上。冬季1月与秋季11月, 筏式养殖的菲律宾蛤仔“斑马蛤2号”存活率明显高于底播养殖(P < 0.05), 其余月份两种养殖方式无明显差异(P > 0.05)。本研究表明两种养殖方式四个季度的海水无机氮(亚硝酸盐, 硝酸盐, 氨盐)、活性磷酸盐、盐度、温度、pH和溶解氧等水环境因子差异不显著(P > 0.05), 从水环境因子分析中未发现两种养殖方式存活率出现差异的主要原因。分析两种养殖方式“斑马蛤2号”存活率出现差异的原因, 一方面可能与采捕方式有关, 筏式养殖中采用聚乙烯网笼养殖“斑马蛤2号”, 在采捕中由于机械损伤造成的破碎率低于1%; 但底播养殖“斑马蛤2号”是采用底拖网采捕, 由于机械损伤造成的破碎率为5%—10%; 另一方面可能与敌害捕食有关, 尽管夏季在“斑马蛤2号”底播养殖区会进行敌害清理, 但敌害清除率一般为50%—70%, 仍旧有部分扁玉螺等敌害生物捕食“斑马蛤2号”, 造成养殖损失。而筏式养殖“斑马蛤2号”受敌害捕食影响很小。
3.2 两种养殖方式“斑马蛤2号”营养成分比较脂肪酸具有多种功能, 在人体健康中发挥重要作用(Behrens et al, 1996; Lopez-Huertas, 2010; 段叶辉等, 2014; 李锦灵等, 2021)。贝类营养丰富, 富含多不饱和脂肪酸(PUFA), EPA和DHA (焦莉等, 2020), 具有较高的开发利用价值。
本研究两种养殖方式营养成分研究结果表明, 除了个别脂肪酸含量有明显差异之外(P < 0.05), 其他脂肪酸含量在两种养殖模式之间均无显著差异(P > 0.05), 说明不同的养殖模式对菲律宾蛤仔中脂肪酸含量影响不大, 这与陈炜等(2018)对不同养殖模式下岩扇贝体内脂肪酸含量的研究结果基本一致。但不同的是4月EPA含量浮筏养殖高于底播养殖。菲律宾蛤仔为滤食性贝类, 主要摄食细小的浮游植物和有机碎屑(乔方利等, 2000), 而饵料种类及密度直接影响蛤仔的脂肪酸组成和含量(Zhu et al, 2020)。浮游植物丰度在渤海区的垂直分布趋势随水深先增加后降低(郭术津等, 2014), 因此春季浮筏养殖比底播养殖的“斑马蛤2号”EPA含量高可能与不同水层中的饵料种类及密度有关。
不同季节对“斑马蛤2号”脂肪酸含量影响较大, 贝类体内脂肪酸含量受物种、温度、饵料、季节等因素的影响。在两种养殖模式下DHA含量均在冬季1月含量最低, 春季4月最高, 且两者差异显著(P < 0.05), 冬季1月的EPA含量显著高于其他季节, 杨东敏等(2017)对越冬期前后菲律宾蛤仔营养成分变化研究中发现, SFA含量在越冬期后显著降低, 而越冬期后的EPA含量高于越冬期前, 与本实验结果相似。在相同养殖模式下4月春季的PUFA含量高于其他月份(P < 0.05), 7月夏季SFA含量最多(P < 0.05)。不同季节的脂肪酸含量差异可能与不同季节的海水温度差异有关。元冬娟等(2009)对六种贝类冬夏两季的脂肪酸研究中发现, 冬季的PUFA含量高于夏季与本研究结果相似。但在本研究中的夏季“斑马蛤2号”SFA含量高于冬季, 与元冬娟等(2009)实验结果不同, 推测是由于不同贝类营养成分差异而产生的。
3.3 筏式蛤仔养殖的经济价值蛤仔筏式养殖成本投入包括梗绳、木橛、浮漂、聚乙烯绳、养殖笼、苗种费、养殖船只养护费、人工费等。蛤仔养殖商品规格为50粒/500 g, 每笼可产商品蛤30 kg, 每笼产值约180—200元。
4 结论本试验结果表明, 大规格菲律宾蛤仔“斑马蛤2号”苗种(平均壳长为2.37 cm, 平均全湿重为2.99 g)经筏式养殖可达到市场规格。“斑马蛤2号”筏式养殖模式具有存活率高, 冬季糖原、单不饱和脂肪酸含量高于底播养殖的优点。“斑马蛤2号”筏式养殖模式试验的成功对蛤仔产品走向高端市场, 产业提质增效具有重要意义。
于佐安, 谭克非, 张明, 等. 2016. 筏式虾夷扇贝养成期不同密度生长与经济效益分析. 水产学报, 40(10): 1624-1633 |
于佐安, 李大成, 王笑月, 等. 2019. 长海县筏养虾夷扇贝大规模死亡原因探讨. 水产科学, 38(3): 420-427 |
王波, 李有乐, 房慧, 等. 1999. 虾夷马粪海胆生物学及增殖养殖技术. 齐鲁渔业, (3): 14-16 |
元冬娟, 邵正, 程小广, 等. 2009. 冬、夏季6种经济贝类脂肪酸组成. 南方水产, 5(4): 47-53 DOI:10.3969/j.issn.1673-2227.2009.04.009 |
王兴章, 常忠岳, 吕劭伟, 等. 2004. 马粪海胆筏式养殖技术研究. 水产科技情报, 31(4): 174-175 DOI:10.3969/j.issn.1001-1994.2004.04.010 |
王崇明, 王秀华, 艾海新, 等. 2004. 栉孔扇贝大规模死亡致病病原的研究. 水产学报, 28(5): 547-553 |
史洁, 魏皓, 赵亮, 等. 2010. 桑沟湾多元养殖生态模型研究: Ⅲ海带养殖容量的数值研究. 渔业科学进展, 31(4): 43-52 DOI:10.3969/j.issn.1000-7075.2010.04.006 |
冯继兴, 许修明, 吴雪, 等. 2016. 贝类筏式养殖产业发展的主要问题及对策. 水产养殖, 37(3): 30-32 DOI:10.3969/j.issn.1004-2091.2016.03.012 |
乔方利, 袁业立, 朱明远, 等. 2000. 长江口海域赤潮生态动力学模型及赤潮控制因子研究. 海洋与湖沼, 31(1): 93-100 DOI:10.3321/j.issn:0029-814X.2000.01.015 |
刘述锡, 崔金元, 林勇. 2013. 浮筏养殖虾夷扇贝的自然稀疏效应. 水产学报, 37(10): 1513-1520 |
闫喜武, 2005. 菲律宾蛤仔养殖生物学、养殖技术与品种选育. 青岛: 中国科学院研究生院(海洋研究所)博士学位论文
|
陈炜, 曹善茂, 肖逸啸, 等. 2018. 浮筏养殖与底栖野生岩扇贝营养成分的分析与比较. 大连海洋大学学报, 33(1): 45-51 |
何毛贤, 张红玉, 袁涛. 2009. 养殖密度、水层和养殖地点对马氏珠母贝选育群体生长存活的影响. 热带海洋学报, 28(6): 68-71 DOI:10.3969/j.issn.1009-5470.2009.06.009 |
杨东敏, 王化敏, 丁鉴锋, 等. 2017. 越冬期前后菲律宾蛤仔体成分变化研究. 大连海洋大学学报, 32(6): 694-699 |
杨红生, 张福绥. 1999. 浅海筏式养殖系统贝类养殖容量研究进展. 水产学报, 23(1): 84-90 |
李华琳, 于佐安, 张明, 等. 2021. 影响筏养虾夷扇贝生长的因素分析. 河北渔业, (3): 24-26 DOI:10.3969/j.issn.1004-6755.2021.03.006 |
李锦灵, 林立龙, 李治寰. 2021. 不饱和脂肪酸对人脂肪间充质干细胞生物学特性的影响. 生命科学研究, 25(1): 15-23, 52 |
林志强, 2006. 海上筏式鲍鱼养殖标准化几个关键因素研究. 武汉: 华中农业大学硕士学位论文
|
欧洪来. 2018. 浅海牡蛎筏式养殖技术. 江西水产科技, (4): 33, 35 |
罗俊标, 骆明飞, 刘建勇. 2016. 九孔鲍新品系筏式养殖技术. 海洋与渔业, (4): 64-65 DOI:10.3969/j.issn.1672-4046.2016.04.034 |
赵春暖, 蔡忠强, 郑言鑫, 等. 2017. 渤海湾魁蚶吊笼与底播增殖实验研究. 海洋科学, 41(11): 15-24 DOI:10.11759/hykx20170712003 |
段叶辉, 李凤娜, 李丽立, 等. 2014. n-6/n-3多不饱和脂肪酸比例对机体生理功能的调节. 天然产物研究与开发, 26(4): 626-631, 479 |
郭术津, 李彦翘, 张翠霞, 等. 2014. 渤海浮游植物群落结构及与环境因子的相关性分析. 海洋通报, 33(1): 95-105 |
梁淼, 李德鹏, 孙丽艳, 等. 2018. 曹妃甸表层沉积物和底栖生物重金属的污染特征与生物积累. 海岸工程, 37(4): 17-28 DOI:10.3969/j.issn.1002-3682.2018.04.003 |
焦莉, 孙丽华, 王勇, 等. 2020. 水产动物脂类营养研究进展. 广东饲料, 29(6): 38-43 |
谢玺, 滕炜鸣, 李华琳, 等. 2020. 魁蚶生物学与增养殖技术研究进展. 大连海洋大学学报, 35(6): 930-938 |
Behrens P W, Kyle D J, 1996. Microalgae as a source of fatty acids. Journal of Food Lipids, 3(4): 259-272 DOI:10.1111/j.1745-4522.1996.tb00073.x |
Ezgeta-Balić D, Najdek M, Peharda M et al, 2012. Seasonal fatty acid profile analysis to trace origin of food sources of four commercially important bivalves. Aquaculture, 334-337: 89-100 DOI:10.1016/j.aquaculture.2011.12.041 |
Galimany E, Lunt J, Freeman C J et al, 2017. Feeding behavior of eastern oysters Crassostrea virginica and hard clams Mercenaria mercenaria in shallow estuaries. Marine Ecology Progress Series, 567: 125-137 DOI:10.3354/meps12050 |
Ghiselli F, Milani L, Chang P L et al, 2012. De Novo assembly of the Manila clam Ruditapes philippinarum transcriptome provides new insights into expression bias, mitochondrial doubly uniparental inheritance and sex determination. Molecular Biology and Evolution, 29(2): 771-786 DOI:10.1093/molbev/msr248 |
Lopez-Huertas E, 2010. Health effects of oleic acid and long chain omega-3 fatty acids (EPA and DHA) enriched milks. A review of intervention studies. Pharmacological Research, 61(3): 200-207 |
Peharda M, Ezgeta-Balić D, Davenport J et al, 2012. Differential ingestion of zooplankton by four species of bivalves (Mollusca) in the Mali Ston Bay, Croatia. Marine Biology, 159(4): 881-895 DOI:10.1007/s00227-011-1866-5 |
Zhu H R, Yu Y, Liu X B, 2020. An overview of studies on marine macrobenthic community structure and biodiversity in the Bohai Sea. Marine Science Bulletin, 22(1): 32-42 |