 2021, Vol. 52
2021, Vol. 52

三亚蜈支洲岛热带海洋牧场渔业资源现状及季节变动

http://dx.doi.org/10.11693/hyhz20210500124
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 王新元, 李建龙, 章翔, 陈孟玲, 吴沛霖, 马文刚, 彭思颖, 张玺, 谢建军, 高菲, 许强, 王爱民. 2021.
- WANG Xin-Yuan, LI Jian-Long, ZHANG Xiang, CHEN Meng-Ling, WU Pei-Lin, MA Wen-Gang, PENG Si-Ying, ZHANG Xi, XIE Jian-Jun, GAO Fei, XU Qiang, WANG Ai-Min. 2021.
- 三亚蜈支洲岛热带海洋牧场渔业资源现状及季节变动
- CURRENT SITUATION AND SEASONAL VARIATION OF FISHERY RESOURCES IN TROPICAL MARINE RANCH IN WUZHIZHOU ISLAND, SANYA, HAINAN
- 海洋与湖沼, 52(6): 1557-1566
- Oceanologia et Limnologia Sinica, 52(6): 1557-1566.
- http://dx.doi.org/10.11693/hyhz20210500124
文章历史
-
收稿日期:2021-05-28
收修改稿日期:2021-08-13
引用本文
王新元, 李建龙, 章翔, 陈孟玲, 吴沛霖, 马文刚, 彭思颖, 张玺, 谢建军, 高菲, 许强, 王爱民. 2021. 三亚蜈支洲岛热带海洋牧场渔业资源现状及季节变动[J]. 海洋与湖沼, 52(6): 1557-1566

WANG Xin-Yuan, LI Jian-Long, ZHANG Xiang, CHEN Meng-Ling, WU Pei-Lin, MA Wen-Gang, PENG Si-Ying, ZHANG Xi, XIE Jian-Jun, GAO Fei, XU Qiang, WANG Ai-Min. 2021. CURRENT SITUATION AND SEASONAL VARIATION OF FISHERY RESOURCES IN TROPICAL MARINE RANCH IN WUZHIZHOU ISLAND, SANYA, HAINAN[J]. Oceanologia et Limnologia Sinica, 52(6): 1557-1566.
三亚蜈支洲岛热带海洋牧场渔业资源现状及季节变动
海南大学海洋学院 南海海洋资源利用国家重点实验室 海口 570228
摘要:蜈支洲岛海洋牧场为海南省首个国家级海洋牧场示范区,为评估蜈支洲岛海洋牧场的资源养护效果,进而为下一步海洋牧场的渔业资源科学管理提供依据。采用渔业资源水声学调查方法,对蜈支洲海洋牧场近岛人工鱼礁区及海棠湾湾区的渔业资源现状及其季节变动进行了研究。2019年4月、8月、12月进行了3次调查,结果显示:4月近岛人工鱼礁区域共捕获游泳生物63种,平均资源量密度为63.71 t/km2,8月、12月海棠湾湾区共捕获游泳生物68种和120种,平均资源量密度分别为8.29 t/km2和7.21 t/km2。4-12月物种多样性指数(H')均值分别为3.532、3.478、4.414。三次调查中,鱼类优势种中多齿蛇鲻(Saurida tumbil)、短鳄齿鱼(Champsodon snyderi)、宽条鹦天竺鯛(Ostorhinchus fasciatus)均出现2次,其余种类均只出现一次。4-12月重要经济生物占总资源量的比重依次为72.46%、64.74%、57.59%。综上所述,蜈支洲岛海洋牧场近岛人工礁区及海棠湾湾区渔业资源丰富、物种多样性高,尤其是海洋牧场建设对于渔业资源的聚集起到了积极的作用,但仍存在重要经济鱼类少,个体较小等问题。
关键词:海洋牧场 渔业资源 渔业水声学 蜈支洲岛
CURRENT SITUATION AND SEASONAL VARIATION OF FISHERY RESOURCES IN TROPICAL MARINE RANCH IN WUZHIZHOU ISLAND, SANYA, HAINAN
The Ocean College of Hainan University, State Key Laboratory of Marine Resource Utilization in South China Sea, Haikou 570228, China
Abstract: Wuzhizhou Island Marine Ranching (WIMR) is the first national marine ranching demonstration area in South China Sea in Hainan Province, China. To evaluate the conservation effect of marine ranching resources in the island, and provide a basis for scientific management of marine ranching fishery resources in the next phase, the present situation and seasonal variation of fishery resources in artificial reef areas in nearby island of WIMR and Haitang Bay area were studied using underwater acoustic method. Three surveys were conducted in April, August and December of 2019. Results show that 63 species of nekton were captured in the artificial reef areas in April with an average resource density of 63.71 t/km2; and 68 and 120 species of nekton were captured in Haitang Bay Area in August and December, with an average resource density of 8.29 and 7.21 t/km2, respectively. The mean values of species diversity index (H') from April to December were 3.532, 3.478, and 4.414, respectively. In the three surveys, Saurida tumbil, Champsodon snyderi, and Ostrhinchus fasciatus all appeared twice, while the other species appeared only once. From the April to December, the proportion of important economic organisms in the total resources was 72.46%, 64.74%, and 57.59%, respectively. Therefore, the artificial reef areas of WIMR and Haitang Bay area were rich in fishery resources and high species diversity. In particular, the construction of marine ranching has played a positive role in the accumulation of fishery resources. However, some problems remain, such as few important economic fish and small individuals.
Key words:
marine ranching fishery resources fishery hydroacoustics Wuzhizhou Island
现代海洋牧场是集生境修复、资源养护、休闲渔业和景观生态于一体的渔业生产新业态, 真正实现了海洋经济发展和海洋生态环境保护并重(许强等, 2018)。渔业资源养护效果是评价海洋牧场建设成效的重要指标之一, 其中渔业水声学调查方法具有快速高效、对生物资源损伤小及调查海域广等优点, 这种水声学结合拖网的调查模式已逐渐成为我国渔业资源调查研究的重要手段之一(Simmonds et al, 2005; 张俊等, 2015; 李斌等, 2018), 也十分适合于海洋牧场区域的渔业资源评估。
蜈支洲岛位于海南省三亚市海棠湾内, 岛周海域拥有丰富的海洋生物资源, 由于过度捕捞, 蜈支洲岛海域渔业资源曾面临严重衰退的问题。为修复渔业资源、保护生态系统, 2012年蜈支洲岛海域开始投放人工鱼礁建设海洋牧场, 迄今为止已建设面积200 hm2, 岛周先后投放各类人工鱼礁4万余空m3, 2019年蜈支洲岛海洋牧场获批成为海南省首个国家级海洋牧场示范区。
本研究于2019年4月、8月和12月对蜈支洲海洋牧场近岛人工鱼礁区及海棠湾湾区开展了3个航次渔业资源水声学调查, 分析了牧场区渔业资源结构特征、数量密度和资源量密度的空间分布、渔业资源密度与水深的关系以及鱼类单体目标强度的空间结构分布, 并对上述指标进行了不同季节航次的平行比较, 以期评估蜈支洲岛海洋牧场的资源养护效果, 为下一步海洋牧场的渔业资源科学管理提供依据。
1 材料与方法 1.1 调查海域、时间与航线设计研究时间为2019年4月、8月和12月。研究区域位于蜈支洲海洋牧场近岛人工鱼礁投放区域(下面简称近岛礁区)及海棠湾湾区(下面简称“湾区”) (18°15′—18°23′N, 109°44′—109°50′E; 图 1), 近岛人工鱼礁投放区位于牧场范围内(图 1)。
 |
| 图 1 调查区域 Fig. 1 The survey area |
该研究主要采用等间距平行断面航线(以下简称平行航线; 图 2)进行声学调查。4月份调查范围重点覆盖近岛礁区且避开了天然珊瑚礁区, 共分为岛北、岛东和岛西3个调查区域, 分别进行近岛礁区北侧、东侧、西侧的平行航线调查(图 2a)。8月和12月航线拓展至海棠湾湾区(下面简称湾区), 以探究牧场区对更大范围内渔业资源的养护效果, 在湾区平行航线调查外, 另设计两条靠近海南岛海岸的航线, 分别命名为“湾区西南航线”和“湾区西北航线”(下面简称“西南”航线和“西北”航线; 图 2b, 2c)。
 |
| 图 2 4月(a)、8月(b)、12月(c)近岛礁区、湾区渔业资源声学调查及生物学拖网取样航线 Fig. 2 Acoustic survey and biological trawl sampling route of fishery resources in near island reef area and Bay Area in April (a), August (b), and December (c), 2019 注: 黑色示声学调查航线, 红色示拖网航线 |
声学调查的同时进行游泳动物拖网调查以辅助声学数据分析, 共布设SY1、SY2和SY3三条拖网航线(如图 2红色部分)。因4月声学调查航线覆盖区外缘距离人工鱼礁区过近, 拖网调查航线无法进入该区域, 故在保证安全的前提下尽最大可能靠近人工鱼礁区, 以为声学调查提供定性数据支持。8月和12月拖网航线纳入声学调查范围内, 且与4月份航线保持一致。
1.2 数据采集 1.2.1 渔业生物群落拖网调查游泳动物拖网调查船为单拖网渔船(琼临渔00136, 船长27.0 m, 船宽5.6 m, 主机450 kW)。网具尺寸为24 m长×16.8 m宽×0.58 m高, 网目为4.5 cm, 网囊网目为2.5 cm。拖网调查中, 每个航线拖拽时间为50—62 min, 拖速为1.7—2.0 n mile/h。
每站渔获物全部保留, –20 ℃冷藏带回实验室进行鉴定和生物学指标测定。记录每网产量, 个体数量≤50尾(只)的物种均进行生物学测定, 个体数量 > 50尾(只)的物种随机取50尾(只)进行测定。鱼类的测定指标为体长和体重; 头足类的测定指标为胴长和体重, 结合以上数据, 并根据拉丁文学名在FishBase网站查阅物种相关信息, 在Length at first maturity / Size / Weight / Age一栏找到所需物种的成熟期对应体长, 进而进行生物幼体与成体信息统计。渔获物中的虾类和蟹类未分析。
1.2.2 渔业水声学数据收集采用美国BIOSONICS DT-X分裂波束科研鱼探仪进行声学数据采集, 工作频率200 kHz。声学调查船为蜈支洲岛旅游公司玻璃钢制快艇“海鹭2号”, 艇长12.5 m, 艇宽2.5 m, 主机320 kW。将换能器置于导流罩内并置于水下1 m, 通过螺杆及固定架固定于船只左舷中部且不与船体接触(张俊等, 2015)。按照国际通用的标准球法对鱼探仪系数进行现场校正(SIMRAD, 2008)。声学扫测时航速控制在4节左右。声学数据采用设备标配的Visual Analyser软件进行分析, 剔除声学数据中换能器表面以下1 m以内及海底以上0.5 m以内的数据, 基本积分航程单元设为0.5 n mile, 积分阈值设为–70 dB以屏蔽浮游生物等弱散射体的回波信号(赵宪勇等, 2002; Higginbottom et al, 2008; 张俊, 2011)。由于剔除了距海底0.5 m以内的回声数据, 因此各航次中底层生物如鲆鲽类、鰕虎鱼类、蛸类、虾类、蟹类等不参与声学评估。
1.3 统计分析 1.3.1 渔业资源结构评价指标及方法(1) 相对重要性指数IRI及计算 用相对重要性指数IRI来研究游泳动物优势种的优势度, 公式为:
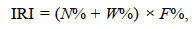 (1)
(1)式中, N%为某一物种尾数占总尾数的百分比; W%为该物种重量占总重量的百分比; F%为为某一物种出现的站数占调查总站数的百分比。
(2) 生态评价指数及计算(江艳娥等, 2009)
香农威纳(Shannon-Wiener)物种多样性指数(CH′), 公式为:
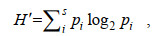 (2)
(2)式中, H′为物种多样性指数值; S为样品中的总种数; Pi为第i种的个体丰度(ni)与总丰度(N)的比值(ni/N)。
Pielou均匀度指数, 公式为:
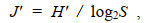 (3)
(3)式中, J′表示均匀度指数值; H′表示物种多样性指数值; S表示样品中总种数。
Margalef丰富度指数, 公式为:
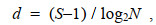 (4)
(4)式中, d表示丰富度指数值; S表示样品中的总种数; N表示群落中所有物种的总丰度。
单纯度指数, 公式为:
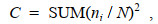 (5)
(5)式中, C表示单纯度指数; N表示群落中所有物种丰度或生物量; ni表示第i个物种的丰度或生物量; SUM表示求和。
1.3.2 渔业声学数据处理方法按照多种类海洋渔业资源声学评估技术与方法(赵宪勇等, 2002; 李永振等, 2003)的工作流程, 对鱼类资源的数量密度和资源量密度进行评估, 以拖网调查的渔获物组成信息作为积分值分配的主要依据(李斌等, 2018)。调查区域内第i种鱼类的数量密度ρi-a(个/km2)和资源量密度ρi-b(t/km2)分别为:
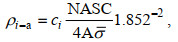 (6)
(6)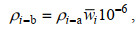 (7)
(7)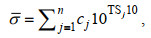 (8)
(8)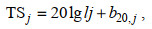 (9)
(9)式中, ci(%)是分析水域内第i种鱼类占渔获物的数量百分比; NASC(m2/n mile2)是分析水域内参与积分值分配生物种类的总积分值(nautical area scattering coefficient); 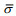
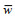
表 1 海洋牧场声学评估种类b20值 Tab. 1 Acoustic estimation species of marine ranch and their b20 values
| 种类 | 拉丁文学名 | b20(dB) | 种类 | 拉丁文学名 | b20(dB) | |
| 狗母鱼科 | Synodontidae | –72.5 | 黄斑鲾 | Leiognathus bindus | –80.0 | |
| 鲉科 | Scorpaenidae | –72.5 | 发光鲷 | Acropoma japonicum | –68.0 | |
| 石首鱼科 | Sciaenidae | –68.0 | 短尾大眼鲷 | Priacanthus macracanthus | –68.0 | |
| 鲹科 | Carangidae | –72.5 | 日本红娘鱼 | Lepidotrigla japonica | –68.0 | |
| 金线鱼科 | Nemipteridae | –68.0 | 竹荚鱼 | Trachurus japonicus | –72.5 | |
| 鲭科 | Scombrida | –76.0 | 鳗鲡类 | Anguilliformes spp. | –76.0 | |
| 石鲈科 | Pomadasyidae | –68.0 | 蛇鲻类 | Saurida sp. | –72.5 | |
| 鲀科 | Tetraodontidae | –76.0 | 小公鱼类 | Stolephorus sp. | –68.0 | |
| 天竺鲷科 | Apogonidae | –68.0 | 其他鲾类 | Other Leiognathus | –72.5 | |
| 无磷烟管鱼 | Fistularia commersonii | –76.0 | 棱鳀类 | Thrissa sp. | –72.5 | |
| 鯻 | Terapon theraps | –72.5 | 犀鳕类 | Bregmaceros sp. | –72.5 | |
| 细鳞鯻 | Terapon jarbua | –72.5 | 石斑鱼类 | Epinephelus sp. | –72.5 | |
| 南海带鱼 | Trichiurus nanhaiensis | –66.1 | 绯鲤类 | Parus spp. | –72.5 | |
| 蓝圆鲹 | Decapterus maruadsi | –72.5 |  |
Callioymidae spp. | –72.5 | |
| 六指马鲅 | Polydactylus sexfilis | –80.0 | 鲈类 | Percidae spp. | –72.5 | |
| 多鳞鱚 | Sillago sihama | –72.5 | 鳄齿鱼类 | Champsodon sp. | –68.0 | |
| 银鲳 | Pampus argenteus | –80.0 | 枪乌贼类 | Loliginidae | –78.0 | |
| 刺鲳 | Psenopsis anomala | –80.0 | 其他乌贼类 | other Sepiidae | –76.0 |
海里面积散射系数NASC是衡量资源量的重要指数之一。为了解渔业资源垂直分布特征, 本研究以10 m为区间对各航线声学数据进行垂直划分, 统计各水层NASC值及百分比。
2 结果 2.1 渔业生物多样性与优势种分析4月共捕获鱼类59种, 分别属于9目、35科, 捕获头足类4种; 8月捕获鱼类64种, 除两个未知种外, 其余属于11目、40科, 捕获头足类4种。12月捕获鱼类115种, 分别属于11个目、49个科, 捕获头足类5种。三次调查, 12月渔获物种类数最多, 4月最少。
根据IRI结果分析(见表 2), 三次调查中, 鱼类优势种多齿蛇鲻(Saurida tumbil)、短鳄齿鱼(Champsodon snyderi)、宽条鹦天竺鲷(Ostorhinchus fasciatus)均出现2次, 布希勒副单角鲀(Paramonacanthus pusillus)、短吻鲾(Leiognathus brevirostris)、细纹鲾(Leiognathus berbis)、福氏钝塘鳢(Amblyeleotris fontanesii)、小牙鲾(Gazza mimuta)、长棘鲾(Leiognathus fasciatus)、布氏长鳍天竺鲷(Archamia bleekeri)、丝鳍美尾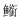
表 2 各次调查渔获物优势度前5位鱼类生物学组成信息 Tab. 2 Biological composition of the top 5 fish in catch dominance in each survey
| 调查月份 | 种类 | 优势度 | 仔稚鱼比例 | 体长(cm) | 体重(g) | |||
| 范围 | 平均值 | 范围 | 平均值 | |||||
| 4月 | 多齿蛇鲻 | 3557.16 | 1.12% | 8.00—20.00 | 14.00 | 4.09—107.16 | 31.15 | |
| 短鳄齿鱼 | 3087.40 | 100.00% | 3.40—7.60 | 5.10 | 1.27—1.74 | 1.60 | ||
| 布希勒副单角鲀 | 2707.53 | 87.50% | 6.00—10.60 | 7.40 | 7.15—41.85 | 12.67 | ||
| 宽条鹦天竺鲷 | 2570.52 | 98.81% | 2.40—9.30 | 12.60 | 0.79—25.20 | 1.18 | ||
| 短吻鲾 | 581.13 | 98.09% | 2.20—6.50 | 2.70 | 0.65—7.50 | 0.61 | ||
| 8月 | 多齿蛇鲻 | 6325.35 | 50.00% | 3.00—57.70 | 11.70 | 0.15—197.58 | 28.30 | |
| 短鳄齿鱼 | 2387.76 | 100.00% | 0.32—6.10 | 4.30 | 0.25—5.98 | 0.99 | ||
| 宽条鹦天竺鲷 | 1122.04 | 94.29% | 1.90—8.00 | 3.60 | 0.05—16.72 | 1.74 | ||
| 细纹鲾 | 977.96 | 42.31% | 3.90—11.40 | 7.60 | 1.65—54.68 | 18.79 | ||
| 福氏钝塘鳢 | 853.23 | 93.00% | 2.20—8.70 | 4.10 | 0.10—7.72 | 0.98 | ||
| 12月 | 小牙鲾 | 5626.61 | 47.60% | 4.10—11.60 | 7.00 | 1.43—42.65 | 9.25 | |
| 长棘鲾 | 3528.34 | 47.62% | 3.50—20.00 | 9.40 | 1.02—252.00 | 43.20 | ||
| 布氏长鳍天竺鲷 | 2663.59 | 99.50% | 2.90—9.50 | 5.70 | 0.53—14.16 | 5.16 | ||
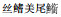 |
1016.75 | 98.26% | 4.10—8.50 | 7.50 | 0.80—6.77 | 3.40 | ||
| 长体银鲈 | 971.11 | 0.00% | 12.20—15.00 | 13.40 | 49.67—92.90 | 62.99 | ||
三次底拖网调查渔获物生态评价指数值见表 3。在重量评价中, 12月的多样性指数(H’)、均匀度指数(J’)、丰富度指数(d)均为三个月份最大值, 单纯度指数(C)为三个月份最小值; 4月和8月各项数据大小互有交替。说明12月份群落结构更加稳定, 生态环境更健康。
表 3 各次底拖网调查渔获物生态评价指数值 Tab. 3 Ecological evaluation index value of catch in each bottom trawl survey
| 调查月份 | 重量评价 | |||
| 物种多样性指数(H′) | 均匀度指数(J′) | 丰富度指数(d) | 单纯度指数(C) | |
| 4月 | 3.532 | 0.681 | 2.857 | 0.150 |
| 8月 | 3.478 | 0.657 | 3.304 | 0.185 |
| 12月 | 4.414 | 0.716 | 5.354 | 0.108 |
研究分析了各航次单体生物目标强度TS值的频度分布(图 3), 发现该海域内单体生物目标强度分布差异不大, 主要分布于–65—–25 dB, 其中以–65—–60 dB占据主要部分, 指示小个体渔业生物, 4—12月该部分回声信号分别约占80.97%、74.77%和85.73%。
 |
| 图 3 4月、8月、12月目标强度频度分布图 Fig. 3 Target intensity frequency distribution in April, August, and December, 2019 |
为考察渔业资源的垂直空间分布特征, 对渔业资源在垂直水深方向上的空间分布进行统计分析, 以10 m的深度为间隔, 分别分析各水层内的声学积分值及其所占百分比(图 4)。4—12月游泳生物存在趋向浅水层移动的现象, 这可能与仔稚鱼的昼夜垂直分布与底质环境相关, 根据所有渔获物仔稚鱼数量的比例分析, 4—12月份渔获中仔稚鱼数量比分别为78.53%、82.63%、67.34%。
 |
| 图 4 各月积分值垂直分布 Fig. 4 Vertical distribution of the nautical area scattering coefficient (NASC) |
根据水声学数据分析三次调查的平均数量密度和平均资源量密度, 见表 4。
表 4 2019年4—12月平均资源量密度与平均数量密度 Tab. 4 Average resource density and average quantity density from April to December in 2019
| 调查月份 | 区域 | 平均资源量密度(t/km2) | 平均数量密度(尾/km2) |
| 4月 | 近岛礁区北侧 | 12.24 | 1.20×106 |
| 近岛礁区西侧 | 19.55 | 1.92×106 | |
| 近岛礁区东侧 | 150.50 | 1.48×107 | |
| 8月 | 湾区西北航线 | 11.00 | 9.89×105 |
| 湾区西南航线 | 110.47 | 9.92×106 | |
| 湾区平行航线 | 8.29 | 7.42×105 | |
| 12月 | 湾区西北航线 | 4.02 | 3.03×105 |
| 湾区西南航线 | 177.99 | 7.47×107 | |
| 湾区平行航线 | 7.21 | 5.45×105 |
4月份近岛礁区东侧的两项密度数据均为最高, 其次为西侧。将全部航线资源量密度经lg转化后作图(图 5)分析, 4月份渔业资源主要分布在岛东及岛东南, 尤其岛东南资源量最高。
 |
| 图 5 4月(a)、8月(b)、12月(c)资源量密度 Fig. 5 Resource density in April (a), August (b), and December (c), 2019 |
8月份湾区“西北”、“西南”航线两项密度数据均高于平行航线, 其中“西南”航线平均数量密度和平均资源量密度分别为平行航线的13.38倍和13.81倍; 12月份湾区“西南”航线两项密度数据最高, 均为平行航线的24.69倍, “西北”航线两项密度数据则低于平行航线。根据图 5分析, 8月份渔业资源在湾区西南方向最高, 其次为湾区东侧; 12月份渔业资源在湾区西南方向最高, 其次为湾区东南方向。
根据实地调查结果表明, 蜈支洲岛东及东南广布天然礁石, 蜈支洲岛西南方向界于蜈支洲岛西南侧和海南岛近岸区域之间, 是一个天然的海流通道, 东侧蜈支洲岛有丰富的人工鱼礁, 西侧近岸区域有海草床分布, 以上均对渔业资源空间分布产生较大影响。
2.4 重要渔获物资源量基于三个航次的所有航线调查数据, 对优势游泳动物进行了声学评估与分析, 由于福氏钝塘鳢属鰕虎鱼科, 不参与声学评估, 因此8月优势度第5位鱼种改为条纹银口天竺鲷(Jaydia striata): 4—12月, 5种优势鱼类平均资源量密度之和分别为20.87、11.17和13.37 t/km2, 占总资源量密度的比例分别为32.75%、52.17%和45.89%。
4—12月优势头足类均为杜氏枪鱿和曲针乌贼, 三航次两种优势头足类平均资源量密度之和分别为25.3、2.69和3.41 t/km2, 占总资源量密度的比例分别为39.71%、12.57%和11.70%。
从调查结果来看, 4—12月, 优势鱼类和优势头足类资源量密度之和占总资源量密度的比值呈下降趋势, 优势头足类占总资源量密度的比值亦呈下降趋势, 而优势鱼类占总资源量密度的比值8月最高, 4月最低(图 6)。
 |
| 图 6 优势游泳动物平均资源量密度的季节变化 Fig. 6 Seasonal variation in the average resource density of dominant species of swimming animals |
在此次声学评估中共分析头足类的杜氏枪鱿及曲针乌贼两种(图 7)。
 |
| 图 7 头足类平均资源量密度的季节变化 Fig. 7 Seasonal variation in the average resource density of cephalopods |
根据杜氏枪鱿和曲针乌贼的平均资源量密度比较分析, 4—12月杜氏枪鱿平均资源量密度呈降低趋势, 4月杜氏枪鱿平均资源量密度为曲针乌贼资源量的4.46倍, 8月为1.82倍, 而12月曲针乌贼平均资源量密度超越杜氏枪鱿, 为杜氏枪鱿的3.16倍。根据杜氏枪鱿和曲针乌贼的资源量占比比较分析同样可得, 杜氏枪鱿资源量占比随季节变化明显下降, 而曲针乌贼资源量占比变化相对较小。
2.4.2 鲹科鱼类在此次声学评估中共分析鲹科鱼类的蓝圆鲹(Decapterus maruadsi)及日本竹荚鱼(Trachurus japonicus)两种。4—12月蓝圆鲹平均资源量密度分别为0.38、0.20和0.18 t/km2 (图 8)。由此可见, 蓝圆鲹平均资源量密度随季节变化呈下降趋势。4—8月日本竹荚鱼平均资源量密度分别为0.61、0.24 t/km2, 12月未捕获到日本竹荚鱼(图 8), 其原因可能与季节性洄游性有关。
 |
| 图 8 重要经济鱼类平均资源量密度的季节变化 Fig. 8 Seasonal variations in the average resource density of important economic fish |
4—12月南海带鱼平均资源量密度分别为0.46、0.48和0.51 t/km2 (图 8)。由此可见, 4—12月南海带鱼的平均资源量密度呈上升趋势。
2.4.4 金线鱼科鱼类在本研究3个航次的声学评估中共分析的3个种类有日本金线鱼(Nemipterus japonicus)、黄缘金线鱼(N. thosaporni)、深水金线鱼(N. bathybius)。4—12月金线鱼类平均资源量密度分别为0.11、0.36和0.76 t/km2 (图 8)。由此可见, 4—12月金线鱼科鱼类的平均资源量密度呈上升趋势。
3 讨论 3.1 渔业资源量现状评价在本文的三次调查研究中, 4—12月渔获物物种多样性指数H'高于福建东山湾海域(张静等, 2013)、福建三沙湾海域(林龙山等, 2005)、台湾海峡的全年平均多样性水平(沈长春, 2011), 低于南海北部大陆架海域的全年平均多样性水平(江艳娥等, 2009)。多样性指数低于南海北部大陆架海域, 可能是该调查结果去除了虾蟹类相关数据导致的, 但仍然高于福建、台湾等海域, 可以说海棠湾生物多样性、群落结构稳定性以及生态环境相对较好。
2014年11月、2015年8月及2016年1月在海南岛陵水湾进行了3个航次的渔业资源声学调查, 渔业资源的平均资源量密度依次为5.08、0.93和0.32 t/km2 (李斌等, 2018)。本研究均高于以上数据, 可能由于自2011年蜈支洲岛海洋牧场启动建设以来, 通过合理投放人工鱼礁以及海洋牧场区严格的保护与管理措施, 对岛周毗邻海域的渔业资源恢复起到了积极作用。
在重要经济生物资源量占比的比较中, 头足类所占比重最大, 但其所占比重随季节持续下降, 其直接原因为杜氏枪鱿的平均资源量密度占比随季节变化明显下降, 这可能由于杜氏枪鱿的繁殖行为息息相关。1997—1999年在南海北部海域的调查中, 南海带鱼平均资源量密度约为1.34 t/km2, 蓝圆鲹为0.43 t/km2, 竹荚鱼为0.28 t/km2 (陈国宝等, 2005), 本研究与之相比, 除南海带鱼类平均资源量密度相差较大外, 蓝圆鲹和竹荚鱼平均资源量密度相似性颇高。整体来看, 本研究中重要经济鱼类的总资源量普遍偏小, 这可能与对经济鱼类的过度捕捞有关。
3.2 渔业资源时空变化从垂直空间分布来看, 随季节变化, 游泳生物存在趋向浅水层移动的现象。相关研究表明, 部分仔稚鱼具有在昼伏夜出习性(林楠, 2019), 白天在深水层躲避捕食者, 夜间上浮摄食。由于4月水域为人工鱼礁区, 营造了良好的底层栖息地, 为渔业资源提供了适宜的避敌与索饵场所, 因此4月份渔业资源回声信号分布于底层区域; 8月仔稚鱼比例高于12月份, 且本研究航次调查均在白天进行, 因此8月渔业资源分布水层较12月份深。
根据礁区水平空间分布结果显示, 礁区东及东南方向渔业资源密集, 据前期海底地形地貌勘测分析, 蜈支洲岛岛东及东南区域水深较深, 海底天然礁石分布面积广, 同时建有人工鱼礁区, 适合于鱼类作为栖息场所, 因此形成了岛东及东南资源量密集的现象。
根据湾区水平空间分布结果显示, 湾区西南方向渔业资源密集, 该区域界于蜈支洲岛西南侧和海南岛近岸区域之间, 是一个天然的海流通道, 东侧蜈支洲岛有丰富的人工鱼礁, 西侧近岸区域有海草床分布, 因此该区域渔业资源密集。
根据湾区总体调查结果显示, 夏季资源量高于冬季, 可能由于8月进行资源调查时刚刚经历了几个月的休渔期, 渔业资源密度较大; 12月近岸海表温度低, 受部分鱼类游向深水区越冬影响(王迪等, 2006), 造成近岸渔业资源密度降低。但12月份渔获物种类最为丰富, 这可能与部分鱼类的迁移性相关, 并且温度、海水溶解氧度等海洋环境因子及其变化也同样是影响鱼类分布、季节性洄游、迁徙的重要因素(胡奎伟等, 2012; 李斌等, 2016)。
3.3 水声学调查与其他调查方法的分析热带海洋牧场由于珊瑚礁及人工鱼礁的存在, 给渔业资源调查带来重重考验。传统资源调查方式分非捕捞调查方法包括光学法(水下摄像和记录)和直接捕捞法(拖网和钓具) (汪振华等, 2010; 陈丕茂等, 2013; 陈勇等, 2014)。光学法受限于水下光的传播距离以及专业的潜水调查人员; 直接捕捞方法效率较低, 易受生境等条件限制, 如传统拖网调查难以在海洋牧场内部核心区应用。渔业资源声学评估方法能沿着调查航线对表层盲区和底层死区外的全水层鱼类分布及其资源量进行三维定量研究, 全面反映鱼类的时空分布及变动, 具有高效、不破坏生物资源、时空数据丰富等优点, 是目前海洋渔业资源调查主流方法之一, 已成为海洋牧场渔业资源评估新的有效方法(Simmonds et al, 2005; Hamano, 2010; 李娜娜等, 2011; 张俊等, 2014; Huang et al, 2014)。声学法与直接捕捞法或光学法的有机结合可能是评估海洋牧场渔业资源最好的选择之一(张俊等, 2015)。
4 结论根据资源量的时空分布得出, 本研究调查区域渔业资源量较为丰富, 生物多样性、群落结构稳定性以及生态环境相对较好, 且渔业资源密集区主要集中于人工鱼礁投放区、海草床毗邻区, 因此蜈支洲岛海洋牧场的建设对渔业资源的恢复以及生态资源的修复具有积极意义; 根据仔稚鱼的比例判断, 休渔期制度的执行也在一定程度上为渔业资源的有效繁衍生息提供重要保障; 但整体来看, 该海域重要经济鱼类在总资源量中所占比重普遍偏小, 这可能与经济鱼类的过度捕捞有关, 因此合理控制捕捞量, 充分发挥海洋牧场在资源养护中的作用, 维持资源的可持续利用显得尤为重要。综上所述, 依托岛礁的海洋牧场建设模式是南海热带海域开展渔业资源养护与修复的有效手段, 蜈支洲岛海洋牧场应继续加大对于不同功能的组合型人工鱼礁投放力度, 为海洋生物营造更加适宜的栖息环境, 找到资源养护与旅游开发的契合点。政府应继续严格执行休渔期制度, 合理控制捕捞量及渔获物个体大小, 为资源恢复提供重要保障。
参考文献
王迪, 林昭进. 2006. 珠江口鱼类群落结构的时空变化. 南方水产, 2(4): 37-45 |
江艳娥, 林昭进, 黄梓荣. 2009. 南海北部大陆架区渔业生物多样性研究. 南方水产, 5(5): 32-37 |
许强, 刘维, 高菲, 等. 2018. 发展中国南海热带岛礁海洋牧场——机遇、现状与展望. 渔业科学进展, 39(5): 173-180 |
李斌, 陈国宝, 于杰, 等. 2018. 海南陵水湾口海域不同季节鱼类资源声学探查. 水产学报, 42(4): 544-556 |
李斌, 陈国宝, 郭禹, 等. 2016. 南海中部海域渔业资源时空分布和资源量的水声学评估. 南方水产科学, 12(4): 28-37 DOI:10.3969/j.issn.2095-0780.2016.04.004 |
李永振, 陈国宝, 孙典荣, 等. 2003. 南海区多鱼种声学评估工作程序. 湛江海洋大学学报, 23(1): 35-40 |
李永振, 陈国宝, 赵宪勇, 等. 2005. 南海北部海域小型非经济鱼类资源声学评估. 中国海洋大学学报, 35(2): 206-212 |
李娜娜, 陈国宝, 于杰, 等. 2011. 大亚湾杨梅坑人工鱼礁水域生物资源量声学评估. 水产学报, 35(11): 1640-1649 |
汪振华, 章守宇, 王凯, 等. 2010. 三横山人工鱼礁区鱼类和大型无脊椎动物诱集效果初探. 水产学报, 34(5): 751-759 |
沈长春. 2011. 福建三沙湾鱼类群落组成特征及其多样性. 海洋渔业, 33(3): 258-264 |
张俊, 2011. 基于声学数据后处理系统的黄海鯷鱼资源声学评估. 上海: 上海海洋大学硕士学位论文
|
张俊, 陈丕茂, 房立晨, 等. 2015. 南海柘林湾-南澳岛海洋牧场渔业资源本底声学评估. 水产学报, 39(8): 1187-1198 |
张俊, 陈作志, 陈国宝, 等. 2014. 南海鸢乌贼水声学测量和评估相关技术研究. 南方水产科学, 10(6): 1-11 |
张静, 陈永俊, 宋普庆, 等. 2013. 福建东山湾游泳动物群落物种组成及其多样性. 海洋渔业, 35(1): 15-23 |
陈勇, 杨军, 田涛, 等. 2014. 獐子岛海洋牧场人工鱼礁区鱼类资源养护效果的初步研究. 大连海洋大学学报, 29(2): 183-187 |
陈丕茂, 袁华荣, 贾晓平, 等. 2013. 大亚湾杨梅坑人工鱼礁区渔业资源变动初步研究. 南方水产科学, 9(5): 100-108 |
陈国宝, 李永振, 赵宪勇, 等. 2005. 南海北部海域重要经济鱼类资源声学评估. 中国水产科学, 12(4): 445-451 |
陈国宝, 李娜娜, 陈丕茂, 等. 2010. 南海黄斑蓝子鱼的目标强度测量研究. 中国水产科学, 17(6): 1293-1299 |
林楠. 2019. 长江口水域仔稚鱼垂直分布研究. 浙江海洋大学学报(自然科学版), 38(6): 502-508 |
林龙山, 郑元甲, 马春艳. 2005. 台湾海峡夏秋季游泳动物资源分布及群落结构. 应用生态学报, 16(10): 1948-1951 |
赵宪勇, 陈毓桢, 李显森等, 2002. 多种类海洋渔业资源声学评估技术与方法. 见: 我国专属经济区和大陆架勘测研究专项学术交流会论文集. 北京: 海洋出版社
|
胡奎伟, 许柳雄, 陈新军, 等. 2012. 海洋遥感在渔场分析中的研究进展. 中国水产科学, 19(6): 1078-1087 |
Chen G B, Li Y Z, Chen P M et al, 2012. Measurement of single-fish target strength in the South China Sea. Chinese Journal of Oceanology and Limnology, 30(4): 554-562 |
Hamano A, 2010. Assessment of the effect of artificial reef on fish distribution: the combined use of acoustic data and GIS. In: Proceedings of the 13th French-Japanese Oceanography Symposium. Marseille, France: Springer, 99-103
|
Higginbottom I, Woons S, Schneider P, 2008. Hydroacoustic Data Processing for Standard Stock Assessment Using Echoview: Technical Manual. Australia: Myriax Software Pty Ltd Publication, 1-139
|
Huang R N, Han J, Tong J F, 2014. Assessment of fishery resource of a marine ranching based on a DIDSON. In: OCEANS 2014-TAIPEI. Taipei, China: IEEE, 1-5
|
Simmonds E J, MacLennan D N, 2005. Fisheries Acoustics: Theory and Practice. 2nd ed. Oxford: Blackwell Science
|
SIMRAD, 2008. Simrad ER60 scientific echo sounder software reference manual. Norway: Simrad Maritime AS Kongsberg
|
Zhang J, Chen P M, Chen G B et al, 2013. Acoustic target strength measurement of banded grouper[Epinephelus awoara (Temming & Schlegel..1842)] and threadsial filefish[Stephanolepis cirrhifer (Temming & Schlegel..1850)] in the South China Sea. Journal of Applied Ichthyology, 29(6): 1453-1455 |