 2022, Vol. 53
2022, Vol. 53
仿生态条件下星洲红鱼(Singapore red tilapia)和奥尼罗非鱼(Oreochromis niloticus)生长和肌肉营养特性的比较研究

http://dx.doi.org/10.11693/hyhz20210700169
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 林祥日. 2022.
- LIN Xiang-Ri. 2022.
- 仿生态条件下星洲红鱼(Singapore red tilapia)和奥尼罗非鱼(Oreochromis niloticus)生长和肌肉营养特性的比较研究
- COMPARISON IN GROWTH AND MUSCLE NUTRITION OF SINGAPORE RED TILAPIA AND OREOCHROMIS NILOTICUS UNDER SIMULATED ECOLOGICAL CONDITIONS
- 海洋与湖沼, 53(1): 141-149
- Oceanologia et Limnologia Sinica, 53(1): 141-149.
- http://dx.doi.org/10.11693/hyhz20210700169
文章历史
-
收稿日期:2021-07-26
收修改稿日期:2021-08-31
引用本文
林祥日. 2022. 仿生态条件下星洲红鱼(Singapore red tilapia)和奥尼罗非鱼(Oreochromis niloticus)生长和肌肉营养特性的比较研究[J]. 海洋与湖沼, 53(1): 141-149

LIN Xiang-Ri. 2022. COMPARISON IN GROWTH AND MUSCLE NUTRITION OF SINGAPORE RED TILAPIA AND OREOCHROMIS NILOTICUS UNDER SIMULATED ECOLOGICAL CONDITIONS[J]. Oceanologia et Limnologia Sinica, 53(1): 141-149.
仿生态条件下星洲红鱼(Singapore red tilapia)和奥尼罗非鱼(Oreochromis niloticus)生长和肌肉营养特性的比较研究
厦门海洋职业技术学院海洋生物学院 福建厦门 361100
摘要:为比较星洲红鱼(Singapore red tilapia)和奥尼罗非鱼(Oreochromis niloticus)的生长和肌肉营养特性,实验利用两种鱼的不同体色将其混养在同一池塘中,在相同的环境条件和养殖管理下研究其生长特性,并分析肌肉营养成分。结果表明:两种罗非鱼生长在前60 d无差异,60 d后星洲红鱼的体长、体质量日增长和特定生长率均大于奥尼罗非鱼,且差异显著(P < 0.05)。据此得到了星洲红鱼和奥尼罗非鱼的体长(L)、体质量(W)生长与养殖天数(d)的回归方程。星洲红鱼含肉率高于奥尼罗非鱼9.65%(P < 0.05),肌肉水分高于奥尼罗非鱼,蛋白质、粗脂肪和灰分含量均低于奥尼罗非鱼,其中脂肪含量差异显著(P < 0.05),其他不显著。两种罗非鱼肌肉的氨基酸组成全面,氨基酸组成均符合FAO/WHO的理想模式,除精氨酸含量有差异外,其他氨基酸含量稍有差异但不显著。星洲红鱼和奥尼罗非鱼肌肉EAA/TAA、EAA/NEAA、DAA/TAA和EAAI分别为41.04%和40.95%,70.00%和69.34%,40.71%和39.08%,67.46%和67.08%,差异不显著。星洲红鱼、奥尼罗非鱼肌肉∑SFA和∑UFA分别为28.03%和28.60%,63.87%和61.38%,各脂肪酸含量除C14:0和C18:2外,其他含量差异不显著。研究表明,星洲红鱼和奥尼罗非鱼生长快、营养丰富、食用价值高,是优质淡水鱼类养殖品种,其中星洲红鱼含肉率高,生长快,养殖效益明显。
关键词:星洲红鱼(Singapore red tilapia) 奥尼罗非鱼(Oreochromis niloticus) 生长 肌肉营养评价
COMPARISON IN GROWTH AND MUSCLE NUTRITION OF SINGAPORE RED TILAPIA AND OREOCHROMIS NILOTICUS UNDER SIMULATED ECOLOGICAL CONDITIONS
College of Marine Biology, Xiamen Ocean Vocation College, Xiamen 361100, China
Abstract: To compare in growth and muscle nutrition of Singapore red tilapia (ST) and Oreochromis niloticus (O. niloticus♀×O. aureus♂) (ON), two species of tilapia in different body colors were polycultured in the same pond under the same environmental conditions and culture management. Biochemical analysis was performed to compare the meat content and muscle nutrient composition. The results show that there was no difference before the first 60 days in the growth of body length and body weight of the two tilapias. After 60 days, the growth rate of body length, body weight, and specific growth rate (P < 0.05) of ST were higher than ON. The regression equations of body length (L), body weight (W) growth, and culture days (d) of ST and ON were established. The meat content had 9.65% more in ST than in ON (P < 0.05). The protein, crude fat, and ash content in muscles of ST were lower than those of ON except for water content. The amino acid composition in muscles of the two tilapias was comprehensive, and conformed to ideal mode of FAO/WHO. Except for arginine content, there was no significant difference among the other amino acid contents. EAA/TAA, EAA/NEAA, DAA/TAA and EAAI of ST and ON were 41.04% and 40.95%, 70.00% and 69.34%, 40.71% and 39.08%, 67.46% and 67.08%, respectively, with no significant difference. The contents of ∑SFA and ∑UFA in the muscle of ST and ON were 28.03% and 28.60%, 63.87% and 61.38%, respectively. Except for C14:0 and C18:2, there was no significant difference among other fatty acids. This study indicated that both ST and ON are fast-growing quality freshwater fishes, being featured with strong stress resistance, rich nutrition, and high edible value. ST had higher meat content, faster growth, and shall have greater breeding benefits than those of ON.
Key words:
Singapore red tilapia Oreochromis niloticus (O. niloticus♀×O. aureus♂) growth evaluation of muscle nutrition
罗非鱼隶属鲈形目(Perciformes)、丽鲷科(Cichlidae)、罗非鱼属(Tilapia), 是仅次于鲤科(Carps)和鲑科(Salmonids)的世界上第三大养殖品种。罗非鱼生长快、抗逆性强、肉质鲜美、营养价值高, 素有“白肉三文鱼”之称。1976年联合国粮农组织(FAO)向世界推广养殖罗非鱼, 2019年全球罗非鱼养殖产量约为650万t, 我国为164.17万t, 占全球罗非鱼产量的25.23%, 成为我国主要的养殖鱼类之一(农业农村部渔业渔政管理局等, 2020)。
星洲红鱼(Singapore red tilapia)是从(Oreochromis niloticus×O. mossambicus)杂交后代中选择红色体型的个体, 并进一步选育而获得的杂交品种, 其体色红艳、生长快、个大肉厚、适应强, 养殖优势明显, 可集观赏、养殖、垂钓于一体, 具有发展潜力的淡水养殖新品种(黄永春等, 2012)。奥尼罗非鱼(Oreochromis niloticus)是奥利亚罗非鱼(O. aureus×O. niloticus) 杂交子一代, 其具有雄性率高、生长快、抗病力强及肉质细腻等杂交优势明显的养殖品种(夏德全, 2003), 两种罗非鱼产业优势明显。
目前, 有关奥尼罗非鱼的健康养殖(杨弘等, 2005)、生长(林仕梅等, 2007; 王辉等, 2008; 强俊等, 2009; Florio et al, 2009; 肖炜等, 2012; 彭淇等, 2014; 张宇航等, 2020), 遗传改良(Ponzoni et al, 2011; Ma et al, 2012; Trọng et al, 2013a, b)和肌肉营养(乐贻荣等, 2015; 王煜坤等, 2018; 肖炜等, 2020)方面已有研究, 星洲红鱼的研究主要在形态(黄永春等, 2011)、养殖(许洪玉, 2007)和生长(许洪玉, 2007; 康自强, 2014; 黄永春, 2015)的报道, 肌肉营养鲜有报道, 两种鱼的生长比较仅见许洪玉(2007)星洲红鱼和奥尼罗非鱼养殖对比试验, 这些实验仅对某一种鱼类所设计的。由于两种罗非鱼的体色不同, 本实验选择室外常规3口的相邻池塘, 将两种罗非鱼混养在一起。根据罗非鱼的摄食特性、生活空间, 配养滤食性的鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)及杂食性的鲤(Cyprinus carpio)等, 利用其养殖生态位互补的特点循环利用营养物质, 实施养殖系统的生物调控与自我修复, 维持和改善池塘生态系统的健康状况, 从而改善水质、生态防病、提升质量安全、提高养殖效益(陈家长等, 2010; 钟全福, 2019)。同时在相同的养殖管理和环境条件下研究两者的生长特性, 分析评价肌肉营养成分, 为这两种鱼类的健康养殖、饵料研制和加工利用提高科学依据, 促进罗非鱼绿色健康养殖的可持续发展。
1 材料与方法 1.1 材料星洲红鱼和奥尼罗非鱼, 体长(4.8±0.2) cm, 体质量(3.01±0.12) g, 购自厦门市水产良种繁育场, 暂养5 d后选择健康无病、活力强的样品。放养密度均为1 500尾/m2, 搭配鳙50尾/m2 [体长(17.5±0.2) cm]、鲢100尾/m2 [体长(12.7±0.3) cm]、鲤50尾/m2 [体长(8.5±0.1) cm]和团头鲂(Megalobrama amblycephala) 20尾/m2 [体长(6.5±0.2) cm]。利用不同养殖品种的食性、生活水层等多种鱼类混养, 充分利用池塘饵料生物和空间形成多营养级的生态养殖模式。3口临近的池塘水深1.8~2.0 m, 面积分别为1 500、1 600和1 667 m2, 投喂粗蛋白含量为31%的配合饲料, 每天投喂2~3次, 投喂量为体重的3%~5%, 根据鱼类摄食、生长情况适当调整。每口池塘分别配备水车式增氧机2台, 养殖期间溶氧保持在3.5~5.6 mg/L, pH 6.8~7.0, 水温23.6~29.8 ℃, 透明度0.3~0.6 m, 日常管理按罗非鱼健康养殖技术进行。
1.2 生长实验体长、体质量测定和含肉率的测定实验开始后, 每隔30 d每池每种鱼抽样20尾测定体长和体质量, 180 d后每种鱼随机抽样60尾经暂养稳定后, 随机取体质均健壮、无伤病的星洲红鱼和奥尼罗非鱼各30尾, 先将鱼体用干纱布擦干, 依次编号, 分别测定其体长和体质量, 按常规方法分别除去其鳃、鳞、鳍、皮肤、内脏、骨骼及其他非肉质部分, 骨骼经煮、清洗后, 自然干燥并称重, 再用减量法计算出鱼体纯肉重。
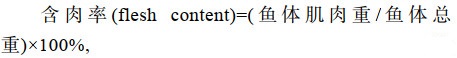 (1)
(1)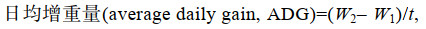 (2)
(2)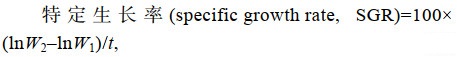 (3)
(3)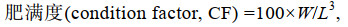 (4)
(4)式中, W1为初始平均体质量(g); W2为终末平均体质量(g); t为饲养天数; W为体质量(g); L为体长。
1.3 肌肉营养成分测定方法 1.3.1 常规营养成分将鱼肉用组织捣碎机充分捣碎搅拌均匀, 然后随机取样用于肌肉水分(GB 5009.3—85)、粗蛋白(GB 5009.5—85)、粗脂肪(GB 5009.6—85)和灰分(GB 5009.4—85)的分析。
1.3.2 氨基酸氨基酸的测定采用盐酸水解法(GB/T 5009.127—2003), 用氨基酸自动分析仪测定。
1.3.3 脂肪酸脂肪酸的测定采用气相色谱法(GB/T 9695.2—2008)。
1.3.4 肌肉营养价值评价根据联合国粮食与农业组织(FAO)和世界卫生组织(WHO)1973年建议的每克氮氨基酸评分标准模式和中国预防医学科学院营养与食品卫生研究所提出的鸡蛋蛋白质评分模式, 将所测得的必需氨基酸含量换算成每克氮中含氨基酸毫克数进行比较(乘以62.5), 分别计算氨基酸评分(AAS)、化学评分(CS)和必需氨基酸指数(EAAI)。
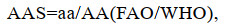 (5)
(5)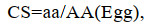 (6)
(6)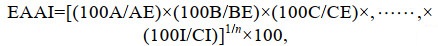 (7)
(7)式中, aa为实验样品氨基酸含量(%); AA(FAO/WHO)为FAO/WHO评分标准模式中同种氨基酸含量(%); AA(Egg)为全鸡蛋蛋白质中同种氨基酸含量(%); n为比较的必需氨基酸个数; A、B、C、…、I为鱼肌肉蛋白质的必需氨基酸含量(%), AE、BE、CE、…, IE为全鸡蛋蛋白质的必需氨基酸含量(%)。
1.4 数据处理实验所得数据用平均值±标准误(mean±SE)表示, 采用SPASS18.0软件处理, 在单因子方差分(ANOVA)基础上采用Duncan氏多重比较法检验组间差异, 显著水平为P < 0.05, 极显著水平为P < 0.01。
2 结果与分析 2.1 两种罗非鱼的可食部分与和肥满度由表 1可知, 星洲红鱼含肉率为64.21%, 高于奥尼罗非鱼9.65%, 且差异显著(P < 0.05); 内脏和头、鳃、骨、鳍、鳞含量各为11.83%和23.96%, 分别低于奥尼罗非鱼3.90% (P > 0.05)和17.75% (P < 0.05); 两种罗非鱼肥满度无显著差异(P > 0.05)。
表 1 两种罗非鱼可食部分与和肥满度的比较(n=30) Tab. 1 Comparison in flesh content and condition factor between two species of tilapia (n=30)
| 种类 | 含肉率/% | 内脏/% | 头、鳃、骨、鳍、鳞/% | 肥满度/% |
| 星洲红鱼 | 64.21±3.23a | 11.83±2.29a | 23.96±3.18a | 2.87±0.85a |
| 奥尼罗非鱼 | 58.56±4.05b | 12.31±1.64a | 29.13±4.25b | 2.84±1.02a |
| 注: 上标不同宇母表示差异显著(P < 0.05)。下同 | ||||
由图 1可知, 在同一池塘中, 两种罗非鱼生长在养殖早期60 d前无差异, 60 d以后差异明显分化, 星洲红鱼体长、体质量在第60、90、120、150和180 d分别比奥尼罗非鱼快12.16%和43.33%、15.70%和87.76%、17.13%和76.62%、22.19%和94.24%、17.13%和54.17%, 且差异显著(P < 0.05)。
 |
| 图 1 星洲红鱼与奥尼罗非鱼体长(a)和体质量(b)生长的比较 Fig. 1 Comparison in body length (a) and body weight (b) between two tilapia species |
两种罗非鱼的体长和体质量的日增长和特定生长率均差异显著(P < 0.05), 存活率分别为67.12%和69.25%, 差异无显著(表 2)。据统计, 星洲红鱼的体长(L)、体质量(W)生长与养殖天数(d)的回归方程分别为: L = –0.431 8d2 + 6.866 1d – 1.454 3 (r=0.999)和W = 2.240 6d2 + 62.8340 d – 84.290 1 (r=0.988); 奥尼罗非鱼的体长(L)、体质量(W)生长与养殖天数(d)的回归方程分别为: L = –0.355 6d2 + 5.501 5d + 0.221 4 (r=0.997)和W = 3.266 7d2 + 20.517 0d – 22.543 1 (r=0.996)。
表 2 两种罗非鱼生长性能与存活率的比较 Tab. 2 Comparison in growth performance and survival rate between two tilapia species
| 种类 | 日增长/% | 特定生长率/% | 存活率/% | |||
| 体长 | 体重 | 体长 | 体重 | |||
| 星洲红鱼 | 0.113±0.059a | 2.394±0.790a | 0.901±0.152a | 2.764±0.893a | 67.12± 8.42a | |
| 奥尼罗非鱼 | 0.098±0.049b | 1.769±1.048b | 0.838±0.167b | 2.597±0.986b | 69.25±10.42a | |
由表 3可知, 星洲红鱼和奥尼罗非鱼肌肉中水分、蛋白质、粗脂肪和灰分含量分别为78.01%和77.26%、18.21%和18.35%、2.53%和3.06%及1.25%和1.33%, 其中除粗脂肪含量差异显著(P < 0.05)外, 其他含量差异均不显著(P > 0.05)。
表 3 两种罗非鱼肌肉营养成分比较(%) Tab. 3 Comparison in proximate compositions in muscle (%) between two tilapia species
| 种类 | 水分 | 蛋白质 | 粗脂肪 | 灰分 |
| 星洲红鱼 | 78.01±1.32a | 18.21±0.75a | 2.53±0.17a | 1.25±0.87a |
| 奥尼罗非鱼 | 77.26±1.67a | 18.35±1.26a | 3.06±0.38b | 1.33±0.38a |
两种罗非鱼肌肉均由17种氨基酸(色氨酸在水解过程中易被破坏, 故未测定)组成, 氨基酸总量分别为18.13%和18.27%, 除精氨酸含量有显著差异(P < 0.05)外, 其他各氨基酸含量稍有不同, 但差异不显著。星洲红鱼和奥尼罗非鱼肌肉中必需氨基酸占其氨基酸总量分别为41.04%和40.95% (P > 0.05), 必需氨基酸含量与非必需氨基酸含量的比值分别为70.00%和69.34% (P > 0.05), 鲜味氨基酸占其氨基酸总量分别为40.71%和39.08% (P > 0.05), 必需氨基酸指数分别为67.46%和67.08% (P > 0.05) (表 4), 说明这两种罗非鱼肌肉的必需氨基酸含量较为丰富。
表 4 两种罗非鱼肌肉氨基酸组成和含量(%鲜样) Tab. 4 Contents of amino acid in the muscle of two tilapia species (% fresh)
| 氨基酸 | 星洲红鱼 | 奥尼罗非鱼 |
| 天门冬氨酸Asp* | 1.90±0.16a | 1.91±0.18a |
| 苏氨酸ThrΔ | 1.00±0.05a | 0.99±0.06a |
| 丝氨酸Ser | 0.73±0.04a | 0.81±0.06a |
| 谷氨酸Glu* | 2.63±0.18a | 2.56±0.16a |
| 甘氨酸Gly* | 1.58±0.10a | 1.46±0.08a |
| 丙氨酸Ala* | 1.31±0.08a | 1.20±0.07a |
| 胱氨酸CysΔ | 0.16±0.01a | 0.14±0.01a |
| 缬氨酸ValΔ | 0.84±0.04a | 0.86±0.05a |
| 蛋氨酸MetΔ | 0.53±0.03a | 0.56±0.04a |
| 异亮氨酸IleΔ | 0.83±0.03a | 0.81±0.06a |
| 亮氨酸LeuΔ | 1.42±0.07a | 1.43±0.09a |
| 酪氨酸TyrΔ | 0.58±0.03a | 0.61±0.02a |
| 苯丙氨酸PheΔ | 1.07±0.03a | 0.82±0.04a |
| 赖氨酸LysΔ | 1.51±0.04a | 1.54±0.06a |
| 组氨酸His | 0.47±0.01a | 0.51±0.03a |
| 精氨酸Arg | 0.63±0.05a | 0.90±0.05b |
| 脯氨酸Pro | 0.92±0.04a | 0.86±0.03a |
| ΣTAA | 18.13±1.15a | 18.27±1.21a |
| EAA/TAA | 41.04±1.22a | 40.95±1.10a |
| EAA/NEAA | 70.00±2.21a | 69.34±1.90a |
| DAA/TAA | 40.71±1.03a | 39.08±0.98a |
| EAAI | 67.46±3.52a | 67.08±2.98a |
| 注: Δ为必需氨基酸(EAA); *为呈鲜味氨基酸(DAA); ΣTAA为氨基酸总量; ΣEAA为必需氨基酸总量; ΣDAA为鲜味氨基酸总含量; ΣNEAA为非必需氨基酸总量; EAAI为必需氨基酸指数 | ||
两种罗非鱼的各种氨基酸评分(ASS)均大于0.60以上, 但在化学评分星洲红鱼和奥尼罗非鱼肌肉中蛋氨酸+胱氨酸和缬氨酸的(CS)小于0.5, 两种罗非鱼的第一限制性氨基酸均为蛋氨酸+胱氨酸, 第二限制性氨基酸为缬氨酸(表 5)。
表 5 两种罗非鱼肌肉必需氨基酸组成的评价 Tab. 5 Evaluation of essential amino acid composition in muscles of two tilapia species
| 必需氨基酸 | 鸡蛋蛋白/(mg/g) | FAO/WHO /(mg/g) | 星洲红鱼/(mg/g) | 奥尼罗非鱼/(mg/g) | |||||
| 氨基酸 | AAS | CS | 氨基酸 | AAS | CS | ||||
| 异亮氨酸Ile | 331 | 250 | 184.38 | 0.74 | 0.56 | 180.63 | 0.72 | 0.55 | |
| 亮氨酸Leu | 534 | 440 | 315.63 | 0.72 | 0.59 | 318.12 | 0.72 | 0.60 | |
| 赖氨酸Lys | 441 | 340 | 335.62 | 0.99 | 0.76 | 341.88 | 1.01 | 0.78 | |
| 苏氨酸Thr | 292 | 250 | 220.63 | 0.88 | 0.76 | 220.00 | 0.88 | 0.75 | |
| 缬氨酸Val | 411 | 310 | 188.12 | 0.61 | 0.46 | 221.88 | 0.61 | 0.46 | |
| 蛋+胱氨酸(Met+Cys) | 386 | 220 | 151.25 | 0.69 | 0.39 | 156.88 | 0.71 | 0.41 | |
| 苯丙+酪氨酸(Phe+Tyr) | 565 | 380 | 365.62 | 0.96 | 0.65 | 316.25 | 0.83 | 0.56 | |
两种罗非鱼肌肉均含有十一种脂肪酸, 三种饱和脂肪酸(SFA) C14:0、C16:0和C18:0, 八种不饱和脂肪酸(UFA), 其中单不饱和脂肪酸(MUFA) C16:1、C18:1和C20:1三种, 多不饱和脂肪酸(PUFA) C18:2、C18:3、C20:5、C22:5和C22:6五种。
由表 6可知, 每100 mg星洲红鱼和奥尼罗非鱼的肌肉中的∑SFA、∑MUFA和∑PUFA分别为28.03、38.50、25.37 mg和28.60、38.90、22.48 mg, 在SFA中棕榈酸均以(C16:0)含量最高, MUFA中油酸(C18:1)含量最高, PUFA中亚油酸(C18:2)含量最高。两种罗非鱼肌肉十一种脂肪酸中除C14:0和C18:2含量差异显著(P < 0.05)外, 其余脂肪酸含量差异不显著(P > 0.05)。
表 6 两种罗非鱼肌肉脂肪酸组成与含量(单位: %) Tab. 6 Fatty acid compositions (unit: %) in muscles of two tilapia species
| 脂肪酸 | 星洲红鱼 | 奥尼罗非鱼 |
| C14:0 | 2.48±0.07a | 1.80±0.06b |
| C16:0 | 20.60±0.32a | 21.70±0.36a |
| C18:0 | 4.95±0.05a | 5.10±0.07a |
| C16:1 | 3.15±0.12a | 3.50±0.15a |
| C18:1 | 33.75±0.52a | 33.90±0.62a |
| C20:1 | 1.60±0.06a | 1.50±0.05a |
| C18:2 | 22.25±0.26a | 19.40±0.19b |
| C18:3 | 1.45±0.08a | 1.40±0.07a |
| C20:5 | 0.09±0.01a | 0.10±0.02a |
| C22:5 | 0.83±0.02a | 0.80±0.02a |
| C22:6 | 0.75±0.01a | 0.78±0.02a |
| 其他 | 8.10±0.21a | 10.02±0.18b |
星洲红鱼体色鲜艳, 呈粉红色, 奥尼罗非鱼体色较深, 以黑色为主, 这两种罗非鱼体色差别明显。王成辉等(2004)研究认为瓯江彩鲤(Oujiang color carp)的体色与生长性能存在一定程度的相关性; 同样, 朱丽艳等(2013)和钱永生等(2019)分别在群体间(交配组合)和家系内均发现具有大块黑斑的“大花”和“粉花”个体具有生长优势。而陈芳等(2009)和吴秀林(2016)在不同体色黄鳝(Monopterus albus)生长的研究中, 发现黄斑鳝无论绝对增重、增重倍数、体重绝对生长率均优于青黄斑鳝和青斑鳝, 在体长绝对生长率、体重绝对生长率和群体生长表现为深黄大斑鳝 > 浅黄细斑鳝 > 青灰鳝。就罗非鱼而言, 公翠萍(2015)对“粉白”、“红斑”、“黑斑”3种体色红罗非鱼的生长性状研究发现, 3种体色红罗非鱼生长性状的变化趋势大致相同, 但不同体色间生长存在一定差异, 粉白红罗非鱼生长最快, 红斑及黑斑红罗非鱼相对较慢, 而且粉白红罗非鱼在初始阶段就表现出明显的生长优势。何金钊等(2016)在3口相同条件的池塘中对3个不同品系红罗非鱼生长对比发现, 粉红色的佛罗里达品系和珍珠白品系在生长速度、起捕平均规格、单位面积净产量和肥满度方面均高于橘红色的关岛品系。而对不同体色罗非鱼生长的比较研究表明尼罗罗非鱼的生长速度、起捕规格、净产量和肥满度均高于台湾红罗非鱼(李家乐等, 2002)。本实验中, 星洲红鱼和奥尼罗非鱼生长在养殖早期无明显差异, 养殖60 d以后星洲红鱼生长优势明显, 差异显著(P < 0.05), 这与许洪玉(2007)试验结果一致。有研究表明不同体色间的生长差异具有其相应的遗传和生理特性(马细兰等, 2015; 钱永生等, 2019), 而且体色多样性与物种的生长、存活、择偶偏好等性状显著相关(Shikano, 2005; Estep et al, 2006; 陈芳等, 2009; 公翠萍, 2015), 至于两种罗非鱼的体色与生长间是否存在基因连锁关系, 尚有待进一步研究。
3.2 不同鱼类含肉率的比较含肉率是衡量鱼类品质、生产性能的重要指标之一, 因鱼的种类、形态特征不同而异。如鳙, 俗称“胖头鱼”, 其含肉率在“四大家鱼”中相对较低(表 7), 因此不同罗非鱼养殖品种的形态特征各有差异, 其含肉率差异明显。同时, 含肉率也因生活环境、饲料营养、养殖模式等的不同而异。在同一池塘中, 各种环境条件、管理方法一致, 可有效地将环境因素对评价效果的影响减少到最低限度。含肉率的差异与养殖品种密切相关, 试验中星洲红鱼的含肉率变动范围在61.06%~67.51%, 奥尼罗非鱼的含肉率变动范围在54.50%~62.57%, 星洲红鱼含肉率高于奥尼罗非鱼9.65% (P < 0.05), 表现出较好的利用价值, 这与许洪玉(2007)试验结果一致。
表 7 几种淡水鱼类含肉率的比较 Tab. 7 Comparison in flesh content of different freshwater fishes
| 种类 | 含肉率/% | 种类 | 含肉率/% | |
| 星洲红鱼* | 64.21 | 草鱼Ctenopharyngodon idellus | 64.38 | |
| 奥尼罗非鱼* | 58.56 | 鲢Hypophthalmichthys molitrix | 61.52 | |
| 新吉富罗非鱼New GIFT | 71.28 | 鳙Aristichthys nobilis | 57.73 | |
| 尼罗罗非鱼O. niloticus | 67.18 | 鲤Cyprinus carpio | 66.50 | |
| 莫桑比克罗非鱼O. mossambicus | 64.05 | 鲫Carassius auratus | 63.63 | |
| 红罗非鱼Red Tilapia | 66.46 | 鳜Siniperca spp. | 67.62 | |
| 注: *为本实验品种, 下同。表内文献引用: 新吉富罗非鱼(马国红等, 2008); 尼罗罗非鱼, 莫桑比克罗非鱼(胡玖等, 1982); 红罗非鱼(陈寅山等, 2003); 草鱼, 鲢(姜巨峰等, 2010); 鳙(王金娜等, 2013); 鲤, 鲫(刘健康, 1990); 鳜(严安生等, 1995) | ||||
研究表明, 除尼罗罗非鱼(郝淑贤等, 2007)肌肉含水量高、蛋白质低、脂肪低外, 几种红罗非鱼(包括星洲红鱼) (陈寅山等, 2003; 许洪玉, 2007; 郝淑贤等, 2007; 赵何勇等, 2018)肌肉营养成分与奥尼罗非鱼(严安生等, 1995; 许洪玉, 2007; 王煜坤等, 2018; 肖炜等, 2020)、奥利亚罗非鱼(郝淑贤等, 2007)和吉富罗非鱼(郝淑贤等, 2007)相近(表 8)。郝淑贤等(2007)分别测定了奥利亚、吉富、尼罗、红罗非鱼和奥尼罗非鱼等5种罗非鱼肌肉的氨基酸组成和含量, 发现这5种罗非鱼肌肉中蛋白质的氨基酸组成基本一致。
表 8 罗非鱼与常规鱼类常规肌肉常规营养成分比较(单位: %) Tab. 8 Comparison in nutritional components in muscle among different tilapias and common fishes (unit: %)
| 种类 | 水分 | 蛋白质 | 粗脂肪 | 灰分 |
| 星洲红鱼* | 78.01±1.32a | 18.21±0.75a | 2.53±0.17a | 1.25±0.87a |
| 奥尼罗非鱼* | 77.26±1.67a | 18.35±1.26a | 3.06±0.38b | 1.33±0.38a |
| 星洲红鱼Singapore red tilapia | 74.00 | 17.80 | 5.54 | 1.66 |
| 红罗非鱼1 Red Tilapia | 78.48 | 17.68 | 2.34 | 1.08 |
| 红罗非鱼2 Red Tilapia | 79.70 | 17.30 | 1.08 | 0.79 |
| 关岛红罗非鱼Guam red tilapia | 77.84 | 20.46 | 1.05 | 1.19 |
| 奥尼罗非鱼1 O. niloticus♀× O. aureus♂ | 77.90 | 18.70 | 2.65 | 1.09 |
| 奥尼罗非鱼2 O. niloticus♀× O. aureus♂ | 77.40 | 20.76 | 1.89 | 1.14 |
| 奥尼罗非鱼3 O. niloticus♀× O. aureus♂ | 79.51 | 16.15 | 2.65 | 1.00 |
| 奥尼罗非鱼4 O. niloticus♀× O. aureus♂ | 74.30 | 18.70 | 4.40 | 1.40 |
| 尼罗罗非鱼O. niloticus | 80.85 | 15.38 | 1.75 | 1.07 |
| 奥利亚罗非鱼O. aureus | 78.53 | 18.01 | 2.26 | 1.19 |
| 吉富罗非鱼GIFT | 79.05 | 17.03 | 2.27 | 1.12 |
| 草鱼Ctenopharyngodon idellus | 81.59 | 15.94 | 0.62 | 1.22 |
| 鲢Hypophthalmichthys molitrix | 79.10 | 18.10 | 0.60 | 1.17 |
| 鳙Aristichthys nobilis | 80.03 | 18.06 | 0.70 | 1.21 |
| 鲤Cyprinus carpio | 79.58 | 16.52 | 2.06 | 1.18 |
| 鲫Carassius auratus | 80.28 | 15.74 | 1.58 | 1.64 |
| 注: *为本实验品种。表内文献引用: 星洲红鱼(许洪玉, 2007); 红罗非鱼1 (郝淑贤等, 2007); 红罗非鱼2 (陈寅山等, 2003); 关岛红罗非鱼(赵何勇等, 2018); 奥尼罗非鱼1 (肖炜等, 2020); 奥尼罗非鱼2 (王煜坤等, 2018); 奥尼罗非鱼3 (胡玖等, 1982); 奥尼罗非鱼4 (许洪玉, 2007); 尼罗罗非鱼, 奥利亚罗非鱼, 吉富罗非鱼(郝淑贤等, 2007); 鲢(贾成霞等, 2019); 鳙(王金娜等, 2013); 草鱼, 鲤, 鲫(刘健康, 1990) | ||||
本实验中星洲红鱼肌肉脂肪比奥尼罗非鱼少, 差异明显, 其他水分、蛋白质和灰分含量基本相近(表 8), 这可能是由于星洲红鱼生活在水的中上层, 比较活跃, 生活空间广, 而奥尼罗非鱼多在水体中下层生活, 活动空间小, 能量消耗少, 脂肪储存多, 从而造成一定的差异, 具体原因有待进一步研究。两种罗非鱼蛋白质含量高, 适口性良好。
两种罗非鱼肌肉氨基酸除精氨酸含量有差异外, 其他各氨酸含量稍有不同, 但差异不显著(表 9)。星洲红鱼和奥尼罗非鱼肌肉EAA/TAA和EAA/NEAA分别为41.04%和40.95%, 70.00%和69.34%, 均符合FAO/WHO的蛋白质组成理想模式, 即EAA/TAA为40%和EAA/NEAA在60%以上, 因此, 两种罗非鱼肌肉的必需氨基酸含量较为丰富, 符合人体对氨基酸的营养需求。
表 9 罗非鱼与常规鱼类肌肉氨基酸和脂肪酸组成比较(单位: %) Tab. 9 Comparison in amino acid and fatty acid compositions in muscle among different tilapias and common fishes (unit: %)
| 种类 | EAA-TAA | EAA-NEAA | DAA-TAA | SFA | MUFA | PUFA |
| 星洲红鱼* | 41.04 | 70.00 | 40.71 | 28.03 | 38.50 | 25.37 |
| 奥尼罗非鱼* | 40.95 | 69.34 | 39.08 | 28.60 | 38.90 | 22.48 |
| 星洲红鱼Singapore red tilapia | 46.29 | 96.29 | 40.84 | — | — | — |
| 红罗非鱼1 Red Tilapia | 50.50 | 105.66 | 36.78 | 35.20 | 20.80 | 37.10 |
| 红罗非鱼2 Red Tilapia | 43.74 | 73.81 | 38.54 | 31.66 | 38.75 | 29.60 |
| 关岛红罗非鱼Guam red tilapia | 41.38 | 70.59 | 38.41 | 34.90 | 31.71 | 33.38 |
| 奥尼罗非鱼1 O. niloticus♀×O. aureus♂ | 40.99 | 69.46 | 39.11 | — | — | — |
| 奥尼罗非鱼2 O. niloticus♀×O. aureus♂ | 40.80 | 68.92 | 39.06 | 33.45 | 27.44 | 39.29 |
| 奥尼罗非鱼3 O. niloticus♀×O. aureus♂ | 50.45 | 101.82 | 37.06 | 36.20 | 20.00 | 35.20 |
| 尼罗罗非鱼O. niloticus | 51.15 | 104.70 | 36.22 | 36.90 | 18.00 | 38.30 |
| 奥利亚罗非鱼O. aureus | 51.57 | 106.49 | 36.10 | 37.20 | 17.80 | 35.80 |
| 吉富罗非鱼GIFT | 50.50 | 102.03 | 36.78 | 35.90 | 20.80 | 37.10 |
| 草鱼Ctenopharyngodon idellus | 40.18 | 67.10 | 36.45 | 21.87 | 54.80 | 23.45 |
| 鲢Hypophthalmichthys molitrix | 40.16 | 104.00 | 38.53 | 32.58 | 16.91 | 50.07 |
| 鳙Aristichthys nobilis | 40.00 | 104.00 | 38.45 | 31.93 | 26.13 | 41.94 |
| 建鲤Jian carp | 38.05 | 74.23 | 36.15 | 25.02 | 29.64 | 45.12 |
| 注: *为本实验品种。表内文献引用: 星洲红鱼(许洪玉, 2007); 红罗非鱼1 (郝淑贤等, 2007); 红罗非鱼2 (陈寅山等, 2003); 关岛红罗非鱼(赵何勇等, 2018); 奥尼罗非鱼1 (王煜坤等, 2018); 奥尼罗非鱼2 (黄永春等, 2011); 奥尼罗非鱼3, 尼罗罗非鱼, 奥利亚罗非鱼, 吉富罗非鱼(郝淑贤等, 2007); 草鱼(刘建康, 1990); 鲢, 鳙(贾成霞等, 2019); 建鲤(王兵兵等, 2015) | ||||||
本实验星洲红鱼肌肉和奥尼罗非鱼以及其他学者研究的罗非鱼的EAA/TAA和DAA/TAA均优于淡水主要养殖鱼类, 其必需氨基酸含量丰富、组成平衡、营养水平高、风味鲜美、属优质动物蛋白。星洲红鱼、奥尼罗非鱼以及其他罗非鱼肌肉中脂肪酸含量、组成与淡水主要养殖鱼类无明显差异, 但星洲红鱼、奥尼罗非鱼以及其他罗非鱼肌肉中水分含量低, 蛋白质含量、脂肪含量均衡的优势明显, 作为优质鱼类广泛分布在我国华南地区养殖, 成为我国主要的淡水养殖鱼类之一。
4 结论本研究利用星洲红鱼和奥尼罗非鱼的不同体色, 将两种鱼混养在同一池塘中, 在相同的环境条件和养殖管理下研究其生长特性并对其肌肉营养成分进行分析研究。结果表明两种罗非鱼生长在前60 d无差异, 60 d后星洲红鱼体长和体质量生长比奥尼罗非鱼快, 其日增长和特定生长率均高于奥尼罗非鱼。两种罗非鱼生长快、营养丰富, 味道鲜美, 属于食用价值高的优质淡水鱼类养殖品种, 其中星洲红鱼含肉率高, 生长快, 养殖效益更为明显。
参考文献
马国红, 张延华, 师吉华, 等. 2008. 新吉富罗非鱼的含肉率及营养价值评定. 长江大学学报(自然科学版), 5(4): 35-37, 44 DOI:10.3969/j.issn.1673-1409-C.2008.04.011 |
马细兰, 张勇, 陈勇智, 等. 2015. 性类固醇激素E2、MT对尼罗罗非鱼(Oreochromis niloticus)雌、雄生长差异的影响. 海洋与湖沼, 46(6): 1487-1493 |
王成辉, 李思发, 曾伟光, 等. 2004. 瓯江彩鲤体色与生长的遗传-环境互作分析. 上海水产大学学报, 13(2): 103-106 |
王兵兵, 孙海坤, 孙建富, 等. 2015. 不同地域黄河鲤、建鲤肌肉生化组成的比较研究. 水产科学, 34(9): 540-545 |
王金娜, 唐黎, 刘科强, 等. 2013. 人工养殖与野生鳙鱼肌肉营养成分的比较分析. 河北渔业, 16(2): 8-14, 16 DOI:10.3969/j.issn.1004-6755.2013.02.003 |
王辉, 强俊, 李瑞伟. 2008. 温度对奥尼罗非鱼仔稚鱼生长、饲料利用和消化酶活力的影响. 广东海洋大学学报, 28(6): 14-19 DOI:10.3969/j.issn.1673-9159.2008.06.004 |
王煜坤, 郝淑贤, 李来好, 等. 2018. 不同地区、品种及养殖模式罗非鱼营养差异分析. 食品工业科技, 39(12): 231-237 |
公翠萍, 2015. 不同体色红罗非鱼生长性能比较及遗传多样性研究[D]. 南京: 南京农业大学: 9-16.
|
乐贻荣, 肖炜, 邹芝英, 等. 2015. 奥尼罗非鱼肌肉营养成分分析和营养价值评定. 中国农学通报, 31(11): 88-93 DOI:10.11924/j.issn.1000-6850.2014-2485 |
朱丽艳, 马玉清, 项松平, 等. 2013. 不同体色瓯江彩鲤生长动态的观察与分析. 上海海洋大学学报, 22(3): 341-348 |
刘健康. 1990. 东湖生态学研究(一). 北京: 科学出版社, 307-331
|
许洪玉. 2007. 星洲红鱼和奥尼罗非鱼养殖对比试验. 齐鲁渔业, 24(6): 37-39 |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会. 2020. 中国渔业统计年鉴-2020. 北京: 中国农业出版社, 21-36
|
严安生, 熊传喜, 钱健旺, 等. 1995. 鳜鱼含肉率及鱼肉营养价值的研究. 华中农业大学学报, 14(1): 80-84 |
李家乐, 李思发, 韩风进. 2002. 台湾红罗非鱼和尼罗罗非鱼的生长特性与养殖效果的比较. 上海水产大学学报, 11(1): 1-5 |
杨弘, 吴婷婷, 夏德全. 2005. 三种奥尼杂交罗非鱼养殖效果比较. 科学养鱼, (3): 18 |
肖炜, 李大宇, 邹芝英, 等. 2012. 四种杂交组合奥尼罗非鱼及其亲本的生长对比研究. 水生生物学报, 36(5): 905-912 |
肖炜, 李大宇, 祝琳, 等. 2020. 4种尼罗罗非鱼♀×奥利亚罗非鱼♂杂交子代肌肉营养成分对比分析. 中国农学通报, 36(8): 139-144 |
吴秀林, 2016. 不同体色黄鳝生长性能、营养价值评价及遗传多样性研究[D]. 南京: 南京农业大学: 11-30.
|
何金钊, 陈诏, 滕云, 等. 2016. 不同品系红罗非鱼池塘养殖生长对比试验. 南方农业, 10(3): 181-182 DOI:10.3969/j.issn.1673-890X.2016.03.108 |
张宇航, 高扬, 李文红, 等. 2020. 低温停食和复温后投喂频率对奥尼罗非鱼幼鱼生长的影响. 西南农业学报, 33(9): 2125-2131 |
陈芳, 杨代勤, 苏应兵. 2009. 3种不同体色黄鳝生长速度的比较. 长江大学学报(自然科学版), 6(3): 33-34, 38 |
陈家长, 孟顺龙, 胡庚东, 等. 2010. 空心菜浮床栽培对集约化养殖鱼塘水质的影响. 生态与农村环境学报, 26(2): 155-159 DOI:10.3969/j.issn.1673-4831.2010.02.011 |
陈寅山, 戴聪杰. 2003. 红罗非鱼肌肉的营养成分分析. 福建师范大学学报(自然科学版), 19(4): 62-66 DOI:10.3969/j.issn.1000-5277.2003.04.015 |
林仕梅, 麦康森, 谭北平. 2007. 菜粕、棉粕替代豆粕对奥尼罗非鱼(Oreochromis niloticus×O. aureus)生长、体组成和免疫力的影响. 海洋与湖沼, 38(2): 168-173 DOI:10.3321/j.issn:0029-814X.2007.02.013 |
赵何勇, 陈诏, 徐鸿飞, 等. 2018. 海水和淡水养殖关岛红罗非鱼肌肉营养成分及品质特性分析. 南方农业学报, 49(7): 1396-1402 DOI:10.3969/j.issn.2095-1191.2018.07.21 |
郝淑贤, 李来好, 杨贤庆, 等. 2007. 5种罗非鱼营养成分分析及评价. 营养学报, 29(6): 614-615, 618 DOI:10.3321/j.issn:0512-7955.2007.06.022 |
胡玖, 张中英, 吴福煌. 1982. 尼罗罗非鱼与莫桑比克罗非鱼的含肉率及鱼肉的生化分析. 淡水渔业, (4): 34-37 |
钟全福. 2019. 罗非鱼为主多品种混养池塘氮磷收支. 广东海洋大学学报, 39(6): 48-53 DOI:10.3969/j.issn.1673-9159.2019.06.007 |
姜巨峰, 王玉佩, 李春艳, 等. 2010. 3种优质淡水鱼类的含肉率、肥满度及肌肉营养成分的分析. 安徽农业科学, 38(26): 14478-14480 DOI:10.3969/j.issn.0517-6611.2010.26.114 |
贾成霞, 曲疆奇, 李永刚, 等. 2019. 密云水库鲢鱼、鳙鱼营养成分分析与评价. 水产科学, 38(1): 40-47 |
夏德全. 2003. 论罗非鱼产业发展. 海洋与渔业, (8): 7-9 |
钱永生, 陈红林, 杜金星, 等. 2019. 4种体色瓯江彩鲤的生长、摄食和呼吸特性差异及其相关性分析. 中国水产科学, 26(4): 695-702 |
黄永春. 2015. 养殖水温对星洲红鱼生长、消化和免疫的影响. 动物营养学报, 27(7): 2264-2271 DOI:10.3969/j.issn.1006-267x.2015.07.034 |
黄永春, 李文静, 林祥日, 等. 2011. 星洲红鱼形态、染色体组型及细胞核DNA含量的分析. 淡水渔业, 41(5): 3-8 DOI:10.3969/j.issn.1000-6907.2011.05.001 |
黄永春, 林祥日, 许洪玉, 等. 2012. 星洲红鱼对温度适应性的研究. 福建水产, 34(5): 398-404 DOI:10.3969/j.issn.1006-5601.2012.05.009 |
康自强, 2014. 盐度胁迫对星洲红鱼生长、消化酶和免疫因子活性及肌肉品质的影响[D]. 厦门: 集美大学: 1-42.
|
彭淇, 王斐, 吴彬, 等. 2014. 2种罗非鱼加工下脚料产物替代秘鲁鱼粉养殖奥尼罗非鱼(Oreochromis niloticus×O. aureus)稚鱼效果评价. 海洋与湖沼, 45(3): 602-607 |
强俊, 王辉, 李瑞伟, 等. 2009. 饲喂频率对奥尼罗非鱼仔稚鱼生长、体成分和消化酶活力的影响. 广东海洋大学学报, 29(4): 79-83 DOI:10.3969/j.issn.1673-9159.2009.04.018 |
ESTEP L K, SHAWKEY M D, HILL G E, 2006. Carotenoid-based breast plumage colour, body condition and clutch size in red fodies (Foudia madagascariensis). Ostrich, 77(3/4): 164-169 |
FLORIO D, GUSTINELLI A, CAFFARA M et al, 2009. Veterinary and public health aspects in tilapia (Oreochromis niloticus niloticus) aquaculture in Kenya, Uganda and Ethiopia. Ittiopatologia, 6: 51-93 |
MA J T D Y, RYE M, WANG Y X et al, 2012. Genetic improvement of tilapias in China: Genetic parameters and selection responses in fillet traits of Nile tilapia (Oreochromis niloticus) after six generations of multi-trait selection for growth and fillet yield. Aquaculture, 366/367: 67-75 DOI:10.1016/j.aquaculture.2012.08.028 |
PONZONI R W, NGUYEN N H, KHAW H L et al, 2011. Genetic improvement of Nile tilapia (Oreochromis niloticus) with special reference to the work conducted by the World Fish Center with the GIFT strain. Reviews in Aquaculture, 3(1): 27-41 DOI:10.1111/j.1753-5131.2010.01041.x |
SHIKANO T, 2005. Marker-based estimation of heritability for body color variation in Japanese flounder Paralichthys olivaceus. Aquaculture, 249(1/4): 95-105 |
TRọNG T Q, MULDER H A, VAN ARENDONK J A M et al, 2013a. Heritability and genotype by environment interaction estimates for harvest weight, growth rate, and shape of Nile tilapia (Oreochromis niloticus) grown in river cage and VAC in Vietnam. Aquaculture, 384/387: 119-127 DOI:10.1016/j.aquaculture.2012.12.022 |
TRọNG T Q, VAN ARENDONK J A M, KOMEN H, 2013b. Genetic parameters for reproductive traits in female Nile tilapia (Oreochromis niloticus): II. Fecundity and fertility. Aquaculture, 416/417: 72-77 DOI:10.1016/j.aquaculture.2013.08.031 |