 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 李琦, 陈朝晖. 2022.
- LI Qi, CHEN Zhao-Hui. 2022.
- 基于ADCP回声的黑潮-亲潮混合区浮游动物昼夜垂直迁移研究
- DIEL VERTIAL MIGRATION OF ZOOPLANKTON IN THE KUROSHIO-OYASHIO MIXED ZONE BASED ON ADCP ECHO
- 海洋与湖沼, 53(2): 305-319
- Oceanologia et Limnologia Sinica, 53(2): 305-319.
- http://dx.doi.org/10.11693/hyhz20211000250
文章历史
-
收稿日期:2021-10-18
收修改稿日期:2021-11-28




2. 青岛海洋科学与技术试点国家实验室 山东青岛 266237
2. Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China
浮游动物的昼夜垂直迁移(diel vertical migration, DVM)是海洋和淡水生态系统中普遍存在的现象, 代表物种有海洋桡足类和淡水枝角类(刘顺会等, 2008)。浮游动物进行DVM的原因是多方面的, 主要是为了逃避捕食者, 以保证自身的生存和种群的维持(张武昌, 2000; Brierley, 2014; Thygesen et al, 2019)。常规类型的DVM表现为昼沉夜浮: 浮游动物夜晚上升到表层摄食, 白天下沉到深层栖息。海洋浮游动物通常从200~1 000 m的海洋中层长距离迁移数百米至200 m以上的海洋表层, 该现象普遍发生在世界各大洋, 是地球上最大规模的生物迁移(Behrenfeld et al, 2019)。作为食物链重要的一环, 浮游动物摄食浮游植物并被鱼类捕食(刘镇盛等, 2013), 通过DVM将海洋上层浮游植物固定的碳输送到深层, 然后在深层呼吸、代谢、排遗和被捕食, 成为碳的垂向输送以及海洋中层能量和物质补充的重要途径, 对海洋生态和生物地球化学循环有重要的影响(Doney et al, 2013; 孙栋等, 2017; Archibald et al, 2019)。DVM现象在约200 a前首次被观察到, 现今的研究手段包括浮游生物拖网、沉积物捕获器捕获、光学和声学设备探测等(孙栋等, 2017)。前两种研究方式效率低、范围窄且非实时, 光学和声学研究通过利用回波可进行大范围的高效探测。光波在海水中的穿透深度很浅, 仅限于海洋表层观测; 而声波的水体穿透能力强, 可对立体的海洋进行全方位探测, 因此被广泛应用于海洋渔业和生态学研究。早在二战期间, 人们利用声呐等声学仪器发现大洋中广泛存在声散射层, 在海洋中层和表层间发生垂直移动且日变化显著, 后续研究表明该现象与中层海洋动物的DVM有关(Benoit-Bird et al, 2016)。
当前, 海洋观测中常用的声学设备包括鱼探仪、声学多普勒流速剖面仪(acoustic doppler current profiler, ADCP)和其他类型的声呐, 可搭载在固定或移动平台上进行观测。其中, ADCP既可安装在船底进行走航观测, 获取大范围的回声剖面, 又可安装在潜标、浮标等固定平台上, 获取定点回声剖面的时间序列。以上两种方式可分别用于研究DVM的空间分布和时间变化。此外, ADCP同步测得的海水流速以及船载或潜、浮标同步观测的水温、营养盐、叶绿素和生物量等要素, 可为研究DVM对水文、生物和化学环境变化的响应提供数据支撑。
船载观测通常结合高低频ADCP来分辨多种尺寸的海洋动物及其DVM的空间分布。比如, Luo等(2000)结合船载150 kHz ADCP和12 kHz声呐, 发现中层鱼DVM的平均深度为3~4 cm/s, 最大速度为10~13 cm/s; 浮游动物DVM的平均速度为2 cm/s, 最大速度为8 cm/s。潜标通常搭载75 kHz ADCP, 可用于研究DVM的长时间变化和对典型海洋-大气过程(如: 中尺度涡、台风等)的响应。Inoue等(2016)基于西北太平洋副热带环流圈的生物地球化学潜标S1的观测, 发现与浮游动物DVM有关的上散射层深度随冬季混合层的加深而增加, 最深可达200 m; 中尺度涡旋过境引起混合层的加深和再层化过程可显著影响上层浮游植物的分布, 从而影响浮游动物的DVM强度。Potiris等(2018)在地中海观测到浮游动物DVM速度随风暴期间的海表降温、云量和风速的增加而减弱。Yang等(2019)发现南海北部陆坡处浮游动物的DVM强度在强台风期间减弱, 浮游动物迁移到深层以躲避强流。
国内采用声学方法研究海洋浮游动物的DVM起步较晚, 从20世纪80年代我国引入声学观测设备后才逐渐开始, 关注的海域主要为近海及边缘海上层水体, 代表性文章有: 李劳钰等(2012), 陈钊等(2016)和徐亚军等(2016)。有关深远海DVM的综合性研究较少(孙栋等, 2017)。作为距离我国最近的远洋渔业捕捞区, 黑潮-亲潮混合区(142°~160°E, 35°~40°N)营养物质丰富, 浮游动、植物的种类和数量多, 是秋刀鱼、柔鱼和鲐鱼等远洋经济鱼类重要的栖息地(马超等, 2018)。作为上述经济鱼类的饵料, 浮游动物在夜间垂直迁移到海洋上层, 吸引大量鱼群前来捕食, 有利于海洋鱼类的捕捞。因此, 厘清该海区浮游动物DVM的时空分布和强度变化, 对今后开展精准捕捞和提升捕捞效率具有重要的意义。此外, 黑潮-亲潮混合区的海洋动力过程丰富, 中纬度风暴频发(Qiu et al, 2010), 特别是黑潮延伸体主轴常年向北甩出大量反气旋式的中尺度暖涡(Sugimoto et al, 2017), 为研究水文和气象环境变化对浮游动物DVM的影响提供了良好的试验场。然而, 由于缺乏长期、系统的现场观测, 目前对该海区浮游动物DVM的基本特征及影响机制的认识较为匮乏。本文基于西北太平洋黑潮延伸体定点观测系统(Kuroshio extension mooring system, KEMS)的深海潜标所搭载的ADCP回声等数据, 综合研究了黑潮-亲潮混合区浮游动物的垂向分布、DVM的基本特征、多时间尺度变化及其对暖涡的响应。
1 数据与方法 1.1 数据介绍KEMS现有5套横跨黑潮延伸体主轴的6 000 m级深海潜标(M1~M5, 图 1a)。其中, M2潜标(39°N, 150°E)位于黑潮-亲潮混合区, 其时间跨度长(2016年4月至今)、数据完整度高。潜标主浮体位于500 m, 上面搭载了两台Long Ranger 75 kHz ADCP, 分别设置为上打和下打。ADCP的时间分辨率为1 h, 观测量程为640 m, 垂向分为37层, 每层厚度为16 m。将上下两个ADCP拼接, 可获得上1 000 m的流速和回声强度数据。此外, 在潜标100, 400, 1 500和5 500 m处还安装有温盐深仪(conductivity temperature depth, CTD), 观测的时间分辨率为5 min。为研究该海区浮游动物DVM的基本特征及其季节和季节内变化, 选取M2潜标2016年4月至2017年5月共1年的连续观测数据。该时间段内有较多的中尺度涡过境, 适于开展不同海洋动力环境对DVM影响的研究。
 |
| 图 1 2016年平均的黑潮-亲潮混合区动力和生态环境 Fig. 1 Dynamical and ecological environments in the Kuroshio-Oyashio mixed zone in 2016 注: a: 海表面高度(sea surface height, SSH); b: 海表面温度(sea surface temperature, SST); c: C表示海表面叶绿素浓度(chlorophyll, CHL), 单位: mg/m3 |
此外, 通过哥白尼海洋环境监测中心(copernicus- marine environment monitoring service, CMEMS, https://marine.copernicus.eu/)获取了2016年日平均的海表面高度(sea surface height, SSH)、海表面温度(sea surface temperature, SST)和海表面叶绿素浓度(chlorophyll, CHL), 以展示黑潮-亲潮混合区的年平均动力和生态环境特征(图 1)。
1.2 ADCP数据处理根据陈子飞等(2021)提出的深海锚系潜标ADCP数据处理方法, 依次通过旁瓣干扰、速度与良好率、横摇与纵摇和速度误差与相关幅值对数据进行质控。
ADCP测得的垂向流速通常被近似为浮游动物DVM速度。由于ADCP随主浮体存在一定的垂向运动, 所以需要将测得的垂向流速减去ADCP自身的垂向运动速度, 从而得到实际的DVM速度w (Luo et al, 2000)。通过分析发现, 上100 m的垂向速度基本一致, 故将其深度平均值作为ADCP的垂向运动速度。进一步对w进行1 d的滑动平均, 再进行样条插值, 最终得到可用的DVM速度。此外, 由于DVM期间浮游动物的位置随时间和深度变化, 只出现在ADCP的部分观测层中, 所以选取迁移时间段内的最大速度作为DVM速度, 以避免通过时间平均对DVM速度的低估(Potiris et al, 2018)。
通过直接插值连接上下两台ADCP数据会导致连接处出现偏小的数据带, 因此, 首先去除每台ADCP的第一层数据, 再进行插值得到垂向衔接平滑的剖面。由于潜标ADCP存在垂向的运动, 所以将不同时刻剖面的数据点统一插值到50~1 000 m之间间隔为10 m的深度层。最后, 将温盐剖面进行不等距的上层加密插值, 并将其时间间隔平均为1 h, 以保证与ADCP采样频率一致。
1.3 声体积后向散射强度(volumetric backscattering strength; Sv)ADCP测流原理是捕捉水中各类悬浮粒子的声波散射, 其发射的声波波长一般为mm至cm量级, 主要被尺寸与声波长度相当或稍小的浮游动物及小尺寸鱼、气泡等散射。75 kHz ADCP发射的声波波长为20 mm, 足以分辨典型的海洋浮游动物, 如: 桡足类(~1 mm)、翼足类(~1 cm)和磷虾(~1 cm)等(Teledyne RD Instruments, 2011)。散射声波的粒子浓度越大, 后向散射的回声数越多, 仪器接收到的回声强度E越大。回声强度E受水体对声波的吸收影响, 随测量层到换能器距离的增大而减弱, 无法反映悬浮粒子浓度的真实情况, 因此, 需要转换为后向散射强度Sv。研究表明, Sv和生物量成正比, 可作为表征生物量的指标(Inoue et al, 2016)。Sv的计算参考ADCP厂商TRDI给出的最新公式(Mullison, 2017):
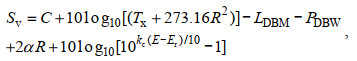 (1)
(1)公式(1)对前人给出的经典公式(Deines, 1999; Gostiaux et al, 2010)做了进一步的修正, 避免了随机误差的干扰, 适用于多种类型的后向散射水体环境。其中, C为常数; LDBM是声脉冲发射长度; PDBW是声脉冲发射功率; 均与ADCP仪器自身有关。根据Mullison (2017)提供的公式和参数, 本研究所用型号的ADCP的常数C取−166.94 dB, PDBW取23.8 dB, 而LDBM通过声脉冲长度Xmt进一步计算得到(LDBM=10log10Xmt, Xmt在WinADCP的file details中可查); Tx是换能器测的温度(℃); α为水体声吸收系数, 取典型值0.023; R是沿波束方向从测量层到换能器的声脉冲回波路径长度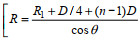
如图 2a所示, 海洋上1 000 m存在两个强声散射层, 分别主要位于200 m以浅的上层海洋和300~800 m之间的中层海洋, 对应不同种类的浮游动物。附近渔场的表层生物采样表明, 该海区浮游动物以桡足类、背囊类、毛颚类及水母类为主要组成大类, 受高温高盐黑潮水的影响, 物种以广温广盐性暖水种居多(靳少非等, 2014)。同纬度日本沿岸的浮游动物采样发现, 0~150 m之间以桡足类的真哲水蚤和太平洋长腹水蚤为主; 150~500 m之间以3种哲水蚤为主, 具有明显的垂向迁移特征(Yamaguchi et al, 2014)。
 |
| 图 2 海洋上1 000 m浮游动物和环境因子的时空分布 Fig. 2 Temporal and spatial distributions of zooplankton and environmental factors above 1000 m 注: Sv: 声体积后向散射强度, a中灰线表示ADCP的位置, 竖直虚线为四季的分界线(春: 4~6月, 夏: 7~9月, 秋: 10~12月, 冬: 1~3月)下同; T: 温度, b中黑色等值线为位势密度异常σθ (单位: kg/m3); C表示海表面叶绿素浓度, 单位: mg/m3, 取以M2潜标为中心、1°范围内的平均值 |
海洋上层和中层浮游动物的生物量和垂向分布均呈现明显的季节变化, 但两层的变化规律有所不同。海洋上层浮游动物的生物量春秋多、夏冬少, 与春秋两季叶绿素浓度的双峰分布一致(图 2a, 2c); 其分布深度夏季最浅, 位于100 m以上, 从10月开始直至次年的1月, 逐渐向下扩展到200 m。浮游动物垂向分布加深的时间和深度与Kobari等(2008)在该海区观测的叶绿素浓度最大层的变化特征一致, 表明秋冬混合层的加深能够带来深层营养盐, 使浮游植物的垂向分布向下扩展, 浮游动物的分布深度随之加深(Inoue et al, 2016)。上述分析表明, 上层海洋浮游动物的生物量和垂向分布的季节变化直接受浮游植物的调控。
海洋中层浮游动物的生物量春夏多、秋冬少, 随水温的降低而减少(图 2a, 2b)。同纬度海区上2 000 m的浮游动物采样表明, 各物种数量在500~1 500 m之间最多; 物种尺寸随深度变化, 较大的物种分布在500 m以下; 500 m以深水体中的物种数量从春季到秋季较大, 而在晚秋和早冬最小(Deevey et al, 1977)。4月底至5月初以及10~11月中旬期间, 冷水上涌, 生物量锐减, 很可能与亲潮水向南入侵有关。浮游动物分布深度的上限基本稳定在300~400 m之间, 而其下限与26.8σθ等密度线变化趋势一致, 呈现春夏深、秋冬浅的特征(图 2a, 2b)。该现象可能与春夏季水温升高, 部分物种(如: 太平洋长腹水蚤)为躲避暖水而迁移到深层有关(Sun et al, 2011)。
2.2 DVM的基本特征如图 3所示, 不同季节的Sv均呈现显著的日周期变化, 反映了浮游动物的DVM。上1 000 m存在多个DVM层, 从浅到深依次为200 m层(春夏较为明显, 图 3a, 3b), 400 m层和800 m层(春冬比较明显, 图 3a, 3d)。声学探测和生物采样结果表明, 进行DVM的浮游动物在400~500 m间主要是蛇鼻鱼, 600~800 m间主要是灯笼鱼(Ariza et al, 2016)。下面仅讨论全年都显著的400 m至表层的DVM。
 |
| 图 3 四季中间10 d的Sv Fig. 3 Sv during 10 days in the middle of the four seasons 注: a: 春季(2017年); b: 夏季(2016年); c: 秋季(2016年); d: 冬季(2017年) |
将每天相同时刻的Sv取平均, 得到全年平均的DVM模式(图 4a)。上下散射层的平均位置分别位于130 m以上和330 m以下, DVM轨迹位于130~330 m之间, DVM幅度最小为200 m。由DVM轨迹可知, 浮游动物大约在傍晚18:00~20:00从白天所在的下散射层迁移到上散射层摄食, 在天亮4:00~6:00向下迁移回到栖息层, 垂向迁移基本上均在日落和日出时刻前后2 h内完成。
 |
| 图 4 DVM的基本特征 Fig. 4 The main characteristics of DVM 注: a: 年平均的DVM模式(Sv的日变化), 红色(蓝色)虚线从左到右分别表示向下(上)迁移的起、止时刻, 黑色虚线从上到下分别表示上散射层的下界和下散射层的上界, 向下(上)迁移的轨迹主要位于红色(蓝色)虚线和黑色虚线组成的矩形框内; b: 昼(夜)平均的Sv剖面, 夜间取日落2 h后到日出2 h前的时间段, 昼间取日出2 h后到日落2 h前的时间段; c: 向上和向下的DVM速度w, 取2 h迁移时间段内的最大值 |
根据DVM起止时刻, 将一天分为昼(日出2 h后到日落2 h前)和夜(日落2 h后到日出2 h前)两个时间段, 分别得到昼和夜平均的Sv剖面(图 4b)。上散射层的Sv夜间大、白天小, 下散射层正好相反, 对应浮游动物昼沉夜浮的迁移特性。昼和夜的Sv剖面相交深度为290 m, 是上下散射层的平均分界深度。
ADCP测得的DVM速度w表明, 向上和向下的迁移速度最大可达9.0 cm/s。DVM轨迹上的全年平均上移速度在0.7~1.9 cm/s之间, 最大值位于160 m; 下移速度在1.4~2.6 cm/s之间, 最大值位于250 m (图 4c), 下移速度总体比上移速度大, 与Potiris等(2018)的观测结果一致。根据200 m的最小DVM幅度和2 h的平均DVM时长, 估计得到平均迁移速度最小为2.8 cm/s。在相应的深度范围130~330 m, ADCP测得的深度平均垂向速度为2.0 cm/s, 比估计值小约1 cm/s。这主要是因为ADCP采样频率较低(1次/h), 在DVM期间最多观测到2个剖面, 难以测到最大DVM速度, 从而低估了真实的DVM速度(Potiris et al, 2018)。已有研究表明, 浮游动物DVM的平均速度在2~5 cm/s, 最高可达10 cm/s (Luo et al, 2000; Jiang et al, 2007; Ariza et al, 2016; 徐亚军等, 2016; Potiris et al, 2018)。因此, 本文中ADCP测得的平均和最大DVM速度均在合理范围内。
2.3 DVM的多时间尺度变化及其影响因素 2.3.1 季节变化上散射层(50~290 m)深度平均的夜间Sv多用于开展DVM随时间的变化研究(Inoue et al, 2016; Yang et al, 2019)。然而, 本文处理后的数据在上150 m有不同程度的连续空缺(图 5a), 直接插值会造成高频信号的缺失。由图 6a可知, 150~250 m深度平均的夜间Sv在整个时间段内连续且与其上散射层平均值的变化趋势一致, 因此被作为DVM强度来表征进行DVM的浮游动物生物量。如图 5a和图 6a所示, DVM强度春季最强、秋冬次之、夏季最弱, 与浮游植物生物量的季节变化一致。春季光照的增加和秋冬混合层的加深都有助于浮游植物的生长, 浮游动物生物量增加(Jiang et al, 2007; Inoue et al, 2016), DVM强度随之增强。夏季, 水体层结增强, 由于营养物质的消耗和被捕食, 浮游植物生物量减少(Xing et al, 2021), 浮游动物生物量随之减少, DVM强度减弱。此外, 早秋10月DVM强度的减弱与冷水上涌引起中层浮游动物生物量锐减有关, 而不是季节变化导致的。
 |
| 图 5 DVM和相关环境因子的时空变化 Fig. 5 Temporal and spatial variations of DVM and relevant environmental factors 注: a: 夜间Sv; b: 流速V, 黑色等值线为位势密度异常σθ (单位: kg/m3); c: 温度异常Ta, 由温度减去其时间平均值得到 |
 |
| 图 6 DVM强度及其小波谱 Fig. 6 DVM strength and its wavelet spectra 注: a: DVM强度(150~250 m深度平均的夜间Sv)和50~290 m深度平均的夜间Sv的时间序列; b: DVM强度的小波谱, DVM强度先后经过了标准化和5~128 d的带通滤波, 粗黑色等值线为95%信度线, 细黑色曲线下方区域表示受边界效应影响的区域; c: DVM强度小波谱的时间平均谱 |
将不同季节每天相同时刻的Sv取平均, 得到季节平均的DVM模式(图 7), 发现DVM的幅度和起止时间也存在显著的季节变化。根据上下散射层的深度范围和DVM轨迹可知(图 5a和图 7), DVM幅度夏季最大、春季次之、秋冬较小, 与浮游植物分布深度的季节变化一致。夏季, 海洋表层水温增高, 水体层结增强, 混合层变浅, 浮游植物随营养盐的垂向分布被局限在海表(Kobari et al, 2008), 浮游动物的摄食深度变浅, 导致DVM幅度增大。秋冬, 中纬度风暴频发, 混合层加深, 营养盐上涌, 浮游植物的垂直分布范围向深处扩展(Inoue et al, 2016), 浮游动物摄食的深度范围扩大, 导致DVM幅度减小。春季, 随着温度的增加, 表层海洋快速层化, DVM幅度相比秋冬有所增大。
 |
| 图 7 季节平均的DVM模式 Fig. 7 Seasonal-averaged patterns of DVM 注: a: 春季(2017年4~6月); b: 夏季(2016年7~9月); c: 秋季(2016年10~12月); d: 冬季(2017年1~3月) |
DVM起止时间与日落和日出时刻的季节变化同步(图 8, 李劳钰等, 2012; Potiris et al, 2018)。春分和秋分昼夜等长(12 h), 日出和日落时间为6:00和18:00。从夏季到冬季, 昼长夜短逐渐变为昼短夜长, DVM的起止时刻分别逐渐提前和推迟, 浮游动物夜间摄食时间随之延长, 最长可达15 h。从冬季到夏季的变化则相反。上述特征说明浮游动物DVM是在太阳光照长期调节下形成的生物节律, 光照强度是激发DVM的信号(Bianchi et al, 2013; Brierley, 2014)。
 |
| 图 8 DVM强度和速度的时刻-日期图 Fig. 8 Hour vs day plot of DVM strength and velocity 注: a. 150~250 m深度平均的Sv, 红色和蓝色实线分别表示日出和日落时刻; b. DVM速度w, 红(蓝)色虚线从上到下分别表示日出(日落)之后和之前1 h |
如图 6所示, 小波谱表明DVM强度还具有显著的季节内变化, 谱峰值从高到低分别位于100, 60, 30和15d周期附近。季节内变化在时间上不连续, 具有偶然性发生的特点, 推测与海洋动力过程引起的环境变化有关(Jiang et al, 2007; Yang et al, 2019)。该海区位于地形平坦的深海大洋, 潮汐相对较弱(Zhao et al, 2016), 且中纬度风暴频发, 阴雨天气占主, 月光多被云遮挡, 使得海表接收到的月光强度变化微弱。因此, 月光和潮汐对DVM的影响相对较弱。相反, 该海区中尺度涡旋活跃, 能引起海洋环境的强烈变化, 从而显著影响浮游动物的行为, 使DVM呈现偶然性变化。
通过时间序列的对比可知(图 9), DVM强度的季节内变化与流速和温度有关。对各变量时间序列进行3~120 d的带通滤波, 发现DVM强度的变化与温度呈正相关, 整体同期相关系数为0.52, 在流速较弱的时间段内(如2016年4~7月)局部相关系数高达0.8。相比温度, DVM强度与流速的整体相关性较低, 但在流速剧烈增大的时期(如2016年8~10月, 2017年3~4月), DVM强度表现出随流速的增强而减弱的变化。
 |
| 图 9 DVM强度、流速V和温度异常Ta的时间序列 Fig. 9 Time series of DVM strength, current, and temperature anomaly 注: a: 流速V取150~300 m深度平均, 温度数据点较少, 取150~400 m平均(下同), 细线和粗线分别为各变量的原始和8 d滑动平均的时间序列; b: 3~120 d带通滤波后的DVM强度和流速, R为时间序列的相关系数, L表示DVM强度滞后流速(温度异常)的时间(下同); c: 3~120 d带通滤波后的DVM强度和温度异常 |
由图 5中的流速和温度异常可知, 2016年8~10月和2017年3~4月期间有暖涡过境, 下面针对这两次暖涡事件, 分析暖涡对浮游动物DVM的影响。
如图 10和图 11所示, 暖涡过境期间, DVM路径仍清晰可见, 但由于暖涡边缘和涡中心分别以强流(水平流速, 下同)和高温特征为主, 涡边缘和涡中心的DVM模式有所不同。当涡边缘的强流经过时, DVM强度出现不同程度的减弱。对于2016年的暖涡, 流速增强伴随温度的减弱, 无法排除DVM强度的减弱来自温度的减弱。对于2017年的暖涡, 流速增强期间, DVM强度显著减弱。两次暖涡中心的高温过境时, DVM强度相比强流期间都有所增强。相比强流的影响, 2016年的暖涡高温中心处的上散射层强度增强且分布深度向下扩大, 并在次表层(25.5σθ~26σθ)出现了强散射层。当2017年的暖涡高温中心经过时, 强流期间减弱的DVM强度基本恢复到正常水平。此外, 涡中心的高温引起等密度面的下凹, 上散射层(次表层散射层)下凹程度大于下散射层, 从而导致DVM幅度有所减弱。
 |
| 图 10 暖涡过境期间(2016年8月20日至10月14日)DVM和环境因子的时空变化 Fig. 10 Temporal and spatial variations of DVM and the environmental factors during the pass of the warm eddy form August 20 to October 14, 2016 注: 黑色等值线为位势密度异常σθ (单位: kg/m3); d: 3~120 d带通滤波后的DVM强度、流速和温度异常的时间序列 |
 |
| 图 11 暖涡过境期间(2017年3月15日至4月15日) DVM和环境因子的时空变化 Fig. 11 Temporal and spatial variations of DVM and the environmental factors during the pass of the warm eddy form March 15 to April 15, 2017 注: a, b, c中黑色等值线为位势密度异常σθ (单位: kg/m3); d: 3~120 d带通滤波后的DVM强度、流速和温度异常的时间序列 |
以上两个暖涡的个例分析进一步说明, DVM强度与温度呈正相关, 且还受到强流的削弱作用。暖涡总体上抑制DVM, 但涡旋不同位置对其影响不同: 暖涡边缘的强流减弱DVM强度, 而暖涡中心的高温减小DVM幅度。
3 讨论 3.1 温度和流速对DVM的影响DVM的变化受多种因素包括内因(性别、年龄、体长和生理节律等)和外因(光照、摄食者、饵料丰度、流、温度、盐度、氧气等)的影响(张武昌, 2000)。温度和流速一般是通过影响浮游植物间接影响浮游动物。因此, 温度或流速与DVM强度之间可能存在一定的相关性, 但不能代表二者之间存在直接的因果关系。比如, Jiang等(2007)发现, 浮游动物生物量与温度成反比, 温度升高伴随着叶绿素浓度的降低, 浮游动物生物量随之下降。Yang等(2019)观测到, 台风期间流速增大, 温度下降, 浮游动物DVM减弱。而本文研究结果表明, 在季节尺度内, 温度升高(降低), DVM强度随之增强(减弱); 而水平流速增强, DVM强度随之减弱。
3.2 中尺度涡对DVM的影响中尺度涡在海洋中普遍存在, 对浮游动物分布和群落结构有重要影响。一方面, 中尺度涡裹挟着封闭的水体移动, 将其中的浮游植物和浮游动物输送到不同海区(Chelton et al, 2011; Saito et al, 2014; Condie et al, 2016); 另一方面, 涡旋通常与中尺度和亚中尺度动力过程有关(McWilliams, 2016), 影响涡旋内营养盐和氧气等化学要素的分布, 从而改变浮游植物的生物量和分布(Labat et al, 2009; Hauss et al, 2016; Karstensen et al, 2017; Liu et al, 2022), 最终影响浮游动物的分布和群落结构。通常认为, 冷涡的上凸等密度面在涡中心引起上升流, 带来高营养, 有利于浮游植物生长; 暖涡的下凹等密度面在涡中心引起下降流, 将浮游植物带到深水中, 由于营养消耗过多且光照不足, 浮游植物消亡, 浮游动物生物量相应减少(Labat et al, 2009; Chang et al, 2017)。而在涡旋边缘, 锋面造成的次级环流补充了营养盐, 浮游动物生物量较多(Jiang et al, 2007; Labat et al, 2009; Liu et al, 2020)。浮游植物是决定浮游动物分布和生物量的决定性因素。因此一些观测发现在反气旋暖涡内, 由于叶绿素浓度高, 浮游动物生物量反而更高(Jiang et al, 2007; Saito et al, 2014)。此外, Xue等(2021)在黑潮延伸体海区的走航观测发现Sv与温度有关, 北部亲潮冷水区的生物量少, 而南部黑潮暖水区的生物量多, 且暖涡高温中心的生物量比涡边缘多, 这与早期Aoki等(1992)在该海区的观测结果一致。
本文分别针对暖涡边缘的水平强流和暖涡中心的高温对DVM的影响进行了分析, 首次提出涡边缘的强流减弱DVM强度, 而涡中心的高温减小DVM幅度。由于缺乏同步的生化要素观测, 上述结论尚无法揭示温度和强流影响DVM的途径(比如, 改变浮游植物的生物量和垂向分布)。因此, 研究涡旋对浮游动物的影响, 不仅要考虑海洋动力过程, 还要结合生化要素(特别是叶绿素浓度、初级生产力和物种类型)的综合影响, 从而厘清海洋动力过程影响浮游动物的途径。
3.3 展望全球海洋范围内, 有关浮游动物DVM的研究主要集中在入海口和近海(Yang et al, 2019; Liu et al, 2020), 大西洋(Labat et al, 2009; Ariza et al, 2016), 尤其是湾流区(Jiang et al, 2007; Eden et al, 2009), 地中海(van Haren, 2014; Potiris et al, 2018)和阿拉伯海(Luo et al, 2000; Ashjian et al, 2002), 而黑潮-亲潮混合区的相关观测和研究都很少。本文为该海区浮游动物的垂向分布和DVM研究提供了一定的参考, 为该海区生态模型的优化和改进以及渔业资源评估和预测奠定了基础(Lehodey et al, 2015)。当前, KEMS已经积累了多年的潜标观测的回声、流速和温盐数据, 可用于研究涡旋、锋面(Labat et al, 2009)甚至更长时间尺度的海洋动力过程和气候变化(Chiba et al, 2009, 2013)等对浮游动物的分布、群落结构和DVM的影响。然而, 对于研究浮游动物DVM在生态和生物地球化学循环中的作用, 还需要物理(温度、盐度、海流等)、化学(营养盐、溶解氧、碳等)和生物(浮游动物采样)的同步观测数据作为支撑。
4 结论通过分析黑潮-亲潮混合区深海潜标所搭载的ADCP得到的后向散射强度Sv, 本文发现: (1) 该海区上1 000 m存在两个浮游动物组成的强散射层, 分别位于200 m以浅的上层海洋和300~800 m之间的中层海洋, 具有显著的季节变化。上层浮游动物的生物量春秋多、夏冬少, 随叶绿素浓度的升高而增加; 其分布深度夏季最浅, 秋冬随混合层的加深逐渐向下扩展到200 m, 总体上分别受浮游植物生物量和垂向分布的影响。中层浮游动物的生物量春夏多、秋冬少, 随水温的降低而减少; 其分布深度春夏深、秋冬浅, 随水温的升高而加深; 总体上与水温的季节变化一致。(2) 400 m层的浮游动物存在显著的DVM行为, 表现为: 日落时向上迁移到表层, 日出时向下迁移回到白天的栖息层。年平均的DVM幅度、时长和速度分别200, 2 h和2.8 cm/s, 瞬时DVM速度可高达9.0 m/s。(3) DVM具有显著的多时间尺度变化。季节上, DVM强度春季最强、秋冬次之、夏季最弱, DVM幅度(速度)夏季最大、春季次之、秋冬小, 二者分别受浮游植物生物量和垂向分布的影响。DVM起止时间则受光照的调控, 与日落日出的季节变化同步。季节内, DVM强度的变化与温度和流速相关。DVM强度随温度的升高(降低)而增强(减弱), 同期相关系数整体为0.52, 局部高达0.8。DVM强度还受到水平强流的削弱作用, 在强流期间显著减弱。(4) 暖涡总体上抑制DVM, 但涡旋不同位置对其影响不同: 暖涡边缘的水平强流减弱DVM强度, 而暖涡中心的高温减小DVM幅度。
马超, 庄之栋, 刘勇, 等, 2018. 西北太平洋公海灯光敷网渔获组成及主要种类渔业生物学特性研究. 渔业研究, 40(2): 141-147 |
刘顺会, 孙松, 韩博平, 2008. 浮游动物昼夜垂直迁移机理的主要假说及其研究进展. 生态科学, 27(6): 515-521 DOI:10.3969/j.issn.1008-8873.2008.06.014 |
孙栋, 王春生, 2017. 深远海浮游动物生态学研究进展. 生态学报, 37(10): 3219-3231 |
李劳钰, 王辉武, 吕连港, 2012. 基于声学方法的南黄海浮游动物垂直迁移季节变化研究. 海洋科学, 36(9): 96-101 |
张武昌, 2000. 浮游动物的昼夜垂直迁移. 海洋科学, 24(11): 18-21 DOI:10.3969/j.issn.1000-3096.2000.11.008 |
陈子飞, 任强, 南峰, 等, 2021. 深海锚系潜标ADCP数据处理的质量控制研究. 海洋技术学报, 40(4): 54-61 |
陈钊, 吕连港, 杨光兵, 等, 2016. 基于船载ADCP和LADCP观测的南海声散射层. 海洋科学进展, 34(2): 240-249 DOI:10.3969/j.issn.1671-6647.2016.02.009 |
徐亚军, 赵亮, 原野, 2016. 基于声学仪器与粒径分析仪研究东海浮游动物昼夜垂直迁移过程. 海洋学报, 38(8): 123-130 |
靳少非, 唐峰华, 戴立峰, 等, 2014. 2010年北太平洋柔鱼渔场浮游动物多样性. 海洋环境科学, 33(4): 503-508 |
AOKI I, INAGAKI T, 1992. Acoustic observations of fish schools and scattering layers in a Kuroshio warm-core ring and its environs. Fisheries Oceanography, 1(2): 137-142 DOI:10.1111/j.1365-2419.1992.tb00032.x |
ARCHIBALD K M, SIEGEL D A, DONEY S C, 2019. Modeling the impact of zooplankton diel vertical migration on the carbon export flux of the biological pump. Global Biogeochemical Cycles, 33(2): 181-199 DOI:10.1029/2018GB005983 |
ARIZA A, LANDEIRA J M, ESCÁNEZ A, et al, 2016. Vertical distribution, composition and migratory patterns of acoustic scattering layers in the Canary Islands. Journal of Marine Systems, 157: 82-91 DOI:10.1016/j.jmarsys.2016.01.004 |
ASHJIAN C J, SMITH S L, FLAGG C N, et al, 2002. Distribution, annual cycle, and vertical migration of acoustically derived biomass in the Arabian Sea during 1994–1995. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 49(12): 2377-2402 DOI:10.1016/S0967-0645(02)00041-3 |
BEHRENFELD M J, GAUBE P, PENNA A D, et al, 2019. Global satellite-observed daily vertical migrations of ocean animals. Nature, 576(7786): 257-261 DOI:10.1038/s41586-019-1796-9 |
BENOIT-BIRD K J, LAWSON G L, 2016. Ecological insights from pelagic habitats acquired using active acoustic techniques. Annual Review of Marine Science, 8: 463-490 DOI:10.1146/annurev-marine-122414-034001 |
BIANCHI D, GALBRAITH E D, CAROZZA D A, et al, 2013. Intensification of open-ocean oxygen depletion by vertically migrating animals. Nature Geoscience, 6(7): 545-548 DOI:10.1038/ngeo1837 |
BRIERLEY A S, 2014. Diel vertical migration. Current Biology, 24(22): R1074-R1076 DOI:10.1016/j.cub.2014.08.054 |
CHANG Y L, MIYAZAWA Y, OEY L Y, et al, 2017. The formation processes of phytoplankton growth and decline in mesoscale eddies in the western North Pacific Ocean. Journal of Geophysical Research: Oceans, 122(5): 4444-4455 DOI:10.1002/2017JC012722 |
CHELTON D B, SCHLAX M G, SAMELSON R M, 2011. Global observations of nonlinear mesoscale eddies. Progress in Oceanography, 91(2): 167-216 DOI:10.1016/j.pocean.2011.01.002 |
CHIBA S, DI LORENZO E, DAVIS A, et al, 2013. Large-scale climate control of zooplankton transport and biogeography in the Kuroshio-Oyashio Extension region. Geophysical Research Letters, 40(19): 5182-5187 DOI:10.1002/grl.50999 |
CHIBA S, SUGISAKI H, NONAKA M, et al, 2009. Geographical shift of zooplankton communities and decadal dynamics of the Kuroshio-Oyashio currents in the western North Pacific. Global Change Biology, 15(7): 1846-1858 DOI:10.1111/j.1365-2486.2009.01890.x |
CONDIE S, CONDIE R, 2016. Retention of plankton within ocean eddies. Global Ecology and Biogeography, 25(10): 1264-1277 DOI:10.1111/geb.12485 |
DEEVEY G B, BROOKS A L, 1977. Copepods of the Sargasso sea off Bermuda: species composition, and vertical and seasonal distribution between the surface and 2000 M. Bulletin of Marine Science, 27(2): 256-291 |
DEINES K L, 1999. Backscatter estimation using Broadband acoustic Doppler current profilers [C] // Proceedings of the IEEE Sixth Working Conference on Current Measurement (Cat. No. 99CH36331). San Diego, CA, USA: IEEE: 249-253.
|
DONEY S C, STEINBERG D K, 2013. The ups and downs of ocean oxygen. Nature Geoscience, 6(7): 515-516 DOI:10.1038/ngeo1872 |
EDEN B R, STEINBERG D K, GOLDTHWAIT S A, et al, 2009. Zooplankton community structure in a cyclonic and mode-water eddy in the Sargasso Sea. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 56(10): 1757-1776 DOI:10.1016/j.dsr.2009.05.005 |
GOSTIAUX L, VAN HAREN H, 2010. Extracting meaningful information from uncalibrated backscattered echo intensity data. Journal of Atmospheric and Oceanic Technology, 27(5): 943-949 DOI:10.1175/2009JTECHO704.1 |
HAUSS H, CHRISTIANSEN S, SCHÜTTE F, et al, 2016. Dead zone or oasis in the open ocean? Zooplankton distribution and migration in low-oxygen modewater eddies. Biogeosciences, 13(6): 1977-1989 DOI:10.5194/bg-13-1977-2016 |
INOUE R, KITAMURA M, FUJIKI T, 2016. Diel vertical migration of zooplankton at the S1 biogeochemical mooring revealed from acoustic backscattering strength. Journal of Geophysical Research: Oceans, 121(2): 1031-1050 DOI:10.1002/2015JC011352 |
JIANG S N, DICKEY T D, STEINBERG D K, et al, 2007. Temporal variability of zooplankton biomass from ADCP backscatter time series data at the Bermuda Testbed Mooring site. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 54(4): 608-636 DOI:10.1016/j.dsr.2006.12.011 |
KARSTENSEN J, SCHÜTTE F, PIETRI A, et al, 2017. Upwelling and isolation in oxygen-depleted anticyclonic modewater eddies and implications for nitrate cycling. Biogeosciences, 14(8): 2167-2181 DOI:10.5194/bg-14-2167-2017 |
KOBARI T, MOKU M, TAKAHASHI K, 2008. Seasonal appearance of expatriated boreal copepods in the Oyashio–Kuroshio mixed region. ICES Journal of Marine Science, 65(3): 469-476 DOI:10.1093/icesjms/fsm194 |
LABAT J P, GASPARINI S, MOUSSEAU L, et al, 2009. Mesoscale distribution of zooplankton biomass in the northeast Atlantic Ocean determined with an Optical Plankton Counter: relationships with environmental structures. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 56(10): 1742-1756 DOI:10.1016/j.dsr.2009.05.013 |
LEHODEY P, CONCHON A, SENINA I, et al, 2015. Optimization of a micronekton model with acoustic data. ICES Journal of Marine Science, 72(5): 1399-1412 DOI:10.1093/icesjms/fsu233 |
LIU Y L, GUO J S, XUE Y H, et al, 2022. Seasonal variation in diel vertical migration of zooplankton and micronekton in the Andaman Sea observed by a moored ADCP. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 179: 103663 DOI:10.1016/j.dsr.2021.103663 |
LIU H J, ZHU M L, GUO S J, et al, 2020. Effects of an anticyclonic eddy on the distribution and community structure of zooplankton in the South China Sea northern slope. Journal of Marine Systems, 205: 103311 DOI:10.1016/j.jmarsys.2020.103311 |
LUO J G, ORTNER P B, FORCUCCI D, et al, 2000. Diel vertical migration of zooplankton and mesopelagic fish in the Arabian Sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 47(7/8): 1451-1473 |
MCWILLIAMS J C, 2016. Submesoscale currents in the ocean. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 472(2189): 20160117 DOI:10.1098/rspa.2016.0117 |
MULLISON J, 2017. Backscatter estimation using broadband acoustic Doppler current profilers-updated [C] // Proceedings of the ASCE Hydraulic Measurements & Experimental Methods Conference. Durham, NH, USA: 1-5.
|
POTIRIS E, FRANGOULIS C, KALAMPOKIS A, et al, 2018. Acoustic Doppler current profiler observations of migration patterns of zooplankton in the Cretan Sea. Ocean Science, 14(4): 783-800 DOI:10.5194/os-14-783-2018 |
QIU B, CHEN S, 2010. Eddy-mean flow interaction in the decadally modulating Kuroshio Extension system. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 57(13/14): 1098-1110 |
SAITO R, YAMAGUCHI A, YASUDA I, et al, 2014. Influences of mesoscale anticyclonic eddies on the zooplankton community south of the western Aleutian Islands during the summer of 2010. Journal of Plankton Research, 36(1): 117-128 DOI:10.1093/plankt/fbt087 |
SUGIMOTO S, AONO K, FUKUI S, 2017. Local atmospheric response to warm mesoscale ocean eddies in the Kuroshio-Oyashio Confluence region. Scientific Reports, 7(1): 11871 DOI:10.1038/s41598-017-12206-9 |
SUN S, TAO Z C, LI C L, et al, 2011. Spatial distribution and population structure of Euphausia pacifica in the Yellow Sea (2006–2007). Journal of Plankton Research, 33(6): 873-889 DOI:10.1093/plankt/fbq160 |
Teledyne R D, 2011. Acoustic Doppler current profiler principles of operation a practical primer [R]. San Diego: RD Instruments.
|
THYGESEN U H, PATTERSON T A, 2019. Oceanic diel vertical migrations arising from a predator-prey game. Theoretical Ecology, 12(1): 17-29 DOI:10.1007/s12080-018-0385-0 |
VAN HAREN H, 2014. Internal wave–zooplankton interactions in the Alboran Sea (W-Mediterranean). Journal of Plankton Research, 36(4): 1124-1134 DOI:10.1093/plankt/fbu031 |
XING X G, BOSS E, CHEN S L, et al, 2021. Seasonal and daily-scale photoacclimation modulating the phytoplankton chlorophyll-carbon coupling relationship in the mid-latitude northwest pacific. Journal of Geophysical Research: Oceans, 126(10): e2021JC017717 |
XUE M H, TONG J F, TIAN S Q, et al, 2021. Broadband characteristics of zooplankton sound scattering layer in the Kuroshio-Oyashio confluence region of the northwest Pacific Ocean in summer of 2019. Journal of Marine Science and Engineering, 9(9): 938 DOI:10.3390/jmse9090938 |
YAMAGUCHI A, MATSUNO K, ABE Y, et al, 2014. Seasonal changes in zooplankton abundance, biomass, size structure and dominant copepods in the Oyashio region analysed by an optical plankton counter. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 91: 115-124 DOI:10.1016/j.dsr.2014.06.003 |
YANG C H, XU D F, CHEN Z Z, et al, 2019. Diel vertical migration of zooplankton and micronekton on the northern slope of the South China Sea observed by a moored ADCP. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 167: 93-104 DOI:10.1016/j.dsr2.2019.04.012 |
ZHAO Z X, ALFORD M H, GIRTON J B, et al, 2016. Global observations of open-ocean mode-1 M2 internal tides. Journal of Physical Oceanography, 46(6): 1657-1684 |