 2022, Vol. 53
2022, Vol. 53

雷州半岛沿岸海域浮游桡足类的分布特征及其影响因素

http://dx.doi.org/10.11693/hyhz20210900227
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 彭宇航, 张才学, 孙省利, 郑金容, 刘耀谦, 姚昕. 2022.
- PENG Yu-Hang, ZHANG Cai-Xue, SUN Xing-Li, ZHENG Jin-Rong, LIU Yao-Qian, YAO Xin. 2022.
- 雷州半岛沿岸海域浮游桡足类的分布特征及其影响因素
- DISTRIBUTION CHARACTERISTICS OF PLANKTONIC COPEPODS AND ITS INFLUENCING FACTORS IN THE COASTAL WATERS OFF LEIZHOU PENINSULA
- 海洋与湖沼, 53(2): 375-383
- Oceanologia et Limnologia Sinica, 53(2): 375-383.
- http://dx.doi.org/10.11693/hyhz20210900227
文章历史
-
收稿日期:2021-09-30
收修改稿日期:2021-10-26
引用本文
彭宇航, 张才学, 孙省利, 郑金容, 刘耀谦, 姚昕. 2022. 雷州半岛沿岸海域浮游桡足类的分布特征及其影响因素[J]. 海洋与湖沼, 53(2): 375-383

PENG Yu-Hang, ZHANG Cai-Xue, SUN Xing-Li, ZHENG Jin-Rong, LIU Yao-Qian, YAO Xin. 2022. DISTRIBUTION CHARACTERISTICS OF PLANKTONIC COPEPODS AND ITS INFLUENCING FACTORS IN THE COASTAL WATERS OFF LEIZHOU PENINSULA[J]. Oceanologia et Limnologia Sinica, 53(2): 375-383.
雷州半岛沿岸海域浮游桡足类的分布特征及其影响因素
广东海洋大学化学与环境学院 广东湛江 524088
摘要:根据2017年10月和2018年1、5、7月对雷州半岛沿岸海域浮游动物四个季度的调查数据, 分析了浮游桡足类群落组成的季节变化及其影响因素。结果共鉴定出浮游桡足类80种, 其中哲水蚤目55种, 占总种类数的68.7%; 剑水蚤目21种, 占总种类数的26.2%; 猛水蚤目3种, 占总种类数的3.8%; 鱼虱目1种, 占总种类数的1.3%; 种类数冬春季最多, 各有45种, 夏季40种, 秋季最少仅29种; 各季节间共有种数在18~25种之间, 季节更替率变化范围为53.2%~67.9%。优势种有9种, 均隶属于哲水蚤目, 分别是红纺锤水蚤(Acartia erythraea)、太平洋纺锤水蚤(Acartia pacifica)、汤氏长足水蚤(Calanopia thompsoni)、微刺哲水蚤(Canthocalanus pauper)、瘦尾胸刺水蚤(Centropages tenuicermis)、小拟哲水蚤(Paracalanus parvus)、亚强次真哲水蚤(Subeucalanus subcrassus)、锥形宽水蚤(Temora turbinata)和真刺水蚤幼体(Eucharta larva), 无全年优势种, 季节更替明显。多样性指数变化范围在0.30~3.20之间, 平均为1.68; 均匀度变化范围为0.16~0.99, 平均为0.70。桡足类丰度变化范围为0.09~790.95 ind./m3, 平均为34.94 ind./m3, 平均值春季最高, 达60.92 ind./m3, 冬季次之, 为43.93 ind./m3, 夏季为22.44 ind./m3, 秋季最低, 仅为12.47 ind./m3。丰度的平面分布显示: 雷州半岛南部沿岸海域4季均出现明显密集区, 东部沿岸海域整体高于西部, 西南部的丰度最低, 与各区域的生物资源和环境因素密切相关; 相关分析显示, 秋季、冬季和春季桡足类丰度与水温和盐度均呈非显著负相关, 夏季与盐度呈极显著正相关, 冬季与叶绿素a含量呈显著正相关。
关键词:浮游桡足类 种类组成 季节变化 雷州半岛 琼州海峡
DISTRIBUTION CHARACTERISTICS OF PLANKTONIC COPEPODS AND ITS INFLUENCING FACTORS IN THE COASTAL WATERS OFF LEIZHOU PENINSULA
School of Chemistry and Environment, Guangdong Ocean University, Zhanjiang 524088, China
Abstract: Based on the four seasonal survey data of zooplankton in the coastal waters off Leizhou Peninsula in October 2017 and January, May, and July 2018, seasonal changes in composition of the planktonic copepod community and influencing factors were analyzed. Eighty species of planktonic copepods were identified, including 55 species of Calanoida, accounting for 68.7% of the total number of species, followed by 21 species of Cyclopoida (26.2%), 3 Harpacticoida (3.8%), and one Caligidae (1.3%). The largest number of species were found in winter and spring, with 45 species each, followed by summer (40 species), and autumn (29 species). The number of species in common among seasons was 18~25, with a seasonal replacement rate of 53.2%~67.9%. Nine dominant species were found, namely, Acartia erythraea, Acartia pacifica, Calanopia thompsoni, Canthocalanus pauper, Centropages tenuicermis, Paracalanus parvus, Subeucalanus subcrassus, Temora turbinata, and Eucharta larva. They all are Calanoida and varied seasonally with no one species occurred all year round in dominant species. The diversity index ranged 0.30~3.20, on average value of 1.68. The evenness index ranged 0.16~0.99, on average of 0.70. The abundance of copepods ranged 0.09~790.95 ind./m3 on average of 34.94 ind./m3, and the average was the highest in spring (60.92 ind./m3), followed by winter (43.93 ind./m3), summer (22.44 ind./m3), and autumn (12.47 ind./m3). In horizontal distribution, the annual copepods density was high in the southern coastal waters off Leizhou Peninsula, higher in the eastern region than in the western, and the lowest density in the southwestern sea. The horizontal distribution was closely related to the biological resources and environmental factors in each region. At last, the correlation analysis revealed that copepod abundance had no significant negative correlation with water temperature and salinity in autumn, winter, and spring, a very significant positive correlation with salinity in summer, and a significant positive correlation with chlorophyll a in winter.
Key words:
planktonic copepods species composition seasonal variation Leizhou Peninsula Qiongzhou Strait
海洋浮游桡足类隶属于节肢动物门、甲壳纲、桡足亚纲, 是海洋浮游动物中的一个重要组成部分, 种类多、数量大、分布广(郑重等, 1978)。桡足类一般摄食浮游植物, 同时本身也是许多经济鱼类特别是幼鱼的主要饵料, 是海洋生态系统中非常重要的次级生产者, 在能量和物质从初级生产者转移到更高的营养水平方面发挥着关键的生态作用(郑重等, 1982; Atkinson, 1996; 王荣等, 2002)。海洋浮游动物特别是桡足类, 其群落结构和时空分布格局与全球环境变化和渔业资源的盛衰有着密切的联系(Piontkovski et al, 2003; 张才学等, 2011), 在指示海流、水团、水域环境状况和探索鱼群、渔场等方面都有着广泛的研究。
雷州半岛处于中国大陆最南端, 属热带季风气候北缘, 沿岸海域同时受到南海和北部湾的影响, 具有复杂的潮波系统和水文特征, 半岛岸线漫长曲折, 拥有珊瑚礁、海草床、红树林、潟湖、基岩等多种海岸类型, 港湾众多。随着雷州半岛沿海经济的快速发展和城市化进程的加快, 人们对海岸带的开发利用强度不断加大, 给沿岸海域的生态环境带来了较大的影响, 开展半岛沿岸海域浮游动物的时空分布研究可揭示其对环境变化的生态响应。
有关雷州半岛沿岸海域的浮游桡足类研究已有报道(尹健强等, 2008; 龚玉艳等, 2015; 曾嘉维等, 2016), 但仅仅局限于某块小的区域, 对于整个半岛沿岸海域浮游桡足类的生态研究未见详细报道。本文基于雷州半岛海岸带的综合调查, 分析了半岛沿岸海域浮游桡足类的种类组成、优势种、数量分布等群落结构特点及其与环境因子的关系, 以期为半岛沿岸海域生态环境的保护, 维持海洋生物资源的可持续利用提供科学依据。
1 材料与方法 1.1 样品的采集与分析2017年10月(秋季)和2018年1月(冬季)、5月(春季)和7月(夏季)对雷州半岛沿岸海域进行了周年的季节综合调查, 共布设19条断面57个站位(图 1), 各站位水深3~18 m。为便于分析, 本文将调查海区整体划分为半岛东部、南部和西部沿岸海域, 并进一步细分为5个区域, 即: 半岛东北部(Ⅰ区: 1~12站位)、半岛东南部(Ⅱ区: 13~21站位), 半岛南部(Ⅲ区: 22~33站位)、半岛西南部(Ⅳ区: 34~45站位)和半岛西北部(Ⅴ区: 46~57站位)沿岸海域。
 |
| 图 1 雷州半岛沿岸海域采样站位 Fig. 1 Sampling stations in the coastal waters off Leizhou Peninsula |
浮游动物样品使用装有流量计的浅水I型浮游生物网(网目孔径0.505 mm, 网口内径50 cm, 网长145 cm)由底层至表层垂直拖网采集, 现场用5%的福尔马林溶液固定保存, 带回实验室在体视显微镜下鉴定计数。表层水温、盐度、pH值分别采用温度计、Orion130A盐度计和Orion3star pH计现场测定。每个采样点采集水样5 L, 用于氨氮、亚硝酸盐、硝酸盐、磷酸盐、硅酸盐和叶绿素a等指标的测定。样品的采集、保存和分析均参照《海洋调查规范》(GB/T 12763—2007), 《海洋监测规范》(GB 17378—2007)。
1.2 数据分析浮游桡足类的多样性和均匀度分别采用Shannon多样性指数(H′)和Pielou均匀度指数(J)进行计算, 公式如下:
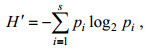 (1)
(1)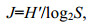 (2)
(2)式中, S为样品中总物种数, pi为第i种在全部样品中的比例(pi=ni/N), ni为第i种的个体数, N为所有种的个体总数。
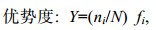 (3)
(3)式中, fi为第i种在采样站位中出现的频率, 以Y > 0.02作为优势种。
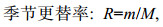 (4)
(4)式中, m为两个季节间不相同的物种数, M为两个季节总物种数。
浮游桡足类相关数据使用Excel 2016处理, 其丰度与环境因子的相关分析采用SPSS软件完成, 利用ArcGIS 10.2、Ocean Data View等软件绘制站位图以及桡足类丰度的平面分布图等。
2 结果 2.1 浮游桡足类种类组成本次调查共发现浮游桡足类80种(含12种幼体), 隶属4目21科35属, 其中哲水蚤目55种, 占总种类数的68.7%; 剑水蚤目21种, 占总种类数的26.2%; 猛水蚤目3种, 占总种类数的3.8%; 鱼虱目1种, 占总种类数的1.3%。种类最多的是大眼剑水蚤属(Corycaeus), 共8种, 其次是唇角水蚤属(Labidocera) (7种), 长腹剑水蚤属(Oithona)有6种, 胸刺水蚤属(Centropages)、角水蚤属(Pontella)、简角水蚤属(Pontellopsis)各4种, 纺锤水蚤属(Acartia)、隆哲水蚤属(Acrocalanus)、拟哲水蚤属(Paracalanus)、宽水蚤属(Temora)、歪水蚤属(Tortanus)各3种, 其他属种类仅有1种或2种。秋季鉴定出桡足类29种, 冬季和春季各45种, 夏季40种, 各季节间共有种数在18~25种之间, 季节更替率变化范围为53.2%~67.9% (表 1), 表明雷州半岛沿岸海域浮游桡足类种类季节更替较明显。
表 1 雷州半岛沿岸海域浮游桡足类季节更替率 Tab. 1 Seasonal replacement rate of planktonic copepods in the coastal waters off Leizhou Peninsula
| 指标 | 秋/冬 | 冬/春 | 春/夏 | 夏/秋 |
| 种数 | 29/45 | 45/45 | 45/40 | 40/29 |
| 共有种数 | 18 | 24 | 25 | 22 |
| 季节更替率 | 67.9% | 63.6% | 58.3% | 53.2% |
桡足类丰度以春季最高、冬季次之、夏季随之、秋季最低(图 2), 平面分布整体呈现半岛南部沿岸海域丰度高, 有明显的密集区, 其次是东部沿岸海域, 西部沿岸海域整体最低(图 3)。
 |
| 图 2 雷州半岛沿岸海域浮游桡足类丰度的季节变化 Fig. 2 Seasonal variation of zooplankton copepods abundance in the coastal waters off Leizhou Peninsula |
 |
| 图 3 雷州半岛沿岸海域浮游桡足类丰度的平面分布和季节变化 Fig. 3 Horizontal distribution and seasonal variation of planktonic copepods abundance in the coastal waters off Leizhou Peninsula |
秋季, 桡足类丰度变化范围在0.23~127.62 ind./m3, 平均为12.47 ind./m3, 其中24号站位丰度最高, 以亚强次真哲水蚤(Subeucalanus subcrassus)为主, 且占该站位总丰度的88.62%, 10号站位丰度最低, 半岛南部沿岸海域桡足类丰度较高, 其他海域桡足类丰度较低且分布较均匀。
冬季, 桡足类丰度变化范围在0.24~790.95 ind./m3, 平均为43.93 ind./m3, 最大值出现在14号站位, 即位于硇洲岛南部海域, 其丰度高达790.95 ind./m3, 最小值在39号站位, 除硇洲岛附近海域外, 在半岛南部和西北部沿岸海域也出现了丰度较密集区。
春季, 桡足类丰度分布不均匀, 变化范围为0.09~466.51 ind./m3, 平均为60.92 ind./m3, 最大值出现在27号站位, 最小值在38号站位, 在半岛南部沿岸海域形成高丰度密集区, 桡足类丰度普遍较高, 其次是东部海域, 西部海域整体较低。
夏季, 桡足类丰度变化范围为0.13~271.19 ind./m3, 平均为22.44 ind./m3, 位于半岛西南部离岸较远的36号站位丰度最高, 为271.19 ind./m3, 14号站位最低, 同样在半岛南部沿岸海域形成密集区, 东部沿岸海域出现小范围的较密集区, 其他海域桡足类丰度相对较低。
2.3 优势种及季节变化各季节出现的优势种共有9种(含1种幼体), 分别是红纺锤水蚤(Acartia erythraea)、太平洋纺锤水蚤(Acartia pacifica)、汤氏长足水蚤(Calanopia thompsoni)、微刺哲水蚤(Canthocalanus pauper)、瘦尾胸刺水蚤(Centropages tenuicermis)、小拟哲水蚤(Paracalanus parvus)、亚强次真哲水蚤(Subeucalanus subcrassus)、锥形宽水蚤(Temora turbinata)、真刺水蚤幼体(Eucharta larva), 均属于哲水蚤目。微刺哲水蚤(Canthocalanus pauper)和亚强次真哲水蚤(Subeucalanus subcrassus)除在春季不为优势种、锥形宽水蚤(Temora turbinata)除在秋季不为优势种外, 在其余季节均为优势种(表 2)。
表 2 雷州半岛沿岸海域浮游桡足类优势种的季节变化 Tab. 2 Seasonal variation of dominant species of planktonic copepods in the coastal waters off Leizhou Peninsula
| 季节 | 优势种类 | 平均丰度/(ind./m3) | 丰度占比/% | 出现频度 | 优势度 |
| 秋季 | 红纺锤水蚤Acartia erythraea | 0.80 | 6.42 | 0.58 | 0.04 |
| 汤氏长足水蚤Calanopia thompsoni | 1.86 | 14.93 | 0.54 | 0.08 | |
| 微刺哲水蚤Canthocalanus pauper | 0.66 | 5.30 | 0.42 | 0.02 | |
| 亚强次真哲水蚤Subeucalanus subcrassus | 7.54 | 60.46 | 0.81 | 0.49 | |
| 冬季 | 太平洋纺锤水蚤Acartia pacifica | 11.80 | 26.85 | 0.81 | 0.22 |
| 微刺哲水蚤Canthocalanus pauper | 1.47 | 3.35 | 0.65 | 0.02 | |
| 瘦尾胸刺水蚤Centropages tenuicermis | 14.58 | 33.18 | 0.96 | 0.32 | |
| 小拟哲水蚤Paracalanus parvus | 6.77 | 15.40 | 0.95 | 0.15 | |
| 亚强次真哲水蚤Subeucalanus subcrassus | 2.37 | 5.39 | 0.70 | 0.04 | |
| 锥形宽水蚤Temora turbinata | 1.79 | 4.07 | 0.72 | 0.03 | |
| 真刺水蚤幼体Eucharta larva | 2.17 | 4.93 | 0.72 | 0.04 | |
| 春季 | 太平洋纺锤水蚤Acartia pacifica | 6.33 | 10.39 | 0.82 | 0.09 |
| 瘦尾胸刺水蚤Centropages tenuicermis | 11.31 | 18.57 | 0.86 | 0.16 | |
| 锥形宽水蚤Temora turbinata | 35.60 | 58.44 | 0.95 | 0.55 | |
| 夏季 | 红纺锤水蚤Acartia erythraea | 1.79 | 7.96 | 0.60 | 0.05 |
| 汤氏长足水蚤Calanopia thompsoni | 1.74 | 7.75 | 0.35 | 0.03 | |
| 微刺哲水蚤Canthocalanus pauper | 5.70 | 25.42 | 0.61 | 0.16 | |
| 亚强次真哲水蚤Subeucalanus subcrassus | 2.41 | 10.75 | 0.74 | 0.08 | |
| 锥形宽水蚤Temora turbinata | 8.16 | 36.34 | 0.63 | 0.23 |
桡足类多样性指数(H′)秋季变化范围为0.30~2.52, 平均为1.30, 冬季变化范围为0.68~3.08, 平均为2.19, 春季在0.49~2.82之间, 平均为1.52, 夏季在0.47~3.20之间, 平均为1.72。春季和秋季的多样性指数较冬季和夏季低, 这是由于春季和秋季优势种较为突出, 如春季的优势种锥形宽水蚤(Temora turbinata)和秋季的优势种亚强次真哲水蚤(Subeucalanus subcrassus)的优势度分别高达0.55、0.49, 致使桡足类的多样性降低。均匀度(J)指数变化范围为0.16~0.99, 其中, 夏季最高(0.77), 其次是冬季(0.72), 秋季和春季分别为0.67、0.64 (图 4)。
 |
| 图 4 雷州半岛沿岸海域浮游桡足类多样性指数(H′)和均匀度指数(J)的季节变化 Fig. 4 Seasonal variation of diversity index(H′) and evenness index (J) in the coastal waters off Leizhou Peninsula |
将桡足类丰度与水温、盐度、叶绿素进行了相关性分析, 结果见表 3。秋季、冬季和春季桡足类丰度与水温和盐度均呈非显著负相关, 夏季与水温呈非显著正相关, 与盐度呈极显著正相关; 冬季桡足类丰度与叶绿素a含量呈显著正相关。各季节环境因子平面分布情况见图 5。
表 3 浮游桡足类丰度与环境因子的相关分析 Tab. 3 The correlation analysis of planktonic copepods abundance and environmental factors
| 环境因子 | 浮游桡足类丰度 | |||
| 秋季 | 冬季 | 春季 | 夏季 | |
| 水温 | –0.090 | –0.057 | –0.020 | 0.050 |
| 盐度 | –0.086 | –0.103 | –0.050 | 0.389** |
| 叶绿素a | –0.194 | 0.266* | 0.088 | –0.226 |
| 注: “*”表示显著水平(P < 0.05, 双尾), “**”表示极显著水平(P < 0.01, two-tail), n=57 | ||||
 |
| 图 5 雷州半岛沿岸海域环境因子的季节变化 Fig. 5 Seasonal variation of environmental factors in the coastal waters off Leizhou Peninsula |
本次调查共发现浮游桡足类80种(含幼体), 分别隶属于哲水蚤目、剑水蚤目、猛水蚤目和鱼虱目, 以哲水蚤居多。并可大致划分为4个生态类群, 种类大都是暖水种, 属于典型的热带、亚热带沿岸群落。其中, 适应偏低温低盐的近岸暖温性生态类群出现的种类较少, 典型的代表种是中华哲水蚤, 暖温性种类的出现可能与南下沿岸流的影响有关(陈柏云, 1985); 适应偏高温低盐的近岸暖水性生态类群则是调查海区的主体类群, 其种类多、数量大且分布范围广, 优势种中大部分属于该类群; 外海暖水性生态类群在雷州半岛沿岸海域也有出现, 说明该片海域受南海外海水的强烈影响(龚玉艳等, 2015); 广温广盐性生态类群由于适应的温度和盐度变化范围较大, 在半岛沿岸海域也有非常广泛的分布。所有优势种均属于哲水蚤目, 季节更替明显, 无全年优势种, 暖水种优势度普遍较高, 这表明雷州半岛沿岸海域浮游桡足类群落有较为明显的热带、亚热带区系特征。
优势种的季节变化体现了半岛沿岸海域的环境变化特征, 调查海区秋季、冬季、春季、夏季平均水温分别为30.0、18.7、27.8、29.1 ℃, 对应盐度为29.44、31.65、30.61、32.27。秋季优势度最大的种类是亚强次真哲水蚤, 平均丰度为7.54 ind./m3, 该种可适应偏高温的环境(张才学等, 2014), 耐高温是其成为优势种的重要原因之一。适应低温环境的瘦尾胸刺水蚤(黄加祺等, 1986)在冬季成为第一优势种, 平均丰度达14.58 ind./m3, 春季随着气温回升, 其丰度有所降低。锥形宽水蚤属偏高温低盐的暖水性种类(张才学等, 2014), 在4个季节均有出现, 冬季由于温度低, 优势度并不明显, 随着春季温度的升高, 其优势度达到最大, 平均丰度为35.60 ind./m3, 夏季和秋季持续高温, 丰度逐渐下降。微刺哲水蚤也属偏高温低盐种, 在夏季成为仅次于锥形宽水蚤的第二优势种, 秋季和冬季优势度低, 春季虽有出现, 但未形成优势种, 可能是冬季温度低, 该种几近消失, 在向春季过渡中难以形成优势。
将本次调查结果与毗邻海域或海湾的历史资料进行了比较(表 4), 本调查浮游桡足类的平均丰度和种数与邻近海域存在差异, 可能与调查时间、调查范围、站位设置不同有关。湛江湾(龚玉艳等, 2015)浮游桡足类丰度高, 可能是因为湛江市区经济发达, 排海物质多并携带有大量的营养盐, 有利于浮游植物的生长繁殖, 为桡足类提供了充足的食物(张才学等, 2011)。调查时间和调查范围的不同是造成与北部湾(曾嘉维等, 2016)之间差异的主要原因, 但平均丰度值处于同一个量级。优势种方面, 历史中出现的强额孔雀哲水蚤、细长腹剑水蚤、驼背隆哲水蚤、简长腹剑水蚤和刺尾纺锤水蚤等在此次调查中并不是优势种, 而出现了新的优势种如红纺锤水蚤、汤氏长足水蚤。
表 4 雷州半岛沿岸海域桡足类群落结构与附近海湾的比较 Tab. 4 Comparison of copepods community structure in the coastal waters off Leizhou Peninsula and nearby bays
| 海域及调查时间 | 采样网具 | 桡足类种数 | 平均丰度/(ind./m3) | 优势种 | 参考文献 |
| 雷州半岛灯楼角珊瑚礁海区(2004.08) | Ⅰ型 | 45 | — | 强额孔雀哲水蚤, 细长腹剑水蚤, 驼背隆哲水蚤, 简长腹剑水蚤等 | 尹健强等, 2008 |
| 湛江湾(2010.10~2011.09) | Ⅰ型 | 30 | 177.84 | 太平洋纺锤水蚤, 瘦尾胸刺水蚤, 刺尾纺锤水蚤等 | 龚玉艳等, 2015 |
| 北部湾(2014.02, 2014.08) | Ⅰ型 | 124 | 59.91 | 异尾宽水蚤, 精致真刺水蚤, 亚强真哲水蚤, 锥形宽水蚤等 | 曾嘉维等, 2016 |
| 雷州半岛沿岸海域(2017.10~2018.07) | Ⅰ型 | 80 | 34.94 | 太平洋纺锤水蚤, 瘦尾胸刺水蚤, 亚强次真哲水蚤, 锥形宽水蚤, 微刺哲水蚤等 | 本次调查 |
雷州半岛海岸带跨度大, 沿岸海域面积广阔, 受大陆径流、入海河流、南下沿岸流以及南海高温高盐水的影响较大, 海水环境因子复杂多变, 将直接影响到浮游桡足类的时空分布。由于缺乏发达的游泳器官, 浮游桡足类一般随波逐流的在海中漂浮, 因此, 海流是影响桡足类地理分布的主要因子(郑重等, 1978), 温度和盐度在不同程度上也会对其分布产生影响(Huntley et al, 1992; Kleppel et al, 1996; Abramova et al, 2005)。作为浮游动物的重要饵料, 浮游植物的丰欠对桡足类丰度的变化也起着重要作用(左涛等, 2004; 孙洁等, 2019)。
调查海区秋季和夏季的平均水温均高于29.0 ℃, 过高的水温不利于桡足类的生存, 使其进入消退期, 某些种类还可能向外海深层水迁移(孙柔鑫等, 2014), 当温度达28 ℃时, 中华哲水蚤濒临死亡, 在50%的存活率情况下, 瘦尾胸刺水蚤温度上限为28.6 ℃ (黄加祺等, 1986; 张武昌等, 2000)。冬季, 水温迅速下降, 其变化范围在17.5~20.6 ℃之间, 较低温的环境有利于一些桡足类的生长繁殖, 特别是适应低温的太平洋纺锤水蚤和瘦尾胸刺水蚤等(黄加祺等, 1986; 高亚辉等, 1999), 在硇洲岛南部海域出现高丰度区, 提高了桡足类丰度的整体水平; 春季, 随着气温回升, 冬季优势种太平洋纺锤水蚤和瘦尾胸刺水蚤丰度有所降低, 但仍处于优势水平, 锥形宽水蚤数量在该季也达到最高峰; 然而随着夏季的到来, 水温持续升高, 桡足类数量又出现减少的趋势。
调查海区四季平均盐度表现为夏季(32.27) > 冬季(31.65) > 春季(30.61) > 秋季(29.44), 各季节间盐度相差不大, 主要是因为研究区域常年受南海高盐海水的调控, 盐度保持相对稳定, 可能对桡足类丰度影响并不明显。
半岛东部沿岸海域浮游桡足类丰度整体较高。冬季, 东南部沿岸海域浮游桡足类在硇洲岛附近出现高丰度, 主要由太平洋纺锤水蚤和瘦尾胸刺水蚤贡献, 由于温度低, 同时浮游植物繁盛, 桡足类食物来源充足, 非常适合适应低温环境的太平洋纺锤水蚤和瘦尾胸刺水蚤的生长繁殖(黄加祺等, 1986; 高亚辉等, 1999), 并形成优势种。其他季节桡足类丰度较低, 主要是因为东南部地区经济欠发达, 沿岸海域受人类生产活动干扰较小, 再加上不规则半日潮促进了沿岸水体与外海水的交换, 使得该片海域水体高盐、寡营养盐(张才学等, 2014), 浮游植物存量低, 进而间接导致浮游桡足类丰度偏低。东北部沿岸海域浮游桡足类丰度在春季最高, 其他三季也出现了较密集区, 该片海域邻近湛江市区, 受工农业生产和城市生活污水影响较大, 入海物质携带大量营养盐(张才学等, 2011), 有利于浮游植物的生长繁殖, 进而会提高浮游桡足类的丰度。
半岛南部沿岸海域浮游桡足类平均丰度最高, 4个季节均出现明显密集区。由于游泳能力较弱, 桡足类一般“随波逐流”(郑重等, 1978), 琼州海峡是粤西海区与北部湾海区进行水体交换的通道, 其潮汐自东向西由不规则半日潮逐渐变为全日潮, 海流强劲(柯佩辉, 1983; Shi et al, 2002), 因此是导致桡足类丰度四季都高的主要原因。桡足类丰度的季节变化与水温的季节性变化密切相关(张才学等, 2011), 琼州海峡桡足类丰度春季最高, 冬季和夏季次之, 秋季最低, 呈单峰型趋势。桡足类最适生长温度在20 ℃左右(林元烧等, 1984; 黄加祺等, 1986; 孙柔鑫等, 2014), 夏秋两季海水温度过高可能是造成该海区桡足类丰度相对于春季和冬季较低的一个重要原因。
半岛西部沿岸海域浮游桡足类丰度整体最低。其中, 西南部沿岸海域拥有大面积的海草床、珊瑚礁和白蝶贝自然保护区, 受人类活动干扰小, 渔业资源丰富(廖宝林等, 2011), 鱼类会大量摄食浮游桡足类, 同时, 该片海域的大型海藻和海草吸收营养盐与浮游植物形成竞争关系, 以及养殖贝类对浮游植物的滤食(张才学等, 2012; 钟超等, 2019), 均会对浮游植物造成不利影响, 桡足类的食物来源减少, 进而丰度较低。西北部沿岸海域浮游桡足类丰度稍高于西南部, 可能与西北部沿岸存在着大面积的养殖基地和红树林生态系统提供的营养物质有关(张才学等, 2014)。
4 结论(1) 雷州半岛沿岸海域浮游桡足类共鉴定出80种(含12种幼体), 隶属4目21科35属, 优势种有9种, 其群落结构较复杂, 属于典型的热带、亚热带沿岸群落。
(2) 浮游桡足类种数季节变化较明显, 冬季和春季各45种, 夏季40种, 秋季种类最少(29种)。多样性指数(H′)全年变化范围在0.30~3.20, 平均为1.68; 均匀度(J)指数全年变化范围在0.16~0.99, 平均为0.70。
(3) 浮游桡足类丰度变化范围在0.09~ 790.95 ind./m3, 平均为34.94 ind./m3, 季节变化呈现春季(60.92 ind./m3) > 冬季(43.93 ind./m3) > 夏季(22.44 ind./m3) > 秋季(12.47 ind./m3), 温度的季节变化对浮游桡足类种数和丰度有着重要影响。丰度的平面分布显示, 半岛南部沿岸海域四季均有明显密集区, 东部沿岸海域整体高于西部, 并且整体有近岸向远岸减小的趋势, 与各区域的生物资源和环境因素密切相关。
(4) 秋季、冬季和春季桡足类丰度与水温和盐度均呈非显著负相关, 夏季与盐度呈极显著正相关, 冬季与叶绿素a含量呈显著正相关。
参考文献
王荣, 张鸿雁, 王克, 等, 2002. 小型桡足类在海洋生态系统中的功能作用. 海洋与湖沼, 33(5): 453-460 DOI:10.3321/j.issn:0029-814X.2002.05.001 |
尹健强, 黄晖, 黄良民, 等, 2008. 雷州半岛灯楼角珊瑚礁海区夏季的浮游动物. 海洋与湖沼, 39(2): 131-138 DOI:10.3321/j.issn:0029-814X.2008.02.006 |
左涛, 王荣, 王克, 等, 2004. 夏季南黄海浮游动物的垂直分布与昼夜垂直移动. 生态学报, 24(3): 524-530 DOI:10.3321/j.issn:1000-0933.2004.03.019 |
孙洁, 孙省利, 刘耀谦, 等, 2019. 脱硫海水受纳海域浮游桡足类对环境因子的群落响应. 广东海洋大学学报, 39(5): 71-77 |
孙柔鑫, 王彦国, 连光山, 等, 2014. 海南岛西北沿岸海域浮游桡足类的分布及群落特征. 生物多样性, 22(3): 320-328 |
张才学, 陈慧妍, 孙省利, 等, 2012. 流沙湾海草床海域浮游植物的时空分布及其影响因素. 生态学报, 32(5): 1527-1537 |
张才学, 龚玉艳, 王学锋, 等, 2011. 湛江港湾浮游桡足类群落结构的季节变化和影响因素. 生态学报, 31(23): 7086-7096 |
张才学, 龚玉艳, 孙省利, 等, 2014. 2010年夏季雷州半岛沿岸海域浮游动物群落结构特征. 海洋学报, 36(4): 91-99 DOI:10.3969/j.issn.0253-4193.2014.04.008 |
张武昌, 王荣, 王克, 2000. 温度对中华哲水蚤代谢率的影响. 海洋科学, 24(2): 42-44 DOI:10.3969/j.issn.1000-3096.2000.02.014 |
陈柏云, 1985. 南海浮游桡足类的分布. 海洋通报, 4(4): 45-50 |
林元烧, 李松, 1984. 厦门港中华哲水蚤生活周期的初步研究. 厦门大学学报(自然科学版), 23(1): 111-117 DOI:10.3321/j.issn:0438-0479.1984.01.013 |
郑重, 李少菁, 李松, 等, 1982. 台湾海峡浮游桡足类的分布. 台湾海峡, 1(1): 69-79 |
郑重, 李松, 李少菁, 1978. 我国海洋浮游桡足类的种类组成和地理分布. 厦门大学学报(自然科学版), (2): 51-63 DOI:10.3321/j.issn:0438-0479.1978.02.004 |
柯佩辉, 1983. 琼州海峡的海流和水交换的初步分析. 热带海洋, 2(1): 42-46 |
钟超, 孙凯峰, 廖岩, 等, 2019. 广东流沙湾海草分布现状及其与不同养殖生境的关系. 海洋环境科学, 38(4): 521-527 |
高亚辉, 林波, 1999. 几种因素对太平洋纺锤水蚤摄食率的影响. 厦门大学学报(自然科学版), 38(5): 751-757 DOI:10.3321/j.issn:0438-0479.1999.05.021 |
黄加祺, 郑重, 1986. 温度和盐度对厦门港几种桡足类存活率的影响. 海洋与湖沼, 17(2): 161-167 |
龚玉艳, 张才学, 陈作志, 等, 2015. 湛江湾浮游动物群落结构特征及其周年变化. 海洋科学, 39(12): 46-55 DOI:10.11759/hykx20141027001 |
曾嘉维, 王学锋, 吕少梁, 等, 2016. 北部湾冬、夏季浮游桡足类群落的结构特征及环境适应性. 广东海洋大学学报, 36(6): 59-64 DOI:10.3969/j.issn.1673-9159.2016.06.010 |
廖宝林, 刘丽, 刘楚吾, 2011. 徐闻珊瑚礁的研究现状与前景展望. 广东海洋大学学报, 31(4): 91-96 DOI:10.3969/j.issn.1673-9159.2011.04.017 |
ABRAMOVA E, TUSCHLING K, 2005. A 12-year study of the seasonal and interannual dynamics of mesozooplankton in the Laptev Sea: significance of salinity regime and life cycle patterns. Global and Planetary Change, 48(1/3): 141-164 |
ATKINSON A, 1996. Subantarctic copepods in an oceanic, low chlorophyll environment: ciliate predation, food selectivity and impact on prey populations. Marine Ecology Progress Series, 130: 85-96 DOI:10.3354/meps130085 |
HUNTLEY M E, LOPEZ M D G, 1992. Temperature-dependent production of marine copepods: a global synthesis. The American Naturalist, 140(2): 201-242 DOI:10.1086/285410 |
KLEPPEL G S, DAVIS C S, CARTER K, 1996. Temperature and copepod growth in the sea: a comment on the temperature-dependent model of Huntley and Lopez. The American Naturalist, 148(2): 397-406 DOI:10.1086/285932 |
PIONTKOVSKI S A, LANDRY M R, 2003. Copepod species diversity and climate variability in the tropical Atlantic Ocean. Fisheries Oceanography, 12(4/5): 352-359 |
SHI M C, CHEN C S, XU Q C, et al, 2002. The role of Qiongzhou strait in the seasonal variation of the South China Sea circulation. Journal of Physical Oceanography, 32(1): 103-121 DOI:10.1175/1520-0485(2002)032<0103:TROQSI>2.0.CO;2 |