 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 马路阔, 陈全震, 程方平, 彭欣, 陈伟峰, 曾江宁, 史西志, 黄伟. 2022.
- MA Lu-Kuo, CHEN Quan-Zhen, CHENG Fang-Ping, PENG Xin, CHEN Wei-Feng, ZENG Jiang-Ning, SHI Xi-Zhi, HUANG Wei. 2022.
- 三门湾游泳动物群落结构与生物多样性研究
- ANALYSIS OF COMMUNITY STRUCTURE AND BIODIVERSITY OF NEKTON IN THE SANMEN BAY
- 海洋与湖沼, 53(2): 384-393
- Oceanologia et Limnologia Sinica, 53(2): 384-393.
- http://dx.doi.org/10.11693/hyhz20211000238
文章历史
-
收稿日期:2021-10-09
收修改稿日期:2021-11-16




2. 宁波大学海洋学院 浙江宁波 315211;
3. 浙江省海洋水产养殖研究所 浙江温州 325005
2. School of Marine Science, Ningbo University, Ningbo 315211, China;
3. Zhejiang Mariculture Research Institute, Wenzhou 325005, China
游泳动物是海洋生态系统中最重要的生态类群之一, 包括鱼类、软体动物中的头足类、甲壳动物中的虾类、蟹类以及海洋哺乳动物(李冠国等, 2011)。作为海洋渔业资源重点开发对象, 游动物物通常具有较高的经济价值, 其物种组成、资源动态及可持续利用问题直接关系到海洋捕捞业(唐启升等, 2001; 苏纪兰等, 2005)。此外, 游泳动物多为海洋食物网中的高级消费者, 可通过摄食低营养级浮游植物和浮游动物进而影响整个海洋生态系统(蒋日进等, 2019)。因此, 开展海洋游泳动物的研究不仅对研究海洋生态系统动力学具有重要理论价值, 同时可为渔业资源的可持续利用提供科学依据。
三门湾位于浙江省三门县以北、宁海县以东, 是浙江省面积仅次于杭州湾的第二大海湾。由于地处中亚热带季风气候区, 饵料丰富, 使其成为海洋生物栖息、繁衍的良好场所, 也是东海渔场的重要组成部分(梁静香等, 2020), 还是鳓(IIisha elongata)、黄鲫(Setipinna taty)、三疣梭子蟹(Portunus trituberculatus)等重要经济种类产卵场以及凤鲫(Coilia mystus)、曼氏无针乌贼(Sepiella maindroni)等的索饵场(周永东等, 2018)。近年来, 随沿岸社会经济的快速发展, 海岸围垦工程逐年增加、涉海工程大规模建设以及人为捕捞强度的增大造成游泳动物栖息地损失, 该海域的渔业资源相较20世纪50年代出现了较为明显的衰退, 主要表现为种类趋于小型化、低龄化、低值化(冀萌萌等, 2016; 聂振林等, 2018)。
鉴于渔业资源可持续利用的迫切需要, 关于游泳动物群落结构的研究逐渐成为热点问题(林龙山等, 2012)。游泳动物的群落特征、结构变化在一定程度上能反映生态系统物质转换和能量流动过程(单秀娟等, 2014)。以往研究中, 学者们已经对三门湾游泳动物种类组成、物种多样性、群落结构等开展了研究(蔡萌等, 2009; 李新等, 2014; 冀萌萌等, 2016; 聂振林等, 2018; 俞松立等, 2021)。然而, 关于该海域游泳动物的群落结构与环境因子相关性研究相对薄弱。本研究基于2019~2020年春、秋两季的调查数据, 分析了三门湾游泳动物的种类组成、资源密度、生物多样性指数等群落结构特征, 并探讨了群落结构与环境因子的关系, 旨在为了解该海域的渔业资源现状提供基础数据, 也为后续捕捞产业的调整及渔业资源管理政策的制定提供科学参考。
1 材料与方法 1.1 调查站位与区域根据《海洋渔业资源调查规范》(SC/T 9403—2012), 于2019年秋季(11月)和2020年春季(5月)在三门湾开展渔业资源调查。调查海域根据地理位置共分为湾内和湾外两个区域, 共20个站位(图 1), 调查范围为121.03°~121.91°E和28.71°~29.20°N。调查网具为单拖网[10.0 m (长)×4.0 m (宽)], 网目范围为20~50 mm, 网囊网目为20 mm。每个站位水平底拖1 h, 平均拖速3.0 kn。渔获物现场采集后, 冷冻保存, 待返回实验室对样品进行鉴定及分类, 并称量、计数和生物学测定。拖网调查同步使用便携式水质分析仪(美国YSI PRO 1020)现场测量水温、盐度、pH、溶解氧等环境参数。
 |
| 图 1 三门湾游泳动物调查站位 Fig. 1 Sampling sites of the fishery survey in the Sanmen Bay |
通过拖网扫海面积法计算游泳生物的相对资源密度D (重量: kg/km2; 尾数: 103 ind./km2), 公式为:
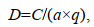 (1)
(1)式中: C为每小时取样面积内的渔获量重量(kg/h)或尾数(ind./h); a为每小时内的扫海面积(km2); q为捕捞系数, 取0.5。
1.2.2 优势种优势种的判定使用相对重要性指数(Index of Relative Importance, IRI):
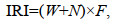 (2)
(2)式中, IRI为相对重要性指数; N为某一物种尾数占总尾数的百分比; W为该物种重量占总重量的百分比; F为某一物种出现的站数占调查总站数的百分比。将IRI > 500设定为优势种, 100≤IRI < 500为重要种, 10≤IRI < 100为常见种。
1.2.3 物种多样性利用Shannon-wiener指数H′, Pielou均匀度指数J′和Margalef丰富度指数D描述游泳动物的多样性特征。
Shannon-wiener多样性指数H′:
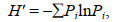 (3)
(3)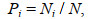 (4)
(4)Pielou均匀度指数J′:
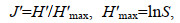 (5)
(5)Margalef丰富度指数D:
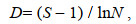 (6)
(6)式中, H′为物种多样性指数, H′max为最大物种多样性指数, Ni为第i物种的数量, N为所有物种的总数量, Pi为第i物种占所有物种的数量百分比, N表示总个体数, S为总物种数。采用独立样本t检验分析多样性参数的季节间差异(SPSS 20.0)。
1.2.4 群落结构相似性分析将游泳动物生物量通过lg(x+1)进行转换并构建不同站位间游泳动物物种组成的Bray-Curtis相似性系数矩阵, 应用聚类分析和非度量多维标度(non-metric multi-dimensional scaling, NMDS)将各站位划分为不同群组并以此为基础划分生态区系。胁迫系数(stress)可以用来衡量非度量多维标度分析法的可信度, 当stress≥0.2时, 排序结果不能正确解释群落结构的关系; 当0.1≤stress < 0.2时, 可用NMDS排序图表示, 且具有一定解释意义; 当0.05≤stress < 0.1时, 排序结果基本可信, 结果较好; 当stress < 0.05时, 排序结果具有很好的代表性。
1.2.5 冗余分析首先对各个季节优势种的资源密度进行去趋势对应分析(detrended correspondence analysis, DCA)分析, 根据DCA分析结果的轴梯度长度(lengths of gradient, LGA)选择分析方法, 若LGA值小于3, 则选用冗余分析(redundancy analysis, RDA); 若介于3和4之间, 则RDA和典范对应分析(canonical correlation analysis, CCA)均可; 若大于4, 则选用CCA分析方法分析春、秋两季的环境理化因子和资源密度的对应关系。根据DCA分析结果, 本文采用线性模型的RDA分析。
调查站位图、资源密度平面分布图以及多样性指数分布图使用Arcgis10.8软件绘制; 组平均聚类法分析和非度量多维标度分析使用PRIMER 6.0软件进行; RDA分析使用Canoco 4.5软件进行。
2 结果 2.1 种类组成和优势种本次调查春、秋季共鉴定出游泳动物83种, 隶属于16目40科61属(见附录, 另列文件)。其中鱼类11目23科33属42种, 占总捕获物种数的50.60%; 甲壳类2目14科24属38种, 占总捕获物种的45.78%; 头足类3目3科3属3种, 占总捕获物种数的3.61%。
春季渔获物共52种, 隶属于12目29科43属, 包括头足类2种、甲壳类24种、鱼类26种。秋季渔获物共68种, 隶属于14目33科50属, 包括头足类2种、甲壳类33种、鱼类33种。
春季各物种IRI范围为0.08~9 260.92, 优势种为三疣梭子蟹(Portunus trituberculatus)、口虾蛄(Oratosquilla orarotia)、中华栉孔虾虎鱼(Ctenotrypauchen chinensis)、龙头鱼(Harpadon nehereus) 4种。秋季各物种IRI范围为0.10~7 897.97, 优势种为脊尾白虾(Exopalaemon carinicauda)、三疣梭子蟹、日本蟳(Charybdis japonica)、口虾蛄、窝纹网虾蛄(Dictyosquilla foveolata)、龙头鱼、棘头梅童鱼(Collichthys lucidus) 7种(见图 2)。在春、秋两季中, 口虾蛄、龙头鱼和三疣梭子蟹均为优势种。
 |
| 图 2 三门湾春、秋季优势种IRI指数 Fig. 2 Index of relative importance of dominant species in spring and autumn in the Sanmen Bay 注: 灰色为春季优势种, 黑色为秋季优势种 |
春、秋季各生物类群的个体数占比和生物量占比如表 1, 其中甲壳类的个体数占比和生物量占比在春、秋季均为最高。
| 生物类群 | 春季 | 秋季 | |||
| 个体数占比 | 生物量占比 | 个体数占比 | 生物量占比 | ||
| 头足类 | 0.27% | 1.61% | 0.04% | 0.05% | |
| 甲壳类 | 65.96% | 68.03% | 63.12% | 66.43% | |
| 鱼类 | 33.76% | 30.36% | 36.84% | 33.52% | |
春、秋季各生物类群以及总体的个体数资源密度以及生物量资源密度见表 2和图 3, 春、秋季均是甲壳类的资源密度较高。
| 种类 | 春季 | 秋季 | |||
| 个体数资源密度/(ind./km2) | 生物量资源密度/(kg/km2) | 个体数资源密度/(ind./km2) | 生物量资源密度/(kg/km2) | ||
| 头足类 | 0.068×103 | 5.69 | 0.007×103 | 0.12 | |
| 甲壳类 | 9.67× 103 | 223.97 | 18.96×103 | 259.54 | |
| 鱼类 | 9.18×103 | 117.07 | 10.25×103 | 106.09 | |
| 总 | 23.45×103 | 356.73 | 28.40×103 | 384.10 | |
 |
| 图 3 三门湾游泳动物资源密度时空分布 Fig. 3 Spatio-temporal distribution in resource density of the nekton in the Sanmen Bay 注: a、c分别为春、秋季个体数资源密度(×103 ind./km2); b、d为春、秋季生物量资源密度(kg/km2) |
三门湾海域春、秋季游泳动物群落结构变化明显, 相似性检验(ANOSIM)非参数多元分析结果显示春、秋季群落差异极显著(R=0.505, P < 0.01)。如图 4, NMDS排序的协强系数为0.16, 可知该海域NMDS排序图具有一定解释意义。在50.15%的相似性水平上可以将春、秋季站位分为六个组群, 其中秋季所有站位为一组, 春季18号站位为一组, 1号站位为一组, 2号站位为一组, 18号站位为一组, 19和20号站位为一组, 剩余春季站位为一组。
 |
| 图 4 三门湾游泳动物群落结构的组平均聚类分析 Fig. 4 Cluster analysis of group average of community structure of nekton in the Sanmen Bay 注: SP1~SP20为春季1~20号站位, AU1~AU20为秋季1~20号站位 |
基于渔获物重量组成, 春季各站位的多样性指数H′和均匀度指数J′的均值分别为1.31和0.33。秋季各站位的多样性指数H′和均匀度指数J′均值分别为1.57和0.37。基于渔获物个体数组成, 春季各站位的多样性指数H′和均匀度指数J′的均值分别为1.69和0.43。秋季各站位的多样性指数H′和均匀度指数J′均值分别为2.13和0.50 (图 5)。
 |
| 图 5 三门湾海域游泳动物生物量多样性指数H′(a)、个体数多样性指数H′(b)、生物量均匀度指数J′(c)、个体数均匀度指数J′(d) Fig. 5 Seasonal variation of biodiversity index H′ (a), individual diversity index H′ (b), biomass uniformity index J′ (c), and individual uniformity index J′ (d) of nekton community in the Sanmen Bay 注: 柱上不同字母a、b表示春、秋季的多样性指数和均匀度指数存在显著差异(P < 0.05) |
秋季的多样性指数、均匀度指数均显著高于春季(P < 0.05)。
2.4.2 地理分布差异以各站基于生物重量的H′和J′为例, 春季, 15号站位的H′最高(2.29), 18号站位的H′最低(0.18)。全部站位中12、13、15、16和19站位的生物多样性水平相对较高, 主要分布于湾外区域。秋季, 17号站位的H′最高(1.97), 4号站位的H′最低(0.77)。秋季生物多样性水平相对较高的站位有6、7、9、17、18和19号站位, 在湾内湾外均有分布(图 6)。基于生物个体数的H′和J′与基于生物量的H′和J′的空间分布趋势基本一致。
 |
| 图 6 三门湾游泳动物群落的H′、J′和D的时空分布 Fig. 6 Spatio-temporal distribution of the H′, J′, and D of nekton community in the Sanmen Bay 注: a, b: 基于个体数和生物量的Shannon-wiener多样性指数; b, d: 基于个体数和生物量的Pielou均匀度指数; e: 丰富度指数 |
春、秋两季生物多样性指数最高的站位均位于湾外区域。
2.5 RDA分析春季筛选出8个主要物种与5个环境因子与进行RDA分析(图 7a), 前两轴5个环境因子累积解释了游泳动物39.09%的物种变量(表 3), 第一轴(特征值0.228 6)和第二轴(特征值0.162 3)的物种-环境相关系数分别为0.770 6和0.730 1, 表明这5种环境因子与这8种游泳动物的相关性较好。秋季筛选出8个主要物种与5个环境因子与进行RDA分析(图 7b), 前两轴5个环境因子累积解释了游泳动物23.88%的物种变量(表 3), 第一轴(特征值0.148 5)和第二轴(特征值0.090 4)的物种-环境相关系数分别为0.869 7和0.823 7, 表明这5种环境因子与这8种游泳动物的相关性较好。
 |
| 图 7 三门湾春、秋季游泳动物优势种与环境因子的RDA排序 Fig. 7 The seasonal distribution RDA sort of dominant species of nekton and environmental factors in spring and autumn in the Sanmen Bay 注: a. 春季分析结果; b. 秋季分析结果; Sal: 盐度; Depth: 深度; DO: 溶解氧; Temp: 温度 |
| 季节 | 特征值 | 物种-环境相关系数 | 解释变量 | |||||
| 轴一 | 轴二 | 轴一 | 轴二 | 轴一 | 轴二 | |||
| 春季 | 0.228 6 | 0.162 3 | 0.770 6 | 0.730 1 | 22.86 | 39.09 | ||
| 秋季 | 0.148 5 | 0.090 4 | 0.869 7 | 0.823 7 | 14.85 | 23.88 | ||
本研究春、秋季共鉴定出游泳动物83种, 这一结果高于冀萌萌等(2016)和聂振林等(2018)在2014~2015年的调查结果。春季鉴定出游泳动物52种, 秋季鉴定出游泳动物68种, 其中鱼类的种类最多为42种。甲壳类的生物量高于鱼类和头足类。这可能与以下两个原因有关: 首先, 鱼类多为季节性洄游物种, 而甲壳类多为短距离洄游或定居物种, 并且三门湾海域存在三疣梭子蟹的产卵场(周永东等, 2018); 其次, 蟹类生长周期短, 性成熟早, 繁殖力强, 在一个产卵期间可以多次产卵(俞存根等, 2003)。
本研究中, 口虾蛄、三疣梭子蟹以及龙头鱼在春、秋两季均为优势种。这个结果与温台渔场、玉环海域的调查结果相似(孙鹏, 2018; 韩晓凤, 2020)。口虾蛄为广温广盐性种类, 因此口虾蛄不仅在三门湾海域, 在整个浙江沿海资源量都十分丰富(梅文骧等, 1996)。三疣梭子蟹也是广温广盐性种类, 它不仅是浙江沿海重要的经济蟹类, 更是中国海捕经济蟹类当中最重要的品种。2020年浙江省三疣梭子蟹的产量超过了15万t (农业农村部渔业渔政管理局等, 2021)。龙头鱼是广温广盐的暖水性近海小型底层鱼类, 性凶猛, 由于过度捕捞, 近年来与龙头鱼竞争食物的高营养级游泳动物逐渐减少, 导致龙头鱼资源数量不断增加(罗海舟等, 2012)。本调查中春、秋两季上述三种优势种在总渔获量的数量占比均大于60%, 表明目前三门湾海域生态系统的能量流动和生态位主要由少数广温广盐的种类控制, 这在舟山群岛东侧海域以及披山海域的生态位分析中也有体现(刘坤等, 2021)。
3.2 生物多样性以及资源密度的时空差异生物多样性指数反映了群落结构的空间异质性, 也能体现群落结构的稳定性(韩晓凤, 2020)。三门湾海域秋季的多样性指数、均匀度指数均高于春季, 此结果与台湾海峡以及东海北部海域的调查结果类似, 这种季节性差异可能与环境条件和游泳动物的生态习性有关(宋普庆等, 2012; 唐政, 2020)。三门湾为强潮海湾, 潮差大、水体交换较快, 带来了丰富的营养盐和生物饵料, 秋季会吸引多种游泳动物洄游至该海域进行索饵(中国海湾志编纂委员会, 1992)。春、秋季多样性指数较高的站位大多位于湾外区域, 主要原因是这些站位远离陆地, 受人类活动影响较小以及湾外区域环境复杂程度更高, 能满足更多游泳动物对不同生境的需求。
三门湾秋季游泳动物的个体数资源密度和生物量资源密度明显高于春季, 这与黄海和东海北部海域以及舟山群岛外海域的调查结果吻合(唐政, 2020; 于南京等, 2021)。春季为大多数鱼类的产卵季节, 一般而言东海越冬鱼群在3月之后离开越冬场, 5月到达产卵场进行产卵, 鱼卵尚未孵化或幼鱼体型过小不易被网具捕获(严利平等, 2019)。秋季为大多数鱼类的索饵育肥季节, 许多鱼类发育为成鱼, 体长、体重等较春季都有所增加, 因此秋季的资源密度高于春季。如表 4, 与历史调查资料比对发现, 本调查的生物量资源密度高于2014年、2015年及2018年结果(冀萌萌等, 2016; 俞松立等, 2021)。其原因可能与伏季休渔制度的实行有关, 表明近年来三门湾海域渔业管理部门实施的资源养护措施取得了明显成效。
| 资源密度 | 调查时间(年.月) | 参考文献 | |
| 鱼类 | 游泳动物 | ||
| 117.07; 9.18×103 | 356.73; 23.45×103 | 2020.04 | 本调查 |
| 106.09; 10.25×103 | 384.10; 28.40×103 | 2019.11 | |
| 286.61; 42.68×103 | 2014.11 | 冀萌萌等, 2016 | |
| 314.44; 34.11×103 | 2015.05 | ||
| 136.40; 17.40×103 | 2018.04 | 俞松立等, 2021 | |
| 152.50; 15.3×103 | 2018.10 | ||
| 注: 资源密度单位分别表示为: 生物量资源密度kg/km2; 个体数资源密度ind./km2 | |||
从空间分布上看, 游泳动物个体数资源密度和生物量资源密度较高的站位主要位于湾内, 其中6号站位的生物量资源密度在春、秋季均为最高。原因可能是6号站位于田湾岛附近, 一方面岛屿附近流态复杂, 营养物质丰富, 适宜多种鱼类生存, 另一方面岛屿之间特殊的海底地形、底质和水流环境多样化满足了不同鱼类的生活要求, 使大多数鱼类有合适的栖息地(郭朋军等, 2020)。此外, 春季部分鱼类进行生殖洄游, 由湾外向湾内进行洄游, 导致湾内资源密度较高(夏陆军等, 2016)。与邻近海域的比较中, 三门湾海域的游泳动物资源密度处于中间水平, 小于闽东近海、东山湾、舟山群岛外海域和瓯江口, 但高于杭州湾(表 5)。其原因可能是不同海湾的开发利用程度不同, 人类活动对渔业资源造成的影响不一(林龙山等, 2012; 刘金虎等, 2020; 于南京等, 2021)。
| 资源密度 | 调查海域 | 参考文献 | |
| 鱼类 | 游泳动物 | ||
| 春: 210.50; 8.08×103 | 舟山群岛外海域 | 于南京等, 2021 | |
| 秋: 829.06; 165.94×103 | |||
| 2015秋: 10.85; 4.92×103 | 杭州湾 | 刘惠等, 2019 | |
| 2017秋: 47.12; 10.06×103 | |||
| 春: 311.0; 257.5×103 | 365.8; 304.6×103 | 瓯江口 | 胡成业等, 2015 |
| 春: 325.57; 3.4×105 | 闽江口 | 黄良敏, 2011 | |
| 秋: 942.38, 9.9×105 | |||
| 春: 756.6; 7.1×104 | 春: 1013.7; 10.2×104 | 闽东近海 | 刘金虎等, 2020 |
| 秋: 565.7; 4.0×104 | 秋: 899.0; 6.6×104 | ||
| 春: 471.53, 4.1×104 | 九龙江口 | 杜建国等, 2012 | |
| 秋: 143.97, 0.4×104 | |||
| 春: 290.66; 8×103 | 春: 366.66; 9×103 | 东山湾 | 林龙山等, 2012 |
| 秋: 309.60; 6×103 | 秋: 390.27; 11×103 | ||
| 春: 117.07; 9.18×103 | 春: 356.73; 23.45×103 | 三门湾 | 本调查 |
| 秋: 106.09; 10.25×103 | 秋: 384.10; 28.40×103 | ||
| 注: 资源密度单位分别表示为: 生物量资源密度kg/km2; 个体数资源密度ind./km2 | |||
冗余分析表明, 三门湾海域游泳动物群落结构受到温度、盐度、深度以及溶解氧等环境因子的主导影响。游泳动物栖息于不同水层, 因此受到不同水深梯度的影响(蒋日进等, 2019)。其群落结构在水深梯度变化不大的情况下对温度存在较大依赖性(刘勇等, 2021)。溶解氧是水生生物生存的必要条件之一, 对海洋生物的生存和新陈代谢至关重要(Ren et al, 2019)。根据渔业水质标准的要求, 水体中的溶解氧一昼夜有16个小时处于大于5 mg/L的水平即可, 本研究中春季的溶解氧虽然高于秋季, 但是春、秋季的溶解氧均较高, 对游泳动物的群落结构无影响(周莹, 2016)。
三门湾海域地形复杂, 岛屿众多, 受潮汐、湾外水团和江浙沿岸流的共同影响, 湾内外温度、盐度分布模式复杂(中国海湾志编纂委员会, 1992), 进而影响到游泳动物的分布格局。根据NMDS分析结果, 在50.15%相似性水平上可以将春、秋季站位分为六个组群。其中春季站位间存在群落结构差异, 春、秋季间存在群落结构差异。春季站位间群落结构存在差异的原因是春季1号站和2号站位于湾内区域, 靠近大陆, 18、19和20号站位于湾外区域, 靠近外海, 湾外海水盐度明显高于湾内区域。盐度会影响游泳动物的生理活动, 如代谢及渗透压调节等, 进而影响游泳动物群落结构。因此湾内区域分布有较多的斑
三门湾海域春、秋季共有的优势种包括口虾蛄、三疣梭子蟹等地方性种类。秋季该海域的平均水温为19.06 ℃, 相较于春季的17.98 ℃更接近口虾蛄以及三疣梭子蟹的适温范围, 这些物种秋季的资源量明显高于春季。由于秋季温度较高, 窝纹网虾蛄以及棘头梅童鱼在秋季也是优势种, 温度条件可以促进鱼类的脑垂体分泌促性腺激素, 调节鱼类的产卵行为, 同时浮游生物在温度较高的情况下繁殖更快, 为游泳动物提供更多食物(黄良敏等, 2013)。因此温度是影响三门湾游泳动物春、秋季群落结构差异的重要因素。
4 结论(1) 本研究共鉴定出游泳动物83种, 甲壳类是三门湾海域主要的游泳动物类群, 游泳动物的优势种存在一定的季节更替现象; 春、秋季游泳动物群落结构存在差异, 可能与春、秋季水温、盐度差异有关。
(2) 与往年三门湾游泳动物调查结果相比, 资源量有明显提高, 表明近年来环三门湾地区管理部门开展的渔业资源养护和生态环境保护措施已取得初步成效。
(3) 本研究中未能采集叶绿素、浮游植物、浮游动物等指标, 因此其他环境和生物要素对三门湾游泳动物群落结构的影响分析仍有待进一步深入。
致谢 自然资源部第二海洋研究所刘清河、于翔、朱根海、武芳竹和浙江省海洋水产养殖研究所叶深在实验过程中协助采样以及物种鉴定, 谨致谢忱。电子附件材料:
于南京, 俞存根, 许永久, 等, 2021. 舟山群岛外海域春秋季鱼类群落结构及生物多样性. 水产学报, 45(8): 1374-1383 |
中国海湾志编纂委员会, 1992. 中国海湾志第五分册: 上海市和浙江省北部海湾. 北京: 海洋出版社, 303
|
刘坤, 俞存根, 许永久, 等, 2021. 舟山群岛东侧海域春秋季主要甲壳类物种的空间生态位分析. 浙江大学学报(理学版), 48(4): 450-460, 480 |
刘金虎, 李毅, 曹亮, 等, 2020. 闽东近海游泳动物群落组成、资源量与生物多样性的时空格局研究. 海洋与湖沼, 51(4): 931-942 |
刘勇, 程家骅, 贾世广, 2021. 东海及黄海南部带鱼的海底水温分布特征及分析方法的优化. 水产学报, 45(6): 871-886 |
刘惠, 俞存根, 郑基, 等, 2019. 杭州湾灰鳖洋海域秋季主要游泳动物生态位与种间联结. 生态学报, 39(11): 3828-3836 |
农业农村部渔业渔政管理局, 全国水产技术推广总站, 中国水产学会, 2021. 2021中国渔业统计年鉴. 北京: 中国农业出版社
|
孙鹏, 2018. 玉环东部及附近海域游泳动物群落特征及多样性研究[D]. 舟山: 浙江海洋大学: 37-38.
|
严利平, 刘尊雷, 金艳, 等, 2019. 东海区延长伏季休渔期渔业资源增殖效果分析. 海洋渔业, 41(5): 513-519 |
苏纪兰, 唐启升, 2005. 我国海洋生态系统基础研究的发展——国际趋势和国内需求. 地球科学进展, 20(2): 139-143 DOI:10.3321/j.issn:1001-8166.2005.02.001 |
杜建国, 刘正华, 余兴光, 等, 2012. 九龙江口鱼类多样性和营养级分析. 热带海洋学报, 31(6): 76-82 DOI:10.3969/j.issn.1009-5470.2012.06.012 |
李冠国, 范振刚, 2011. 海洋生态学. 2版. 北京: 高等教育出版社, 83
|
李新, 周青松, 俞存根, 等, 2014. 浙江三门湾春季鱼类种类组成及多样性研究. 浙江海洋学院学报(自然科学版), 33(6): 488-494 DOI:10.3969/j.issn.1008-830X.2014.06.002 |
宋普庆, 张静, 林龙山, 等, 2012. 台湾海峡游泳动物种类组成及其多样性. 生物多样性, 20(1): 32-40 |
林龙山, 赵贵根, 李渊, 等, 2012. 东山湾及其邻近海域游泳动物多样性的分析. 水生生物学报, 36(6): 1119-1127 |
罗海舟, 张华东, 李鹏飞, 等, 2012. 东海区龙头鱼数量分布与渔业生物学现状分析. 浙江海洋学院学报(自然科学版), 31(3): 202-205, 233 DOI:10.3969/j.issn.1008-830X.2012.03.003 |
周永东, 李圣法, 2018. 东海区主要经济种类三场一通道及保护区图集. 北京: 海洋出版社
|
周莹, 2016. 水生生物对水体溶解氧日变化规律影响[D]. 沈阳: 沈阳师范大学: 30-31.
|
单秀娟, 陈云龙, 戴芳群, 等, 2014. 黄海中南部不同断面鱼类群落结构及其多样性. 生态学报, 34(2): 377-389 |
胡成业, 徐衡, 水柏年, 等, 2015. 温州瓯飞滩邻近海域春季游泳动物群落结构及多样性. 南方水产科学, 11(3): 7-15 DOI:10.3969/j.issn.2095-0780.2015.03.002 |
俞存根, 宋海棠, 姚光展, 等, 2003. 浙江近海蟹类资源合理利用研究. 海洋渔业, 25(3): 136-141 DOI:10.3969/j.issn.1004-2490.2003.03.008 |
俞松立, 王咏雪, 韩晓凤, 等, 2021. 三门湾海域渔业生物群落结构及营养级变化特征. 大连海洋大学学报, 36(3): 478-487 |
聂振林, 王咏雪, 胡成业, 等, 2018. 三门湾春秋季鱼类群落特征及其与环境因子的关系. 水产学报, 42(9): 1390-1398 |
夏陆军, 周青松, 俞存根, 等, 2016. 乐清湾口海域春秋季鱼类群落多样性研究. 渔业现代化, 43(2): 68-75 DOI:10.3969/j.issn.1007-9580.2016.02.013 |
郭朋军, 江新琴, 俞存根, 等, 2020. 舟山沿岸渔场春秋季鱼类群落结构特征分析. 渔业科学进展, 41(4): 1-11 |
唐启升, 苏纪兰, 2001. 海洋生态系统动力学研究与海洋生物资源可持续利用. 地球科学进展, 16(1): 5-11 DOI:10.3321/j.issn:1001-8166.2001.01.002 |
唐政, 2020. 黄海和东海北部海域游泳动物群落结构和多样性时空分布研究[D]. 上海: 上海海洋大学: 14-15.
|
黄良敏, 2011. 闽江口和九龙江口及其邻近海域渔业资源现状与鱼类多样性[D]. 青岛: 中国海洋大学: 23-24.
|
黄良敏, 张会军, 张雅芝, 等, 2013. 入海河口鱼类生物与水环境关系的研究现状与进展. 海洋湖沼通报, (1): 61-68 |
梅文骧, 王春琳, 张义浩, 等, 1996. 浙江沿海虾蛄生物学及其开发利用研究报告. 浙江水产学院学报, 15(1): 1-8 |
梁静香, 周永东, 王忠明, 等, 2020. 三门湾大型底栖动物群落结构及其与环境因子的关系. 应用生态学报, 31(9): 3187-3193 |
蒋日进, 张琳琳, 徐开达, 等, 2019. 浙江中南部近岸海域游泳动物功能群特征与多样性. 生物多样性, 27(12): 1330-1338 DOI:10.17520/biods.2019281 |
韩晓凤, 2020. 温台渔场产卵场保护区及附近海域游泳动物群落结构及多样性研究[D]. 舟山: 浙江海洋大学: 19-20.
|
蔡萌, 徐兆礼, 2009. 浙江三门湾冬夏季鱼类种类组成和数量变化. 上海海洋大学学报, 18(2): 198-205 |
冀萌萌, 田阔, 胡成业, 等, 2016. 三门湾春、秋季游泳动物群落组成及物种多样性研究. 水生态学杂志, 37(2): 42-48 |
REN Q Q, XIAN W W, LIU C L, et al, 2019. Spring-time nektonic invertebrate assemblages of and adjacent to the Yangtze Estuary. Estuarine, Coastal and Shelf Science, 227: 106338 DOI:10.1016/j.ecss.2019.106338 |