 2022, Vol. 53
2022, Vol. 53

两种孔径网具对大洋微(小)型浮游植物调查结果的影响——以西南印度洋海域第34次大洋春季航次为例

http://dx.doi.org/10.11693/hyhz20211200316
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 郭卓然, 冷天泽, 陆斗定, 戴鑫烽. 2022.
- GUO Zhuo-Ran, LENG Tian-Ze, LU Dou-Ding, DAI Xin-Feng. 2022.
- 两种孔径网具对大洋微(小)型浮游植物调查结果的影响——以西南印度洋海域第34次大洋春季航次为例
- IMPACT OF PLANKTON NETS WITH DIFFERENT MESHSIZES ON OCEAN PHYTOPLANKTON SURVEY: TAKE THE 34TH SPRING CRUISE IN THE SOUTHWEST INDIA OCEAN AS AN EXAMPLE
- 海洋与湖沼, 53(3): 637-644
- Oceanologia et Limnologia Sinica, 53(3): 637-644.
- http://dx.doi.org/10.11693/hyhz20211200316
文章历史
-
收稿日期:2021-12-09
收修改稿日期:2022-01-17
引用本文
郭卓然, 冷天泽, 陆斗定, 戴鑫烽. 2022. 两种孔径网具对大洋微(小)型浮游植物调查结果的影响——以西南印度洋海域第34次大洋春季航次为例[J]. 海洋与湖沼, 53(3): 637-644

GUO Zhuo-Ran, LENG Tian-Ze, LU Dou-Ding, DAI Xin-Feng. 2022. IMPACT OF PLANKTON NETS WITH DIFFERENT MESHSIZES ON OCEAN PHYTOPLANKTON SURVEY: TAKE THE 34TH SPRING CRUISE IN THE SOUTHWEST INDIA OCEAN AS AN EXAMPLE[J]. Oceanologia et Limnologia Sinica, 53(3): 637-644.
两种孔径网具对大洋微(小)型浮游植物调查结果的影响——以西南印度洋海域第34次大洋春季航次为例
自然资源部海洋生态系统动力学重点实验室 自然资源部第二海洋研究所 浙江杭州 310012
摘要:浮游植物拖网是海洋浮游植物调查的重要工具, 不同网具可以导致不同的调查结果。为研究两种孔径网具对大洋浮游植物调查的影响, 于2015年4~5月在西南印度洋分别用筛绢孔径为20和77 μm的两种浮游生物网在多个站位进行浮游植物采样。对采集的样品进行浮游植物鉴定, 分析比较了两者浮游植物的物种数、细胞丰度、物种丰富度、多样性指数、均匀度指数和优势种。结果表明, 在相同采样站位, 筛绢孔径为20 μm网采样品鉴定得到的平均物种数、平均细胞丰度和平均物种丰富度分别是筛绢孔径为77 μm网采样品的2.7、17和2倍, 呈显著差异。但两种网采样品的平均多样性指数和平均均匀度指数都无显著差异。此外, 筛绢孔径为20 μm网采样品中有9种优势种, 而筛绢孔径为77 μm网采样品中有11种优势种, 其中共同的优势种有6种。以上结果说明在寡营养海域使用20 μm孔径的网具可以获得更为全面的浮游植物信息。
关键词:孔径 网具 浮游植物 大洋调查
IMPACT OF PLANKTON NETS WITH DIFFERENT MESHSIZES ON OCEAN PHYTOPLANKTON SURVEY: TAKE THE 34TH SPRING CRUISE IN THE SOUTHWEST INDIA OCEAN AS AN EXAMPLE
Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China
Abstract: Phytoplankton trawl net is a effect tool in the survey of marine phytoplankton. Different results could be obtained by using different nets. To investigate the impact of different mesh sizes of plankton trawl nets on ocean phytoplankton survey, phytoplankton samples were collected by trawl nets in two different mesh sizes of 20 μm and 77 μm in the Southwest Indian Ocean from April to May, 2015. The phytoplankton was identified in taxonomy and the species number, cell abundance, species richness, diversity index, evenness index, and dominant species of the samples collected with the nets of two mesh sizes were compared. Results show that the mean number of species, mean cell abundance, and mean species richness of the 20-μm-mesh samples were significantly greater than those of the 77-μm-mesh samples for about 2.7 times, 17 times, and 2 times, respectively. However, no significant difference was shown in mean diversity index and evenness index between the two mesh-size samples. In addition, the number of dominant species was 9 and 11 for 20-μm-mesh and 77-μm-mesh samples, respectively, among them 6 were common. Therefore, using 20-μm-mesh trawl net is recommended for phytoplankton survey in oligotrophic waters as more phytoplankton information can be obtained.
Key words:
mesh size plankton trawl net phytoplankton ocean survey
浮游植物是海洋生态系统中最主要的初级生产者, 其合成的有机物是维持海洋生态系统运行的物质基础。在营养盐较为丰富的近岸海区, 微型(Nano)和小型(Net)浮游植物往往是初级生产者的主要构成部分(Agawin et al, 2000; Fu et al, 2009; Kostadinov et al, 2010); 而在寡营养的大洋海区中微微型浮游植物(Pico)占主导地位, 对总叶绿素a浓度的贡献率高达70%~90% (Odate, 1997; Savidge et al, 1999; Hong et al, 2012)。
浮游植物拖网是开展海洋浮游植物调查的重要工具, 但各国常用的浮游植物网具在孔径上存在差异。例如国际上多采用网筛绢孔径在20 μm左右的网具进行浮游植物群落调查(孙军等, 2003), 而中国的大洋浮游植物调查主要依据《海洋调查规范第6部分: 海洋生物调查GB/T 12763. 6—2007》, 30 m以浅的水域采用浅水Ⅲ型浮游生物网(网口直径37 cm, 网长1.4 m, 筛绢孔径77 μm); 在30 m以深的水域垂直或分段采集小型浮游植物时选择小型浮游生物网(网口直径37 cm, 网长2.8 m, 绢筛孔径77 μm) (中华人民共和国国家质量监督检验检疫总局等, 2008)。这里没有区分近岸和大洋的网具孔径, 但是近岸海域与大洋环境之间的浮游植物群落特征往往存在较大差异, 比如浮游植物细胞丰度近岸高而远洋低。因此, 采用孔径相同的浮游生物网是否能如实反映不同区域的浮游植物群落结构特征存在疑问。
印度洋是世界第三大洋, 印度洋西南海域的主要环流系统为由南印度洋副热带涡流(IOSG)的分支沿非洲东海岸向南运输而形成的厄加勒斯流(Read et al, 2000; Machu et al, 2005; Povinec et al, 2011)。研究表明, 该海区浮游植物生长主要受到硝酸盐的限制, 且浮游植物粒径以微微型(Pico)为主(洪丽莎等, 2012)。为了研究不同孔径网具对调查结果的影响, 本文在西南印度洋采用筛绢孔径分别为20和77 μm的两种浮游生物网在多个相同站位进行浮游植物分别采样, 通过对比分析, 为大洋浮游植物调查中浮游生物网规格的选择与改进提供科学依据。
1 材料与方法 1.1 研究区域2015年4~5月, 搭载中国“大洋一号”调查船第34航次在西南印度洋海域进行浮游植物采样调查, 共设置5个采样站位, 站位信息如表 1和图 1所示。其中, 34IV-SWIR-S017至34IV-SWIR-S019和34IV-SWIR-S039四个站位的空间位置较为接近, 而34IV-SWIR-S006站位的空间位置较其他站位相距甚远。
表 1 2015年西南印度洋第34航次浮游植物采样站位信息 Tab. 1 Sampling station of the 34th cruise in the Southwest Indian Ocean in 2015
| 站位 | 采样日期 | 经度/(°E) | 纬度/(°S) | 水深/m | 网采深度/m | 网具选择 | |
| 20 μm | 77 μm | ||||||
| 34IV-SWIR-S017 | 2015/4/4 | 50.4129 | 37.6426 | 1823 | 200 | Y | Y |
| 34IV-SWIR-S018 | 2015/4/5 | 50.2524 | 37.6282 | 1937 | 200 | Y | Y |
| 34IV-SWIR-S019 | 2015/4/7 | 50.5010 | 37.7809 | 1680 | 200 | Y | Y |
| 34IV-SWIR-S039 | 2015/4/14 | 50.0708 | 37.0031 | 3054 | 200 | Y | Y |
| 34V-IB-S006 | 2015/5/11 | 81.4002 | 22.4747 | 4888 | 200 | N | Y |
| 注: Y表示采样, N表示未采样 | |||||||
 |
| 图 1 2015年西南印度洋第34航次浮游植物采样站位图 Fig. 1 Sampling stations for phytoplankton in the 34th cruise in the Southwest Indian Ocean in 2015 |
为了采集调查0~200 m水层浮游植物, 34IV- SWIR-S017、34IV-SWIR-S018、34IV-SWIR-S019与34IV-SWIR-S039四个站位均采用两种筛绢孔径不同的浮游生物网(网口直径37 cm, 网长2.8 m, 网筛绢孔径分别为77和20 μm)进行分别采样, 34IV-IB-S006站位因条件限制只用孔径为77 μm的浮游生物网进行采样, 共采得网采浮游植物样品9个。采集的样品立即加入中性福尔马林固定保存(福尔马林体积分数终浓度为4%), 并带回实验室分析。
样品鉴定前, 先将样品轻轻充分摇匀, 使用移液枪吸取1 mL样品移置于计数框(Sedgewick Rafter, PYSER-SGI, UK)内, 盖上盖玻片, 利用倒置光学显微镜(Olympus CKX41, Japan), 在100或400倍的视野下进行观察, 进行物种和细胞数量的计数。
1.3 数据处理对所得数据进行香农-威纳指数(H′)、优势度指数(Y)、物种丰富度(D)以及均匀度指数(J′)分析, 计算公式如下:
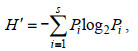 (1)
(1)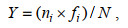 (2)
(2)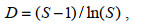 (3)
(3)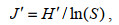 (4)
(4)式中: ni指第i个物种的个体数; N指样品中的总个体数; fi是该物种在样品种的出现频率, 以优势度(Y)≥0.01的物种判定为优势种。S为样品中的浮游植物种类总数; Pi为第i个物种的个体数(ni)与总体数(N)的比值。
物种丰富度(D)通过PRIMER7计算, 分析筛网孔径与采得样品的联系。另因34IV-SWIR-S006站只采集筛绢孔径为77 μm的网采样品, 除独立站位分析外, 不纳入对比统计分析。
2 结果 2.1 浮游植物种类组成2015年4~5月, 从西南印度洋采集取得的样品中共鉴定出浮游植物192种, 隶属于4个门, 包括硅藻门(Bacillariophyta) 106种, 甲藻门(Pyrrophyta) 79种, 金藻门(Chrysophyta) 4种, 蓝藻门(Cyanophyta) 3种, 所占总物种比例分别为55%、41%、2%和2% (图 2b)。硅藻门中最多的为角毛藻属(Chaetoceros spp.) 28种, 根管藻属(Rhizosolenia spp.) 12种, 圆筛藻属(Coscinodiscus spp.) 11种; 甲藻门中最多的为角藻属(Ceratium spp.) 28种, 鳍藻(Dinophysis spp.)和多甲藻属(Peridinium spp.)各8种。金藻门中包括等刺硅鞭藻属(Dictyocha spp.) 3种和异刺硅鞭藻属(Distephanus spp.) 1种; 蓝藻门中包括颤藻属(Oscillatoria spp.)、席藻属(Phormidinum spp.)和束毛藻属(Trichodesmium spp.)各1种。
 |
| 图 2 2015年西南印度洋第34航次各站位物种组成 Fig. 2 The phytoplankton community structure of each site in the 34th cruise in the Southwest Indian Ocean in 2015 注: a. 各站位采用20和77 μm孔径网具采得浮游植物物种组成; b. 采用20和77 μm孔径网具采得浮游植物物种数平均值及总站位物种组成; A、B表明采用两种孔径网具采得物种数平均值差异极显著 |
除S006站位以外其他各个站位不同网具的采样结果对比显示, 筛绢孔径为20 μm的情况下鉴定得到的物种数明显高于筛绢孔径为77 μm的情况(图 2)。筛绢孔径为20 μm的浮游生物网采集到浮游植物共计163种, 隶属于4个门, 包括硅藻门90种, 甲藻门67种, 金藻门4种和蓝藻门2种, 物种数平均值为71, 四个门物种占比各为54.58%、41.20%、2.82%和1.41%; 筛绢孔径为77 μm的浮游生物网采集到浮游植物共计67种, 隶属于3个门, 包括硅藻门43种, 甲藻门22种和蓝藻门2种, 与前者相比未检出金藻门物种, 物种数平均值为26, 而三个门物种占比各为67.31%、27.88%和4.81%。采用绢筛孔径为20 μm的浮游生物网采集得到的平均物种数约为采用孔径为77 μm浮游生物网的2.7倍。
2.2 浮游植物丰度通过样品分析得知, 浮游植物细胞丰度最高值出现在34IV-SWIR-S017站位, 为62 491 cells/m3; 细胞丰度最低值出现在34IV-SWIR-S018站位, 为13 213 cells/m3。硅藻、甲藻和蓝藻的最高丰度均出现在34IV-SWIR-S017站位, 而金藻的最高丰度出现在34IV-SWIR-S039站位。34IV-IB-S006站位的细胞丰度为1 093 cells/m3, 只鉴定出硅藻和甲藻, 分别占总细胞丰度的86.55%和13.45% (图 3a)。将所有数据相加后得到总站位细胞丰度, 其中硅藻占81.88%, 甲藻占16.62%, 金藻占1.01%, 蓝藻占0.48% (图 3b)。
 |
| 图 3 2015年西南印度洋第34航次各站位细胞丰度 Fig. 3 The cell abundance at each stations of the 34th cruise in the Southwest Indian Ocean in 2015 注: a. 各站位采用20和77 μm孔径网具采得的浮游植物细胞丰度; b. 采用20和77 μm孔径网具采得的浮游植物细胞丰度平均值及总站位细胞丰度; ab表明采用两种孔径网具采得细胞丰度平均值差异显著 |
采用筛绢孔径为20 μm浮游生物网采集到的四个站位的细胞丰度平均值为37 026 cells/m3, 其中硅藻占81.56%, 甲藻占17.05%, 金藻占1.08%, 蓝藻占0.31%。而采用筛绢孔径为77 μm浮游生物网采集到除34IV-IB-S006站位以外的四个站位的细胞丰度平均值为2 183 cells/m3, 其中硅藻占86.76%, 甲藻占9.76%, 蓝藻占3.48%。前者的平均值约为后者的17倍, 即采用20 μm孔径网具可以采到的浮游植物细胞数量更多, 且物种比例与总站位相近。
2.3 浮游植物物种多样性指数浮游植物的群落特征通过香农-威纳指数(H′)、物种数(S)、均匀度指数(J′)以及物种丰富度(D)这四种参数表征。其中, 使用20 μm孔径的网具采到的样品物种数和细胞丰度都显著高于采用77 μm孔径的网具(图 4b, 4d), 而两种不同孔径的网采样品之间, 香农-威纳多样性与均匀度指数结果无显著差异(图 4a, 4c)。因此, 采用较细密绢筛制作的网具可以采到更多的物种。但使用孔径不同的网具, 得到的物种多样性指数无显著差异。
 |
| 图 4 2015年西南印度洋第34航次各站位多样性指数 Fig. 4 The diversity indices of the 34th cruise in the Southwest Indian Ocean in 2015 注: a、b、c和d分别表示各站位采用两种不同孔径网具采样获得的香农-威纳指数H′、物种数S、均匀度指数J′和物种丰富度D差异; A、B表明两者差异极显著; a、a表明两者差异不显著 |
西南印度洋调查海域浮游植物优势物种共计14种, 其中筛绢孔径为20 μm的网采样品与筛绢孔径为77 μm的网采样品分析结果中分别得出的优势种也存在差异, 为9种和11种。两种网采样品的共同优势种包括条纹小环藻(Cyclotella striata var. striata)、圆筛藻sp.1 (Coscinodiscus sp.1)、细弱海链藻(Thalassiosira subtilis)、长海毛藻(Thalassiothrix longissima)、夜光藻(Noctiluca scintillans)、美丽漂流藻(Planktoniella formosa) 6种。除上述6种外, 在孔径为20 μm的网采样品中优势种还包括五角角藻(Ceratium pentagonum)、双角角管藻(Cerataulina bicornis)和棘角毛藻(Chaetoceros imbricatus); 在孔径为77 μm的网采样品中优势种还包括翼根管藻(Rhizosolenia alata f. alata)、大西洋角毛藻(Chaetoceros atlanticus var. atlanticus)、汉氏束毛藻(Trichodesmium hildebrandtii)、翼根管藻纤细变型(Rhizosolenia alata f. gracillima)、笔尖形根管藻(Rhizosolenia styliformis var. styliformis)。其中条纹小环藻的细胞丰度和优势度最高, 笔尖形根管藻的细胞丰度和优势度最低。各优势种的细胞丰度、所占比例以及优势度如表 2所示。
表 2 2015年西南印度洋第34航次调查的浮游植物优势种情况 Tab. 2 Dominant species of phytoplankton during the 34th cruise of the Southwest Indian Ocean in 2015
| 种名 | 20 μm孔径网筛 | 77 μm孔径网筛 | |||||
| 细胞丰度/(cells/m3) | 比例/% | 优势度 | 细胞丰度/(cells/m3) | 比例/% | 优势度 | ||
| 条纹小环藻 Cyclotella striata var. striata |
43 103 | 29.10 | 0.22 | 713 | 7.26 | 0.03 | |
| 圆筛藻sp.1 Coscinodiscus sp.1 |
14 277 | 9.64 | 0.10 | 340 | 3.46 | 0.01 | |
| 细弱海链藻 Thalassiosira subtilis |
12 870 | 8.69 | 0.04 | 2 486 | 25.31 | 0.20 | |
| 五角角藻 Ceratium pentagonum |
5 387 | 3.64 | 0.04 | — | — | — | |
| 长海毛藻 Thalassiothrix longissima |
3 960 | 2.67 | 0.03 | 887 | 9.03 | 0.09 | |
| 夜光藻 Noctiluca scintillans |
3 926 | 2.65 | 0.03 | 360 | 3.66 | 0.04 | |
| 双角角管藻 Cerataulina bicornis |
2 882 | 1.95 | 0.01 | — | — | — | |
| 棘角毛藻 Chaetoceros imbricatus |
4 187 | 2.83 | 0.01 | — | — | — | |
| 美丽漂流藻 Planktoniella formosa |
1 846 | 1.25 | 0.01 | 251 | 2.56 | 0.02 | |
| 翼根管藻 Rhizosolenia alata f. alata |
— | — | — | 681 | 6.93 | 0.07 | |
| 大西洋角毛藻 Chaetoceros atlanticus var. atlanticus |
— | — | — | 1 034 | 10.53 | 0.04 | |
| 汉氏束毛藻 Trichodesmium hildebrandtii |
— | — | — | 276 | 2.81 | 0.02 | |
| 翼根管藻纤细变型 Rhizosolenia alata f. gracillima |
— | — | — | 354 | 3.60 | 0.02 | |
| 笔尖形根管藻 Rhizosolenia styliformis var. styliformis |
— | — | — | 109 | 1.11 | 0.01 | |
| 注: 表格只显示优势种(优势指数≥0.01种为优势种), “—”表示无数据。 | |||||||
使用浮游生物网采集样品是研究浮游植物群落结构及生态演替的常用方法之一。相较于近岸海域的浮游植物采样调查, 大洋海域的调查更具有耗时长、成本高以及风险大等特点。为了在大洋调查中尽可能得到准确的浮游植物物种及细胞丰度数据, 根据调查区域特点和实验目的选择适合的浮游生物网十分必要。然而, 我国对于大洋调查中浮游植物群落调查的采样网具大多参考《海洋调查规范第6部分: 海洋生物调查GB/T 12763. 6—2007》。也有部分调查会根据自己需求采用特定网具, 所以在不同的大洋航次调查中, 浮游生物网的选择也各有不同。例如, 陈卓等(2018)在2016年秋季热带西太平洋调查中, 采用WP2型浮游生物网(网口直径为57 cm, 网长2.71 m, 网筛绢孔径为20 μm); 而郭术津等(2018)分别于2014年秋季和2016年冬季在热带西太平洋Y3海区和M2海山邻近海域调查时采用小型浮游生物网(网口直径37 cm, 网长2.7 m, 网筛绢孔径为76 μm)。因此, 本次调查对比两种孔径网具的采样结果, 有助于科研人员在大洋调查中根据需求来选择适用的网具。
本次调查研究显示筛绢孔径为20 μm网采样品鉴定得到的物种数和浮游植物细胞丰度都显著高于筛绢孔径为77 μm的网采样品, 说明采用较细密筛绢制作的网具可以截留到更细微的个体, 从而更加准确地分析调查海域的浮游植物群落现状。这一点也可从优势物种结果比较得到印证, 除6种共同优势种外, 20 μm孔径网采样品还包含了77 μm孔径网采样品中的其他5种优势物种。两者的差异主要来自对不同细胞大小的截留程度不同, 例如, 夜光藻的细胞个体较大, 范围在150~2 000 μm之间(胡翠林等, 2015), 因此可同时被20和77 μm孔径网具所采集到, 但五角角藻仅出现在采用20 μm孔径网具采集的样品当中。从物种方面看, 本次鉴定得到的金藻门物种包括六异刺硅鞭藻(Distephanus speculum)和小等刺硅鞭藻(Dictyocha fibula)及其变种共计4种, 金藻门物种的粒径主要在2~20 μm范围内(Brotas et al, 2013), 因此只在20 μm孔径网具采集的样品中发现。
在西南印度洋附近海域其他人的研究也呈现类似结果。以2012年至2016年东印度洋浮游植物调查为例, 调查时间都在3~5月期间, 且调查海域均在80°~98°E, 10°N~10°S区域内, 但每年的站位点数量与经纬度位置存在差异。2013年调查采用了77 μm孔径的浮游生物网, 其余年份均采用20 μm孔径的网具。2012年至2016年调查鉴定结果如表 3所示。其中, 由于2013年调查的站位较多, 因此结果中总物种数量仅次于2012年调查结果, 但站位平均物种数最少, 表明使用77 μm孔径的网具采样时, 鉴定到的物种数量少于使用20 μm孔径的网具采样。此外, 该海域内甲藻物种比例均高于硅藻, 2013年调查鉴定所得硅藻物种数与甲藻物种数分别占总物种数的46.29%和56.29%, 而其他年份鉴定得到的甲藻物种数均为硅藻物种数的两倍左右, 表明使用20 μm孔径的网具可以截留到个体更小的甲藻(薛冰, 2017; 李艳等, 2019)。
表 3 东印度洋浮游植物历史数据比较 Tab. 3 Historical data of phytoplankton assemblage in the Eastern Indian Ocean
| 年份 | 网具孔径/μm | 门 | 总物种数 | 站位平均物种数 | 硅藻占比/% | 甲藻占比/% | 细胞丰度/(×103 cells/m3) | 参考文献 |
| 2012 | 20 | 4 | 408 | 12 | 27.21 | 73.28 | 2.907~303.537 | (薛冰, 2017)16~17 |
| 2013 | 77 | 4 | 350 | 4 | 46.29 | 52.29 | 0.5~140.3 | (李艳等, 2019)827 |
| 2014 | 20 | 4 | 310 | 7 | 36.45 | 58.06 | 0.158~8.972 | (薛冰, 2017)16~17 |
| 2015 | 20 | 4 | 340 | 13 | 35.29 | 61.47 | 3.436~111.340 | (薛冰, 2017)16~17 |
| 2016 | 20 | 4 | 292 | 13 | 33.22 | 64.04 | 5.836~3 311.878 | (薛冰, 2017)16~17 |
大洋海域远离大陆, 与近岸海域相比, 较难得到陆源的营养补充, 因此通常为寡营养水域, 而近岸海域往往营养盐丰富。印度洋的营养受限于氮(Moore et al, 2001), 属于寡营养水域。依据浮游植物粒级的划分(Sieburth et al, 1978), 微微型(Pico)粒级浮游植物的大小在0.2~2 μm之间, 微型(Nano)粒级浮游植物的大小在2~20 μm之间, 小型(Net)粒级浮游植物的大小在20~200 μm之间。三种不同粒级的浮游植物在海洋中的分布也有所不同。在寡营养海域, Pico粒级的浮游生物在生物量上占据优势, 而在富营养海域, Net和Nano粒级的浮游生物占据优势(Agawin et al, 2000, 2002; 傅明珠等, 2009; Kostadinov et al, 2010)。其原因在于不同粒级的浮游植物在海洋中的功能有所差异, 以个体较大的浮游植物为主导的群落通常具有较高的光合速率, 初级生产力高; 以微型或微微型浮游植物为主导的群落则具有更为复杂的食物网, 初级生产力较低(Sathyendranath, 2014)。由此推论, 在富营养海域, 例如近岸, 使用77 μm孔径的网具采集网样, 再配以水采样品, 可能已基本了解该海域内的浮游植物群落现状。而且近岸海域的水深较浅, 悬浮颗粒含量远高于大洋环境(薛宇欢等, 2015), 这容易对采样与鉴定工作造成阻碍。孔径较大的网具可滤去部分细微泥沙, 以防网眼堵塞, 利于采样和样品的鉴定。在寡营养海域, 水质较为清澈, 粒级较小的浮游植物占主导地位, 因此采用筛绢较细密的网具更为适宜。此外, 在本次研究中我们没有在近岸营养丰富海域开展不同孔径网具的调查对比, 这将是进一步的研究内容。
4 结论从2015年西南印度洋第34航次的浮游植物调查中, 可以得到以下结果:
(1) 使用20 μm孔径的网具采到的样品物种数和细胞丰度都显著高于采用77 μm孔径的网具。在本次研究结果中, 采用20 μm孔径网具采得的平均物种数与平均细胞丰度分别约为采用77 μm孔径网具的2.7倍与17倍, 呈显著差异。
(2) 20和77 μm两种不同孔径的网采样品之间, 平均多样性指数分别为2.23和2.72, 平均均匀度指数分别为0.69和0.64, 结果无显著差异。
(3) 采用20和77 μm两种不同孔径的网具, 采得样品之间的平均物种丰富度指数分别为6.69和3.31, 前者约为后者的2倍, 呈显著差异。
(4) 筛绢孔径为20 μm的网采样品与筛绢孔径为77 μm的网采样品分析结果中分别得出的优势种也存在差异。前者有9种, 后者有11种, 共同的优势种有6种。
(5) 应根据具体调查区域与研究目的选择合适孔径网具, 本文的研究结果表明在寡营养海域更适宜使用20 μm孔径的网具。
参考文献
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6—1991[S]. 北京: 中国标准出版社: 157.
|
孙军, 刘东艳, 冯士筰, 2003. 近海生态系统动力学研究中浮游植物采样及分析策略. 海洋与湖沼, 34(2): 224-232 DOI:10.3321/j.issn:0029-814X.2003.02.014 |
李艳, 孙萍, 袁超, 等, 2019. 东印度洋南部春季网采浮游植物群落特征及长期变化. 海洋环境科学, 38(6): 825-832 |
陈卓, 孙军, 张桂成, 2018. 2016年秋季热带西太平洋网采浮游植物群落结构. 海洋科学, 42(7): 114-130 |
傅明珠, 王宗灵, 孙萍, 等, 2009. 南黄海浮游植物初级生产力粒级结构与碳流途径分析. 海洋学报(中文版), 31(06): 100-109 DOI:10.3321/j.issn:0253-4193.2009.06.011 |
胡翠林, 金海卫, 李振华, 等, 2015. 赤潮生物夜光藻的研究进展. 浙江海洋学院学报(自然科学版), 34(4): 379-386 DOI:10.3969/j.issn.1008-830X.2015.04.016 |
洪丽莎, 王春生, 周亚东, 等, 2012. 夏季西南印度洋叶绿素a分布特征. 生态学报, 32(14): 4525-4534 |
郭术津, 孙晓霞, 郑珊, 等, 2018. 热带西太平洋Y3和M2海山区网采浮游植物群落结构特征. 海洋科学, 42(10): 31-40 DOI:10.11759/hykx20180528003 |
薛冰, 2017. 春季季风间期东印度洋赤道及其邻近海域浮游植物群落结构年际变化[D]. 天津: 天津科技大学: 16-17.
|
薛宇欢, 熊学军, 刘衍庆, 2015. 中国近海海水透明度分布特征与季节变化. 海洋科学进展, 33(1): 38-44 DOI:10.3969/j.issn.1671-6647.2015.01.005 |
AGAWIN N S R, AGUSTÍ S, DUARTE C M, 2002. Abundance of Antarctic picophytoplankton and their response to light and nutrient manipulation. Aquatic Microbial Ecology, 29(2): 161-172 |
AGAWIN N S R, DUARTE C M, AGUSTI S, 2000. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnology and Oceanography, 45(3): 591-600 DOI:10.4319/lo.2000.45.3.0591 |
BROTAS V, BREWIN R J W, SÁ C, et al, 2013. Deriving phytoplankton size classes from satellite data: validation along a trophic gradient in the eastern Atlantic Ocean. Remote Sensing of Environment, 134: 66-77 DOI:10.1016/j.rse.2013.02.013 |
HONG L S, WANG C S, ZHOU Y D, et al, 2012. The distribution of chlorophyll a in the tropical eastern Indian Ocean in austral summer. Acta Oceanologica Sinica, 31(5): 146-159 DOI:10.1007/s13131-012-0244-6 |
KOSTADINOV T S, SIEGEL D A, MARITORENA S, 2010. Global variability of phytoplankton functional types from space: assessment via the particle size distribution. Biogeosciences, 7(10): 3239-3257 DOI:10.5194/bg-7-3239-2010 |
MACHU E, BIASTOCH A, OSCHLIES A, et al, 2005. Phytoplankton distribution in the Agulhas system from a coupled physical–biological model. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 52(7): 1300-1318 DOI:10.1016/j.dsr.2004.12.008 |
MOORE J K, DONEY S C, GLOVER D M, et al, 2001. Iron cycling and nutrient-limitation patterns in surface waters of the World Ocean. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 49(1/2/3): 463-507 |
ODATE T, 1997. Abundance and size composition of the summer phytoplankton communities in the western North Pacific Ocean, the Bering Sea, and the Gulf of Alaska. Oceanographic Literature Review, 44(1): 41 |
POVINEC P P, BREIER R, COPPOLA L, et al, 2011. Tracing of water masses using a multi isotope approach in the southern Indian Ocean. Earth and Planetary Science Letters, 302(1/2): 14-26 |
READ J F, LUCAS M I, HOLLEY S E, et al, 2000. Phytoplankton, nutrients and hydrography in the frontal zone between the Southwest Indian Subtropical gyre and the Southern Ocean. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 47(12): 2341-2367 DOI:10.1016/S0967-0637(00)00021-2 |
SATHYENDRANATH S, 2014. Phytoplankton functional types from Space [R]. Dartmouth: International Ocean Colour Coordinating Group.
|
SAVIDGE G, GILPIN L, 1999. Seasonal influences on size-fractionated chlorophyll a concentrations and primary production in the north-west Indian Ocean. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 46(3/4): 701-723 |
SIEBURTH J M, SMETACEK V, LENZ J, 1978. Pelagic ecosystem structure: heterotrophic compartments of the plankton and their relationship to plankton size fractions. Limnology and Oceanography, 23(6): 1256-1263 DOI:10.4319/lo.1978.23.6.1256 |