 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 邱科程, 张荣良, 晁敏, 程方平, 曾江宁, 彭欣, 冯志华, 陈全震, 徐旭丹, 黄伟. 2022.
- QIU Ke-Cheng, ZHANG Rong-Liang, CHAO Min, CHENG Fang-Ping, ZENG Jiang-Ning, PENG Xin, FENG Zhi-Hua, CHEN Quan-Zhen, XU Xu-Dan, HUANG Wei. 2022.
- 浙江三门湾鱼卵、仔稚鱼群聚结构时空变化研究
- SPATIOTEMPORAL VARIATIONS IN CLUSTER STRUCTURE OF FISH EGGS, LARVAE, AND JUVENILES IN THE SANMEN BAY, ZHEJIANG, EAST CHINA SEA
- 海洋与湖沼, 53(3): 684-696
- Oceanologia et Limnologia Sinica, 53(3): 684-696.
- http://dx.doi.org/10.11693/hyhz20211100285
文章历史
-
收稿日期:2021-11-22
收修改稿日期:2022-02-26




2. 自然资源部第二海洋研究所 自然资源部海洋生态系统动力学重点实验室 浙江杭州 310012;
3. 中国水产科学研究院东海水产研究所 上海 200090;
4. 浙江省海洋水产养殖研究所 浙江温州 325005
 (Konosirus punctatus)、鲷科未定种(Sparidae)、蓝点马鲛(Scomberomorus niphonius)、
(Konosirus punctatus)、鲷科未定种(Sparidae)、蓝点马鲛(Scomberomorus niphonius)、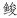 (Liza haematocheila)、舌鳎属(Cynoglossus spp.)、石首鱼科未定种(Sciaenidae)、小公鱼属(Stolephorus spp.); 仔稚鱼优势种为、孔虾虎鱼(Trypauchen vagina)、小公鱼属、细棘虾虎鱼属(Acentrogobius spp.)、缟虾虎鱼属(Tridentiger spp.)。(2) 三门湾湾内与湾外鱼卵、仔稚鱼群聚的组成存在显著差异, 且湾内密度均高于湾外, 反映出优势种鱼类在产卵季对于水文环境(如海流、温度)的空间响应。(3) 三门湾鱼卵、仔稚鱼的群聚组成季节波动明显, 主要由鱼类产卵的季节节律以及以洄游习性等差异形成。结果表明, 三门湾鱼卵、仔稚鱼群聚结构空间、季节差异明显, 与历史资料相比, 鱼卵、仔稚鱼资源从“年际”短时间尺度看已初步显现一定恢复态势。
(Liza haematocheila)、舌鳎属(Cynoglossus spp.)、石首鱼科未定种(Sciaenidae)、小公鱼属(Stolephorus spp.); 仔稚鱼优势种为、孔虾虎鱼(Trypauchen vagina)、小公鱼属、细棘虾虎鱼属(Acentrogobius spp.)、缟虾虎鱼属(Tridentiger spp.)。(2) 三门湾湾内与湾外鱼卵、仔稚鱼群聚的组成存在显著差异, 且湾内密度均高于湾外, 反映出优势种鱼类在产卵季对于水文环境(如海流、温度)的空间响应。(3) 三门湾鱼卵、仔稚鱼的群聚组成季节波动明显, 主要由鱼类产卵的季节节律以及以洄游习性等差异形成。结果表明, 三门湾鱼卵、仔稚鱼群聚结构空间、季节差异明显, 与历史资料相比, 鱼卵、仔稚鱼资源从“年际”短时间尺度看已初步显现一定恢复态势。2. Key Laboratory of Marine Ecosystem Dynamics, Ministry of Natural Resources, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou 310012, China;
3. East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China;
4. Zhejiang Institute of Marine Aquaculture, Wenzhou 325005, China
鱼类早期生活史阶段包括鱼卵、仔鱼和稚鱼期, 是其生命中最脆弱和对栖息环境变化最敏感的阶段(Houde, 1987; 殷名称, 1991)。鱼类群聚是指生活在同一水域或生境中各种鱼类的聚合体, 鱼类早期生活史阶段的成活率直接关系到渔业资源的年际补充量, 是影响鱼类早期生活史群聚结构变化的主要因素(Chambers et al, 1997)。研究鱼卵、仔稚鱼群聚结构是评估渔业资源可持续利用度的重要组成, 对于量化海洋鱼类资源分布、有效实现鱼类物种的增殖养护、确定渔场和渔汛等都具有重要意义(Clark et al, 1953)。
海湾生境多样、饵料丰富, 有利于生物的繁殖和生长, 为海洋鱼类提供了重要产卵场、育幼场和索饵场(Archambault et al, 1999)。鱼类早期资源的群聚组成与海湾环境有密切的联系, 良好的海湾环境有利于提高鱼类早期资源的存活率, 并在塑造鱼类群聚结构和空间分布以及物种多样性方面发挥了重要作用(Bradbury et al, 2008; Álvarez et al, 2015)。三门湾位于浙江省中部沿海, 海域面积775 km2, 是浙江省第二大海湾, 入湾河流30多条, 其中以青溪、白溪和珠游溪最大(曾江宁等, 2011)。三门湾水质肥沃, 为鱼类产卵育肥提供了良好条件, 使其成为东海渔场的重要组成部分, 是蓝点马鲛(Scomberomorus niphonius)、鳓(Ilisha elongata)、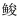
目前, 关于三门湾渔业资源的研究资料十分有限, 仅开展了三门湾游泳动物种类组成、群聚结构特征及物种多样性等少量研究(蔡萌等, 2009; 李新等, 2014; 冀萌萌等, 2016)。但有关三门湾鱼卵、仔稚鱼群聚结构等未见相关报道。本文基于三门湾春、夏两季调查资料, 分析了该海域鱼卵、仔稚鱼的种类组成、时空分布、群聚结构特征等, 探讨了群聚结构与环境因子的相关性。旨在了解环境变异条件下三门湾鱼类种群早期补充群体分布特征, 为科学认识渔业种群早期生活史补充过程提供基础资料和数据支撑。
1 材料与方法 1.1 调查站位和方法三门湾三面环陆, 是呈西北-东南走向的半封闭海湾, 在该海域设置20个调查站位(121.45°~122.18°E, 28.65°~29.36°N), 根据底质类型、水文条件将其分为湾内(1~10)、湾外(11~20)两个区域(图 1) (聂振林等, 2018)。分别开展夏(2019年8月)、春(2020年4月)两季的鱼卵、仔稚鱼资源调查。依照《海洋调查规范》(中华人民共和国国家质量监督检验检疫总局等, 2018), 采用浅水Ⅰ型浮游生物网(网口面积0.2 m2, 网口直径50 cm, 网长145 cm)定量采样, 由海底至海面进行垂直拖网, 网口均配备流量计(Hydro-Bios- 438115, 德国)记录拖网流量值; 采用大型浮游生物网(网口面积0.5 m2, 网口直径80 cm, 网长280 cm)定性采样, 水平拖网10 min, 平均拖速2 kn。同时, 使用多参数水质分析仪(YSI-Pro, 美国)现场测量水温、pH、溶解氧、盐度等环境参数。样品用5%中性甲醛溶液固定保存, 待返回实验室后进行挑选、分类鉴定并计数, 物种鉴定主要参考《中国近海鱼卵与仔鱼》(张仁斋, 1985)、《中国近海及其临近海域鱼卵与仔稚鱼》(万瑞景等, 2016)、《长江口鱼类》(庄平等, 2006)等资料。鱼卵、仔稚鱼的数量分布以每一站位实际采获的个体数表示, 单位分别为粒和尾。
 |
| 图 1 三门湾鱼卵、仔稚鱼调查站位 Fig. 1 Sampling stations of fish eggs, larvae and juveniles in the Sanmen Bay |
(1) 鱼卵、仔稚鱼密度G (ind./m3)
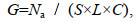 (1)
(1)式中, G为单位体积海水中鱼卵(粒/m3)或仔稚鱼个体数(尾/m3); Na为全网鱼卵或仔稚鱼个体数; S为网口面积(m2); L为流量计转数; C为流量计校正值(C=0.3) (国家海洋局908专项办公室, 2006)。
(2) 物种优势度及更替率
用优势度(Y)确定鱼卵、仔稚鱼的优势种类, 其计算公式为:
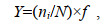 (2)
(2)式中, ni为第i种鱼的鱼卵(或仔稚鱼)数量, N为所有种类的鱼卵(或仔稚鱼)总数量, f为站位出现频率, 取优势度大于0.02的为优势种(McNaughton et al, 1967)。
使用种类组成更替率(R)来衡量鱼卵、仔稚鱼种类组成的季节更替情况, 其公式为:
 (3)
(3)式中, a和b分别表示相邻两个季节的物种的种类数, c为两个季节共有物种的种类数(刘瑞玉, 1992)。
(3) 群聚结构分析
采用Primer 6.0软件进行聚类分析, 为平衡优势种和非优势种在群聚中的作用, 计算前对原始数据矩阵进行lg(x+1)变换, 再计算Bray-Curtis距离, 建立距离矩阵, 最后采用类平均法进行聚类并作树状图(Clarke, 1993; Brazner et al, 1997)。为了验证聚类结果的可信度, 在此基础上进行了非线性多维标度排序分析(Nonmetric Multidimensional Scaling, NMDS); 应用相似性检验(Analysis of similarities, ANOSIM)非参数多元分析方法(Clarke, 1993)检验群落之间的差异显著性; 应用相似性百分比法(Similarity Percentage, SIMPER)分析鱼类对站位组内相似性和组间相异性的贡献率; 用胁强系数(stress)来衡量NMDS结果的优劣, 此值越小越好, 当stress < 0.2时, 可用NMDS的二维点图表示, 其图形有一定的解释意义; 当stress < 0.1时, 可以认为是一个好的排序; 当stress < 0.05时, 具有很好的代表性(Khalaf et al, 2002)。
(4) 环境因子分析
采用Canoco 5.0分析鱼类物种与环境因子之间的关系并绘制排序图, 以揭示不同环境因子对鱼卵、仔稚鱼的影响。首先对物种数据进行消除趋势对应分析(Decentred Correspondence Analysis, DCA), 根据每个轴的梯度长度(Length of Gradient Axis, LGA)选择适宜的排序方法。当LGA < 3时, 选择冗余分析(Redundancy Analysis, RDA); 当LGA > 4时, 则可运用典范对应分析(Canonical Correspondence Analysis, CCA); 当3 < LGA < 4时, 两者皆可(Lepš et al, 2003)。根据DCA分析结果, 本文采用线性模型的RDA分析。
采用SPSS 23.0软件进行单因素方差分析(One-way ANOVA), 检验物种丰度时空差异性。
2 结果 2.1 种类及优势种组成两季共鉴定出鱼卵、仔稚鱼8目21科40种, 其中定性样品鉴定出鱼卵、仔稚鱼8目21科36种, 定量样品共4目11科24种(详见附录)。定量样品中鲈形目(Perciformes)种类最多, 鉴定出7科17种, 占总采获量的51.9%; 鲽形目(Pleuronectiformes)最少, 占总采获量的10.6%。春季调查中, 鉴定出鱼卵4种, 隶属于3目4科, 仔稚鱼6种, 隶属于3目5科, 鱼卵、仔稚鱼共有种3种; 夏季调查中, 鉴定出鱼卵5种, 隶属于3目5科, 仔稚鱼13种, 隶属于3目6科, 鱼卵、仔稚鱼共有种1种; 夏季鱼卵、仔稚鱼采获量均最多, 分别占总采获量的20.6%和51.3%; 鱼卵采获量最少的是春季, 占总采获量的7.5%。
| 中文名 | 学名 | 春季 | 夏季 |
| 鲱形目 | Clupeiformes | ||
| 鲱科 | Clupeidae | ||
斑
|
Konosirus punctatus | ± ▼△ | |
| 鳓鱼 | Ilisha elongata | ▼ | |
| 鲱科未定种 | Clupeidae | + ▼ | |
| 鳀科 | Engraulidae | ||
| 棱鳀属 | Thryssa spp. | - | |
| 日本鳀 | Engraulis japonicus | △ | |
| 小公鱼属 | Stolephorus spp. | ± ▼△ | |
| 鲻形目 | Mugiliformes | ||
| 鲻科 | Mugilidae | ||
|
|
Liza haematocheila | ± ▼△ | |
| 鲈形目 | Perciformes | ||
| 真鲈科 | Percichthyidae | ||
| 花鲈 | Lateolabrax japonicus | △ | |
| 鲷科 | Sparidae | ||
| 黑鲷 | Acanthopagrus schlegelii | - △ | |
| 鲷科未定种 | Sparidae | + ▼△ | |
| 石首鱼科 | Sciaenidae | ||
| 白姑鱼 | Argyrosomus argentatus | - | |
| 黄姑鱼 | Nibea albiflora | - △ | |
| 小黄鱼 | Larimichthys polyactis | ▼△ | |
| 石首鱼科未定种 | Sciaenidae | + ▼ | |
| 虾虎鱼科 | Gobiidae | ||
| 缟虾虎鱼属 | Tridentiger spp. | - △ | |
| 沟虾虎鱼属 | Oxyurichthys spp. | - △ | |
| 拟矛尾虾虎鱼 | Parachaeturichthys polynema | - △ | |
| 矛尾虾虎鱼 | Chaemrichthys stigmatias | - △ | |
| 舌虾虎鱼属 | Glossogobius spp. | - △ | |
| 细棘虾虎鱼属 | Acentrogobius spp. | - △ | |
| 六丝钝尾虾虎鱼 | Amblychaeturichthys hexanema | - △ | |
| 深虾虎鱼 | Bathygobius fuscus | - | |
| 斑尾刺虾虎鱼 | Acanthogobius ommaturus | △ | |
| 竿虾虾虎鱼 | Luciogobius guttatu | △ | |
| 带鱼科 | Trichiuridae | ||
| 小带鱼 | Eupleurogrammus muticus | ▼△ | + ▼ |
 科 科 |
Callionymidae | ||
| 科 |
Callionymidae | ▼△ | |
| 鲭科 | Scombrida | ||
| 蓝点马鲛 | Scomberomorus niphonius | ± ▼△ | |
| 鳗虾虎鱼科 | Taenioidae | ||
| 孔虾虎鱼 | Trypauchen vagina | - △ | |
| 弹涂鱼科 | Periophthalmidae | ||
| 弹涂鱼 | Periophthalmus modestus | - △ | |
 科 科 |
Sillaginidae | ||
| 多鳞鱚 | Sillago sihama | △ | |
| 少鳞鱚 | Sillago japonica | △ | |
| 塘鳢科 | Eleotridae | ||
| 锯塘鳢 | Prionobutis koilomatodon | △ | |
| 鲽形目 | Pleuronectiformes | ||
| 舌鳎科 | Cynoglossoidae | ||
| 焦氏舌鳎 | Cynoglossus joyneri | - | |
| 舌鳎属 | Cynoglossus spp. | ▼ | + |
| 牙鲆科 | Paralichthyidae | ||
| 斑鲆 | Pseudorhombus arsius | △ | |
| 鲀形目 | Tetraodontiformes | ||
| 鲀科 | Tetraodontidae | ||
| 东方鲀属 | Takifugu | △ | |
| 鲉形目 | Scorpaeniformes | ||
| 鲉科 | Scorpaenidae | ||
| 褐菖鲉 | Sebastiscus marmoratus | △ | |
| 鲬科 | Platycephalidae | ||
| 鲬 | Platycephalus indicus | △ | |
| 颌针鱼目 | Beloniformes | ||
| 颌针鱼科 | Belone | ||
| 颌针鱼属 | Belone spp. | △ | |
| 鳚形目 | Blenniiformes | ||
| 鳚科 | Blenniidae | ||
| 美肩鳃鳚 | Omobranchus elegans | △ | |
| 注: “+”表示定量鱼卵样品中出现, “-”表示定量仔稚鱼样品中出现, “▼”表示定性鱼卵样品中出现, “△”表示定性仔稚鱼样品中出现 | |||
根据优势度(Y)大于0.02为优势种的原则, 该调查海域鱼卵和仔稚鱼优势种共有10种, 优势度最高的物种为
| 种名 | 春季 | 夏季 | 生态类型及参考文献 | |||
| 鱼卵 | 仔稚鱼 | 鱼卵 | 仔稚鱼 | |||
| 斑 Konosirus punctatu |
0.038 | Es (中国科学院中国动物志编辑员会, 2001) | ||||
| 鲷科未定种Sparidae | 0.038 | Ma (樊冀蓉等, 2011) | ||||
| 蓝点马鲛Scomberomorus niphonius | 0.025 | Ma (邱盛尧等, 1996) | ||||
| Liza haematocheila |
0.025 | 0.303 | Es (钟俊生等, 2006) | |||
| 舌鳎属Cynoglossus spp. | 0.079 | Ma (宋超等, 2014) | ||||
| 石首鱼科未定种Sciaenidae | 0.082 | Ma (郭昶畅, 2017) | ||||
| 小公鱼属Stolephorus spp. | 0.053 | 0.046 | Es (吴光宗, 1989) | |||
| 细棘虾虎鱼属Acentrogobius spp. | 0.024 | Es (钟俊生, 1997) | ||||
| 缟虾虎鱼属Tridentiger spp. | 0.027 | Ma (何丽斌等, 2010) | ||||
| 孔虾虎鱼Trypauchen vagina | 0.124 | Es (金逍逍, 2013) | ||||
| 注: Es表示河口性鱼类; Ma表示海洋性鱼类。下同 | ||||||
 |
| 图 2 三门湾鱼卵、仔稚鱼优势种季节占比 Fig. 2 Seasonal proportion of dominant species of fish eggs, larvae and juveniles in the Sanmen Bay |
| 区域 | 种名 | 春季 | 夏季 | 生态类型 | ||
| 鱼卵 | 仔稚鱼 | 鱼卵 | 仔稚鱼 | |||
| 湾内 | 斑鰶 | 0.075 | 0.027 | Es | ||
| 鲷科未定种 | 0.075 | Ma | ||||
| 蓝点马鲛 | 0.025 | Ma | ||||
| 鮻 | 0.050 | 0.382 | Es | |||
| 鲱科Clupeidae | 0.021 | Ma | ||||
| 孔虾虎鱼 | 0.157 | Es | ||||
| 舌鳎属 | 0.186 | Ma | ||||
| 石首鱼科 | 0.100 | Ma | ||||
| 细棘虾虎鱼属 | 0.034 | Es | ||||
| 小公鱼属 | 0.029 | Es | ||||
| 湾外 | 鮻 | 0.114 | Es | |||
| 孔虾虎鱼 | 0.077 | Es | ||||
| 石首鱼科未定种 | 0.080 | Ma | ||||
| 小公鱼属 | 0.180 | 0.119 | Es | |||
| 缟虾虎鱼属 | 0.038 | Ma | ||||
三门湾鱼卵、仔稚鱼密度的季节及空间变化如图 3所示, 统计结果显示, 春季鱼卵、仔稚鱼的平均密度均显著低于夏季(ANOVA, P < 0.05), 其中春季鱼卵和仔稚鱼的平均密度分别为0.40 ind./m3和1.04 ind./m3; 夏季鱼卵和仔稚鱼的平均密度分别为0.62 ind./m3和1.70 ind./m3; 此外, 图 3a中出现的异常值, 最大值为夏季仔稚鱼5号站位。空间分布见图 3b, 湾内春、夏两季鱼卵、仔稚鱼密度均高于湾外(1.34 ind./m3 vs 0.54 ind./m3), 除了夏季湾内仔稚鱼密度与湾外比较无显著差异(P > 0.05)外, 春季湾内鱼卵、仔稚鱼密度及夏季湾内鱼卵密度与湾外比较均具有显著差异(P < 0.05)。
 |
| 图 3 三门湾鱼卵、仔稚鱼密度季节及空间变化 Fig. 3 Seasonal and spatial variation in the density of fish eggs, larvae and juveniles in the Sanmen Bay 注: a. 鱼卵、仔稚鱼密度季节变化; b. 鱼卵、仔稚鱼密度空间变化; *表示组间具有显著差异 |
三门湾20个调查站位鱼卵、仔稚鱼群聚结构季节变化聚类分析结果如图 4所示(其中春季5、6、8、13~18号站, 夏季2、14、18、20号站均无采获, 为减少误差故不作统计)。春、夏两个季节的群聚划分明显(7.4%的相似性水平), 相似性检验(ANOSIM)非参数多元分析结果显示2个群聚差异极显著(R=0.589, P < 0.01), 反映出鱼卵、仔稚鱼群聚结构的季节演替明显(图 4a)。NMDS排序的胁强系数为0.01 (stress < 0.05), 说明NMDS排序图具有很好的代表性(图 4b)。两季物种更替率R达到100%, 进一步说明了三门湾鱼卵、仔稚鱼群聚结构季节变化具有极显著的差异。另外, 结合相似性百分比(SIMPER)分析, 结果显示春季形成的湾内、湾外群聚结构差异显著(P < 0.05) (图 4c), 主要分歧种有
 |
| 图 4 三门湾鱼卵、仔稚鱼群聚结构季节变化聚类图、排序图 Fig. 4 Clustering and sorting charts of seasonal changes in cluster structures of fish eggs, larvae and juveniles in the Sanmen Bay 注: a、b. 季节性聚类变化; c. 春季; d. 夏季。 |
| 季节 | 区域 | R | P | 主要分歧物种 | 组间相异性贡献率 |
| 春 | 湾内-湾外 | 0.451 | 0.02 | 、斑、矛尾虾虎鱼 |
76.5% |
| 夏 | 湾内-湾外 | 0.143 | 0.08 | 小公鱼属、孔虾虎鱼、舌鳎属 | 52.9% |
三门湾海域鱼卵、仔稚鱼优势种与环境因子的RDA分析结果如图 5所示。春季, 5个环境因子解释了物种-环境关系的45.8%, 第一RDA轴(RDA1, 特征值=0.425)和第二RDA轴(RDA2, 特征值=0.012)表现出很高的物种-环境相关性(RDA1和RDA2与环境因子相关性分别为0.705和0.458)。蒙特卡洛显著性检验结果显示, 第一排序轴和所有排序轴均差异不显著(第一轴: F=10.3, P=0.09; 所有轴: F=2.2, P=0.09)。影响三门湾春季鱼卵、仔稚鱼群聚的主要环境因子是水温、pH、溶解氧, 其中第一排序轴与水温呈最强正相关, 与水深呈负相关, 第二排序轴与溶解氧呈正相关, 与水深呈最大负相关。结果还表明斑鲫、
 |
| 图 5 三门湾海域鱼卵、仔稚鱼优势种密度与环境因子之间的关系 Fig. 5 Relationship between density of dominant species of fish eggs, larvae and juveniles and environmental factors in the Sanmen Bay |
本研究中定量样品鉴定出的鱼卵、仔稚鱼隶属于4目11科24种, 其中鲈形目种类最多, 为7科17种, 占总采获量的51.9%, 与舟山渔场(鲈形目共19种, 占47.50%)海域研究结果相似(刘连为等, 2021)。三门湾鱼卵主要优势种为斑
20世纪50~60年代, 三门湾渔业种类主要是小黄鱼、带鱼(Trichiurus lepturus)和鲆鲽类等经济价值较高的底层、近底层鱼类(Jin, 2004)。20世纪70~80年代以后, 违反自然规律的过度捕捞已远超渔业资源的再生能力, 使经济价值较高的鱼类资源量不断下降(凌建忠等, 2006)。此外, 围填海、海底管线等近岸工程, 进一步破坏了三门湾传统渔业生物的产卵场和栖息地, 也会影响渔业资源补充过程(黄潘阳等, 2017)。本调查采集到的鱼卵、仔稚鱼以小型低值鱼类如虾虎鱼、斑
本调查中鱼卵、仔稚鱼密度与长江口、舟山渔场、岱衢洋等邻近海域相近(林楠等, 2016; 樊紫薇等, 2020; 刘连为等, 2021)。尽管从“代际”中时间尺度看, 三门湾渔业资源严重衰退, 但从“年际”短时间尺度上看, 本次调查仔稚鱼密度较2013年(仔稚鱼0.49 ind./m3)已有回升趋势(表 4)。其原因可能是: 近年来三门湾周边市县实施限制捕捞规格、伏季休渔期、增殖放流等相关政策, 尤其是2017年史上最严休渔制度的实施发挥了积极作用, 仔稚鱼密度变化趋势反映了三门湾渔业资源正逐步恢复(邵晨等, 2018)。三门湾增殖放流时间较早, 从20世纪90年代即开始放流对虾, 至2019年, 三门湾周边市县增殖放流苗种达2.4亿尾(头)/年, 主要品种为大黄鱼、黑鲷、黄姑鱼、斑
| 位置 | 调查时间(年.月) | 鱼卵/(ind./m3) | 仔稚鱼/(ind./m3) | 鱼卵优势种 | 仔稚鱼优势种 | 参考文献 |
| 长江口 | 1985.08~1986.10 | — | — | 凤鲚、大银鱼、康氏小公鱼、大黄鱼、小黄鱼 | 鳀、鲐 | (杨东莱等, 1990) |
| 长江口 | 1985.08~1986.08 | — | — | 鳀、康氏小公鱼 | 鳀、康氏小公鱼 | (吴光宗, 1989) |
| 长江口 | 1997.05 | — | — | 凤鲚 | 日本鳀、凤鲚 | (徐兆礼等, 1999) |
| 长江口 | 2000.05~2003.08 | — | — | 凤鲚、中华小公鱼、鳀 | 凤鲚、中华小公鱼、鳀 | (蒋玫等, 2006) |
| 长江口 | 2008.05 | — | 0.26 | — | 日本鳀、六丝矛尾虾虎鱼 | (李建生等, 2015) |
| 长江口 | 2006~2007 | — | 0.01~1.85 | — | 鳀、凤鲚、长鳍、底灯鱼 | (王小谷等, 2017) |
| 长江口 | 2014.04~07 | 0.01~0.054 | 0.04~0.12 | 小黄鱼、日本鳀、斑尾刺虾虎鱼 | 小黄鱼、日本鳀、矛尾虾虎鱼、斑尾刺虾虎鱼 | (林楠等, 2016) |
| 杭州湾 北部 |
2013.01~12 | — | 1.25 | — | 髭缟虾虎鱼、棘头梅童鱼、凤鲚、 黄鳍刺虾虎鱼、斑尾刺虾虎鱼 |
(闫欣等, 2014) |
| 杭州湾 | 2008.04~06 | — | — | 黄姑鱼、凤鲚、银鲳、皮氏叫姑鱼、鳓、石首鱼科 | 凤鲚、斑、小黄鱼、蓝点马鲛、 棘头梅童鱼、虾虎鱼类 |
(周永东等, 2011) |
| 舟山渔场 | 2017.04~2019.05 | 0.02~0.52 | 0.04~0.20 | 小带鱼、舌鳎科 | 、丝鳍、鳀、矛尾虾虎鱼 |
(刘连为等, 2021) |
| 岱衢洋 | 2010.05~2012.02 | 0.08 | 0.15 | 舌鳎科、小带鱼 | 、丝鳍 、鳀、矛尾虾虎鱼、 、鳀、矛尾虾虎鱼、中国小公鱼 |
(樊紫薇等, 2020) |
| 象山港 | 2015.04~06 | — | 0.08~5.88 | — | 斑、矛尾虾虎鱼、、斑尾刺虾虎鱼、普氏缰虾虎鱼、拟矛尾虾虎鱼、黑棘鲷、舌虾虎鱼 |
(王宇坛等, 2017) |
| 三门湾 | 2013 | 11.04 | 0.49 | 斑、鲻科 |
龟
|
(中国水产科学研究院东海水产研究所, 2014) |
| 瓯江口 | 2015.04, 2015.10 | — | — | 斑、舌鳎科 |
、花鲈、矛尾虾虎鱼、小公鱼属 |
(杜萍等, 2016) |
| 浙江南部 海岸 |
2011.04~06 | 0.04~1.40 | 0.532~2.90 | 斑、棱、小黄鱼、蓝点马鲛、石首鱼科 |
斑、虾虎鱼类、棱
|
(周永东等, 2013) |
| 三门湾 | 2019.08, 2020.04 | 0.40~0.62 | 1.04~1.70 | 斑、鲷科未定种、蓝点马鲛、、舌鳎属、石首鱼科未定种、小公鱼属 |
、小公鱼属、细棘虾虎鱼属、缟虾虎鱼属、孔虾虎鱼 |
本研究 |
本次调查湾内采获的鱼类优势种多于湾外, 湾内采获的主要为鱼卵。从空间分布上看, 湾内外春、夏两季鱼卵、仔稚鱼的群聚结构差异显著, 且湾内的资源密度均高于湾外。导致这一空间分布格局的主要原因为: 首先, 鱼类优势种的生活习性, 尤其是产卵时的生境偏好导致湾内外的资源量差异。SIMPER分析结果显示,
另一方面, 鱼卵和仔稚鱼的生长发育、数量分布等与海流、盐度以及水温等环境因子密切相关(Miller et al, 2009)。环境因子分析结果表明本研究中引起三门湾海域鱼卵、仔稚鱼群聚结构空间变化的主要因子是温度。其中, 春季湾内平均水温(18.44 ℃)明显高于湾外(16.95 ℃), 根据Hurst等(2010)对鱼卵生长动力学研究, 鱼类在产卵期和仔鱼期的大小与温度呈正相关。春季鱼卵、仔稚鱼优势种斑
群聚结构季节变化聚类分析显示, 调查海域鱼卵、仔稚鱼组成存在明显的季节变化, 与长江口(蒋玫等, 2014)、杭州湾(闫欣等, 2014)、舟山群岛外海域(于南京等, 2020)等临近海域鱼卵、仔稚鱼群聚结构季节变化结果相符(陈渊戈等, 2015; 樊紫薇等, 2020)。一方面, 鱼类产卵存在特定的时间节律。通常鱼类的产卵期为该海域鱼卵大量集中出现的时期(赵优等, 2008), 夏季三门湾的鱼卵、仔稚鱼丰度均达到最高值, 意味着该季节多数鱼类进入产卵期。春、夏两季节鱼卵、仔稚鱼物种(尤其是优势种)更替明显, 且三门湾往年渔业资源调查结果显示, 成鱼的优势种更替情况与之相符(冀萌萌等, 2016; 聂振林等, 2018; 俞松立等, 2021)。另一方面, 长距离洄游物种(如蓝点马鲛), 清明节前后进入湾内产卵并滞留到秋季, 其稚鱼主要在岛屿及岩礁附近海域索饵, 且夏季通常栖息于中上层, 故容易被采获。舌鳎属每年12月起向湾外东部海域越冬洄游, 翌年2月向浙江沿海产卵洄游, 主要产卵期是5~8月, 且主要产卵场在湾外(周永东等, 2018)。这些鱼类的产卵洄游习性很大程度上影响了群聚的季节变化。
此外, 三门湾属暖温带海域, 受陆地径流、江浙沿岸流及台湾暖流等水团共同影响, 季节差异明显, 如海域潮流场流速的季节变化为夏季略大于冬季, 秋季略大于春季(曾江宁等, 2011); 寒暖流交汇使海水扰动, 下层营养物质带到表层, 有利于鱼类繁殖索饵, 相互作用下形成鱼卵、仔稚鱼群聚的季节差异(张启龙等, 2007)。
4 结论(1) 三门湾海域鱼卵、仔稚鱼的数量及分布现状: 两季共采获到鱼卵、仔稚鱼8目21科40种, 其中定性样品共采获鱼卵、仔稚鱼8目21科36种, 定量样品共采获鱼卵、仔稚鱼4目11科24种。总体而言, 三门湾鱼卵、仔稚鱼资源密度与周围海域相近, 优势种种类差异不大。
(2) 空间分布上, 三门湾湾内与湾外鱼卵、仔稚鱼群聚的组成存在明显差异。主要由鱼类优势种的生活习性以及产卵时的生境偏好导致湾内外的资源量差异。SIMPER分析结果显示, 其群聚空间差异主要由
(3) 季节变化上, 三门湾春夏两季鱼卵、仔稚鱼优势种更替明显。春季鱼卵、仔稚鱼优势种主要为斑
于南京, 俞存根, 许永久, 等, 2020. 舟山群岛外海域鱼类数量分布与环境因子的关系. 海洋学报, 42(10): 80-91 |
万瑞景, 张仁斋, 2016. 中国近海及其邻近海域鱼卵与仔稚鱼. 上海: 上海科学技术出版社
|
马路阔, 陈全震, 程方平, 等, 2022. 三门湾游泳动物群落结构与生物多样性研究. 海洋与湖沼, 53(2): 384-393 |
王九江, 刘永, 肖雅元, 等, 2019. 大亚湾鱼卵、仔稚鱼种群特征与环境因子的相关关系. 中国水产科学, 26(1): 14-25 |
王小谷, 孙栋, 林施泉, 2017. 长江口及其邻近海域仔鱼的生态学研究. 上海海洋大学学报, 26(5): 733-742 |
王宇坛, 李圣法, 杨林林, 等, 2017. 象山港春、夏季仔稚鱼种类组成结构特征. 海洋渔业, 39(3): 286-296 DOI:10.3969/j.issn.1004-2490.2017.03.006 |
王建茹, 林元烧, 周美玉, 等, 2010. 福建三沙湾鱼卵和仔稚鱼的生态分布. 厦门大学学报(自然科学版), 49(1): 116-121 |
王星火, 常雪晴, 程方平, 等, 2021. 基于2b-RAD简化基因组测序的三门湾海域3种优势鱼类群体遗传多样性分析. 海洋渔业, 43(5): 513-520 DOI:10.3969/j.issn.1004-2490.2021.05.001 |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6-2007 [S]. 北京: 中国标准出版社.
|
中国水产科学研究院东海水产研究所, 2014. 三门核电3、4号机组工程海洋渔业资源调查专题报告[R]: 7-9.
|
中国科学院中国动物志编辑员会, 2001. 中国动物志-硬骨鱼纲, 鲟形目海鲢目鲱形目鼠鱚目. 北京: 科学出版社
|
宁修仁, 2005. 乐清湾、三门湾养殖生态和养殖容量研究与评价. 北京: 海洋出版社
|
朱维新, 1990. 浙江省三门湾开始放流增殖对虾. 现代渔业信息, (6): 29 |
庄平, 王幼槐, 李圣法, 等, 2006. 长江口鱼类. 上海: 上海科学技术出版社
|
刘连为, 隋宥珍, 蒋日进, 等, 2021. 春季舟山渔场鱼卵、仔稚鱼种类组成与数量分布. 浙江海洋大学学报(自然科学版), 40(1): 1-8 |
刘瑞玉, 1992. 胶州湾生态学和生物资源. 北京: 科学出版社
|
闫欣, 钟俊生, 王明星, 等, 2014. 杭州湾北部水域仔稚鱼种类组成和多样性研究. 上海海洋大学学报, 23(5): 765-773 |
杜萍, 黄伟, 徐旭丹, 等, 2016. 2015年春、秋季瓯江口鱼卵和仔稚鱼生态分布. 海洋学研究, 34(4): 54-62 DOI:10.3969/j.issn.1001-909X.2016.04.007 |
李建生, 胡芬, 林楠, 2015. 长江口及邻近海域春季仔、稚鱼的生态分布研究. 南方水产科学, 11(1): 1-8 DOI:10.3969/j.issn.2095-0780.2015.01.001 |
李新, 周青松, 俞存根, 等, 2014. 浙江三门湾春季鱼类种类组成及多样性研究. 浙江海洋学院学报(自然科学版), 33(6): 488-494 DOI:10.3969/j.issn.1008-830X.2014.06.002 |
杨东莱, 吴光宗, 孙继仁, 1990. 长江口及其邻近海区的浮性鱼卵和仔稚鱼的生态研究. 海洋与湖沼, 21(4): 346-355 |
吴光宗, 1989. 长江口海区鳀鱼和康氏小公鱼鱼卵和仔、稚鱼分布的生态特征. 海洋与湖沼, 20(3): 217-229 |
邱盛尧, 叶懋中, 1996. 黄渤海蓝点马鲛繁殖生物学的研究. 海洋与湖沼, 27(5): 463-470, 570 DOI:10.3321/j.issn:0029-814X.1996.05.002 |
何丽斌, 林琪, 黄瑞芳, 等, 2010. 纹稿虾虎鱼胚胎、早期仔鱼发育与盐度的关系. 福建水产, (3): 10-14, 9 DOI:10.3969/j.issn.1006-5601.2010.03.003 |
宋超, 于亚男, 张涛, 等, 2014. 基于线粒体COⅠ基因部分序列的长江口舌鳎科鱼类系统分类研究. 动物学杂志, 49(5): 716-726 |
张仁斋, 1985. 中国近海鱼卵与仔鱼. 上海: 上海科学技术出版社
|
张启龙, 王凡, 赵卫红, 等, 2007. 舟山渔场及其邻近海域水团的季节特征. 海洋学报, 29(5): 1-9 |
张波, 唐启升, 2004. 渤、黄、东海高营养层次重要生物资源种类的营养级研究. 海洋科学进展, 22(4): 393-404 DOI:10.3969/j.issn.1671-6647.2004.04.001 |
陈渊戈, 毛成责, 林楠, 等, 2015. 长江口和杭州湾碎波带仔稚鱼群聚时空分布特征及相关环境因子分析. 中国水产科学, 22(4): 780-790 |
邵晨, 赵晟, 吴婧慈, 等, 2018. 浙江近海海洋生物增殖放流效果评价. 农村经济与科技, 29(5): 90-92 DOI:10.3969/j.issn.1007-7103.2018.05.035 |
林楠, 程家骅, 姜亚洲, 等, 2016. 长江口两种仔稚鱼网具的采集效率比较. 水产学报, 40(2): 198-206 |
国家海洋局908专项办公室, 2006. 海洋生物生态调查技术规程. 北京: 海洋出版社
|
金逍逍, 2013. 东海16种虾虎鱼类线粒体全基因组序列分析及系统发育研究[D]. 舟山: 浙江海洋学院.
|
周永东, 2004. 浙江沿海渔业资源放流增殖的回顾与展望. 海洋渔业, 26(2): 131-139 DOI:10.3969/j.issn.1004-2490.2004.02.011 |
周永东, 李圣法, 2018. 东海区主要经济种类三场一通道及保护区图集. 北京: 海洋出版社
|
周永东, 金海卫, 张洪亮, 等, 2013. 浙江南部沿岸产卵场春、夏季鱼卵、仔稚鱼种类组成与分布. 海洋与湖沼, 44(3): 590-598 |
周永东, 金海卫, 蒋日进, 等, 2011. 浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布. 水产学报, 35(6): 880-889 |
赵优, 庄平, 章龙珍, 等, 2008. 纹缟虾虎鱼胚胎与早期仔鱼的发育特征. 中国水产科学, 15(4): 533-541 DOI:10.3321/j.issn:1005-8737.2008.04.002 |
钟俊生, 1997. 中国细棘虾虎鱼属(鲈形目, 虾虎鱼科)分类再研究. 上海水产大学学报, (3): 200-211 |
钟俊生, 傅萃长, 郁蔚文, 等, 2006. 鮻鱼稚鱼在沿岸碎波带的出现和滞留时间. 上海水产大学学报, 15(3): 281-285 |
俞松立, 王咏雪, 韩晓凤, 等, 2021. 三门湾海域渔业生物群落结构及营养级变化特征. 大连海洋大学学报, 36(3): 478-487 |
聂振林, 王咏雪, 胡成业, 等, 2018. 三门湾春秋季鱼类群落特征及其与环境因子的关系. 水产学报, 42(9): 1390-1398 |
徐开达, 刘子藩, 2007. 东海区大黄鱼渔业资源及资源衰退原因分析. 大连水产学院学报, 22(5): 392-396 DOI:10.3969/j.issn.1000-9957.2007.05.015 |
徐兆礼, 袁骐, 蒋玫, 等, 1999. 长江口鱼卵和仔、稚鱼的初步调查. 中国水产科学, 6(5): 63-64 |
殷名称, 1991. 鱼类早期生活史研究与其进展. 水产学报, 15(4): 348-358 |
凌建忠, 李圣法, 严利平, 2006. 东海区主要渔业资源利用状况的分析. 海洋渔业, 28(2): 111-116 DOI:10.3969/j.issn.1004-2490.2006.02.005 |
郭昶畅, 2017. 中国沿海石首鱼科鱼类的鉴定、分类和分子系统发育研究[D]. 厦门: 厦门大学.
|
黄永春, 胡石柳, 周泽斌, 等, 2006. 鮸状黄姑鱼早期发育特征及对温盐度的适应性. 中国海洋大学学报, 36(1): 101-106 |
黄潘阳, 陈培雄, 来向华, 等, 2017. 三门湾2003-2013年间围涂工程对水动力环境的影响研究. 中国海洋大学学报, 47(10): 91-98 |
蒋玫, 李磊, 沈新强, 等, 2014. 2010-2011年长江河口及邻近水域春夏季仔稚鱼群落结构及其多样性的研究. 海洋学报, 36(6): 131-137 DOI:10.3969/j.issn.0253-4193.2014.06.016 |
蒋玫, 沈新强, 王云龙, 等, 2006. 长江口及其邻近水域鱼卵、仔鱼的种项组成与分布特征. 海洋学报, 28(2): 171-174 DOI:10.3321/j.issn:0253-4193.2006.02.023 |
曾江宁, 潘建明, 梁楚进, 等, 2011. 浙江省重点港湾生态环境综合调查报告. 北京: 海洋出版社
|
蔡萌, 徐兆礼, 2009. 浙江三门湾冬夏季鱼类种类组成和数量变化. 上海海洋大学学报, 18(2): 198-205 |
樊紫薇, 蒋日进, 张洪亮, 等, 2020. 岱衢洋产卵场鱼卵、仔稚鱼群落结构及其与环境因子的关系. 水产学报, 44(4): 606-620 |
樊冀蓉, 吴仁协, 赵元莙, 等, 2011. 中国鲷科鱼类分类和系统发育研究进展. 中国水产科学, 18(2): 472-480 |
冀萌萌, 田阔, 胡成业, 等, 2016. 三门湾春、秋季游泳动物群落组成及物种多样性研究. 水生态学杂志, 37(2): 42-48 |
ÁLVAREZ I, CATALÁN I A, JORDI A, et al, 2015. Interaction between spawning habitat and coastally steered circulation regulate larval fish retention in a large shallow temperate bay. Estuarine, Coastal and Shelf Science, 167: 377-389 DOI:10.1016/j.ecss.2015.10.015 |
ARCHAMBAULT P, BOURGET E, 1999. Influence of shoreline configuration on spatial variation of meroplanktonic larvae, recruitment and diversity of benthic subtidal communities. Journal of Experimental Marine Biology and Ecology, 238(2): 161-184 DOI:10.1016/S0022-0981(98)00146-4 |
BRADBURY I R, LAUREL B J, ROBICHAUD D, et al, 2008. Discrete spatial dynamics in a marine broadcast spawner: re-evaluating scales of connectivity and habitat associations in Atlantic cod (Gadus morhua L.) in coastal Newfoundland. Fisheries Research, 91(2/3): 299-309 |
BRAZNER J C, BEALS E W, 1997. Patterns in fish assemblages from coastal wetland and beach habitats in Green Bay, Lake Michigan: A multivariate analysis of abiotic and biotic forcing factors. Canadian Journal of Fisheries and Aquatic Sciences, 54(8): 1743-1761 DOI:10.1139/f97-079 |
CHAMBERS R C, TRIPPEL E A, 1997. Early Life History and Recruitment in Fish Populations. London: Chapman & Hall, 78-81
|
CLARK F, MARR J, 1953. Part II: Population Dynamics of the Pacific Sardine [R]. Calcofl Progress Report, 4.
|
CLARKE K R, 1993. Non-parametric multivariate analyses of changes in community structure. Austral Ecology, 18(1): 117-143 DOI:10.1111/j.1442-9993.1993.tb00438.x |
HOUDE E D, 1987. Fish early life dynamics and recruitment variability. American Fisheries Society Symposium, 2: 17-29 |
HURST T P, ABOOKIRE A A, KNOTH B, 2010. Quantifying thermal effects on contemporary growth variability to predict responses to climate change in northern rock sole (Lepidopsetta polyxystra). Canadian Journal of Fisheries and Aquatic Sciences, 67(1): 97-107 DOI:10.1139/F09-171 |
JIN X S, 2004. Long-term changes in fish community structure in the Bohai Sea, China. Estuarine, Coastal and Shelf Science, 59(1): 163-171 DOI:10.1016/j.ecss.2003.08.005 |
KHALAF M A, KOCHZIUS M, 2002. Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba, Red Sea. Marine Ecology Progress Series, 239: 287-299 DOI:10.3354/meps239287 |
LEPŠ J, ŠMILAUER P, 2003. Multivariate Analysis of Ecological Data Using CANOCO. New York: Cambridge University Press
|
MCNAUGHTON S J, 1967. Relationships among functional properties of Californian grassland. Nature, 216(5111): 168-169 |
MILLER B S, KENDALL A W JR, 2009. Early Life History of Marine Fishes. Berkeley: University of California Press
|
MUNDAY P L, DIXSON D L, DONELSON J M, et al, 2009. Ocean acidification impairs olfactory discrimination and homing ability of a marine fish. Proceedings of the National Academy of Sciences of the United States of America, 106(6): 1848-1852 DOI:10.1073/pnas.0809996106 |