 2022, Vol. 53
2022, Vol. 53

基于肥满度-最大养殖收益的底播养殖虾夷扇贝(Patinopecten yessoensis)养殖容量评估及养殖区区划

http://dx.doi.org/10.11693/hyhz20211100284
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 南学良, 魏皓, 蒋增杰, 张海彦, 聂红涛. 2022.
- NAN Xue-Liang, WEI Hao, JIANG Zeng-Jie, ZHANG Hai-Yan, NIE Hong-Tao. 2022.
- 基于肥满度-最大养殖收益的底播养殖虾夷扇贝(Patinopecten yessoensis)养殖容量评估及养殖区区划
- PRODUCTION-CARRYING CAPACITY ASSESSMENT FOR CULTURE AREA PLANNING OF PATINOPECTEN YESSOENSIS BASED ON CONDITION INDEX AND THE MAXIMUM TOTAL PROFIT
- 海洋与湖沼, 53(3): 778-788
- Oceanologia et Limnologia Sinica, 53(3): 778-788.
- http://dx.doi.org/10.11693/hyhz20211100284
文章历史
-
收稿日期:2021-11-21
收修改稿日期:2022-02-02
引用本文
南学良, 魏皓, 蒋增杰, 张海彦, 聂红涛. 2022. 基于肥满度-最大养殖收益的底播养殖虾夷扇贝(Patinopecten yessoensis)养殖容量评估及养殖区区划[J]. 海洋与湖沼, 53(3): 778-788

NAN Xue-Liang, WEI Hao, JIANG Zeng-Jie, ZHANG Hai-Yan, NIE Hong-Tao. 2022. PRODUCTION-CARRYING CAPACITY ASSESSMENT FOR CULTURE AREA PLANNING OF PATINOPECTEN YESSOENSIS BASED ON CONDITION INDEX AND THE MAXIMUM TOTAL PROFIT[J]. Oceanologia et Limnologia Sinica, 53(3): 778-788.
基于肥满度-最大养殖收益的底播养殖虾夷扇贝(Patinopecten yessoensis)养殖容量评估及养殖区区划
1. 天津大学海洋科学与技术学院 天津 300072;
2. 天津市生态环境科学研究院 天津 300191;
3. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 山东青岛 266071
2. 天津市生态环境科学研究院 天津 300191;
3. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 山东青岛 266071
摘要:养殖容量评估是合理规划养殖区的基础。基于虾夷扇贝养殖生态系统模型, 模拟了2006年12月至2017年11月不同放苗密度下底播养殖虾夷扇贝的生长情况。在考虑肥满度对虾夷扇贝养殖收益的影响下, 计算最大养殖收益及其对应的最适放苗密度和养殖容量, 并通过最大养殖收益及其年际差异的聚类分析结果进行养殖区区划。结果表明: 考虑扇贝肥满度的养殖收益能很好的抵消扇贝品质对养殖收益的影响; 底播养殖虾夷扇贝的养殖容量和最适放苗密度有明显的时空差异, 獐子岛和小长山岛之间并向长海海域东北部延伸的条带海域是高收益区; 中收益区在高收益区南北两侧; 低收益区位于研究区西北部和南中收益区南部; 研究区南部由于饵料匮乏导致扇贝生长缓慢, 为不适宜养殖区。开阔海域养殖容量也是有限的, 片面增加放苗密度并不能达到增产增收的效果。研究结果可为合理规划和管理养殖区提供参考。
关键词:底播养殖 虾夷扇贝 养殖生态系统模型 养殖容量 养殖分区 长海海域
PRODUCTION-CARRYING CAPACITY ASSESSMENT FOR CULTURE AREA PLANNING OF PATINOPECTEN YESSOENSIS BASED ON CONDITION INDEX AND THE MAXIMUM TOTAL PROFIT
1. School of Marine Science and Technology, Tianjin University, Tianjin 300072, China;
2. Tianjin Academy of Eco-Environmental Sciences, Tianjin 300191, China;
3. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
2. Tianjin Academy of Eco-Environmental Sciences, Tianjin 300191, China;
3. Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
Abstract: The assessment of production-carrying capacity is the basis for rational planning of aquaculture areas. A Yesso scallop culture ecosystem (YeSCE) model was developed in combination with the ecosystem model of ROMS- CoSiNE (Regional Ocean Modelling System coupled with the Carbon, Silicate and Nitrogen Ecosystem) and dynamic energy budget (DEB) model. The YeSCE model was used to simulate the growth of bottom-cultured Yesso scallop under different densities from December 1, 2006 to November 31, 2017 in Changhai sea area in the North yellow Sea. The spatio-temporal distribution of the total profit was calculated in consideration of the fatness of Yesso scallop cultured under different scallop densities. The optimal scallop density and production-carrying capacity at each grid point of the study area were determined under the constraint of the maximum total profit. The aquaculture areas were planned based on the cluster analysis on the maximum total profit and its interannual differences. The results show that the effect of production quality of Yesso scallop on total profit could be better balanced by considering scallop fatness. There were obvious spatio-temporal differences in the production-carrying capacity and optimal scallop density of the bottom-cultured scallop. The high-profit area is a strip zone between Zhangzi Island and Xiaochangshan Island and extends to the northeast of the Changhai sea area as shown in Fig. 6 of this paper. The medium-profit area is located on both sides of the high-profit area. Due to the restriction of bottom water temperature, the northwest of the study area is low-profit area due to low fatness, and the south of the medium-profit area in the center of the study area is also a low-profit area. In the south of the study area (water depth > 50 m), the growth of the Yesso scallop is slow due to the lack of food availability, which is not suitable for Yesso scallop bottom culture. The production-carrying capacity in the open sea is limited. Increasing merely the scallop density did not increase the total profit. The optimal scallop density planned in this study provided a theoretical support for making full use of production-carrying capacity to reach the maximum total profit in the scallop culture.
Key words:
Bottom culture Patinopecten yessoensis aquaculture ecosystem model production-carrying capacity culture area planning Changhai sea area
近年来, 全球捕捞渔业生产已达到极限(FAO, 2020), 水产养殖业为保障人类蛋白供给、降低天然水域水生生物资源利用强度、促进渔业产业兴旺和渔民生活富裕做出了贡献(麦康森, 2016)。但养殖收益和养殖环境之间的矛盾日益突出, 对水产养殖场进行有效的空间规划和管理对改善养殖环境至关重要(Naylor et al, 2021)。加之我国正在加快推进养殖水域滩涂的环境整治、养殖生产布局的优化和养殖空间的拓展(陈家勇, 2019), 对养殖区的选址是水产养殖绿色发展的基础(Naylor et al, 2021)。养殖容量的评估是科学的规划和管理养殖区、优化养殖生产布局的关键。
养殖容量是将容量应用于水产养殖业的一个重要的生态学特例(唐启升, 1996), 是指单位水体内, 在保护环境、节约资源和保证应有效益等各方面都符合可持续发展要求时的最大养殖量(董双林等, 1998), 也就是在充分利用海区的供饵能力和自净能力的基础上, 养殖群体所能维持的最大现存量(杨红生等, 1999)。养殖容量需要做到养殖收益和生态效益的有效统一。由于养殖环境的不稳定, 养殖容量具有明显的动态性(刘剑昭等, 2000; 贾后磊等, 2003)。大多数情况下, 为了获得最大养殖产量而无限度增加放苗密度(即超容量养殖)使得养殖环境恶化反而达不到预想的养殖收益。20世纪70年代日本科学家首先注意到容量对贝类养殖的影响(唐启升, 1996), 通过调整扇贝养殖密度和养殖数量的方法, 使得扇贝产量稳定、病害减少、死亡降低(李庆彪, 1990)。20世纪80年代使用生态模型研究养殖容量成为主流(Carver et al, 1990; Bacher et al, 1997; 朱明远等, 2002; Guyondet et al, 2010; Filgueira et al, 2014; Zhao et al, 2019)。我国对养殖容量的研究始于20世纪90年代初, 李庆彪(1990)介绍了扇贝大量死亡受养殖容量的影响, 并说明确定养殖容量的方法; 刘庆余(1993)研究了紫贻贝养殖容量问题, 阐明了超容量养殖的后果和应对策略; 方建光等(1996a, 1996b)在桑沟湾评估了扇贝、海带、牡蛎等养殖物种的养殖容量, 并提出了贝藻间养的方法; 大亚湾(黎辉等, 1999)、同安湾(杜琦等, 2000)、大港湾(卢振彬等, 2000)、泉州湾(卢振彬等, 2005)等海域也相继开展了对贝类养殖容量的研究, 这一时期主要通过野外实验评估养殖生物在所在区域的养殖容量(Zhang et al, 2006)。随后, 生态模型被用于研究养殖容量, 如朱明远等(2002)在桑沟湾建立了贝藻混养生态模型评估贝类的养殖容量; 随着养殖生态系统模型的发展, 养殖容量的研究在不同物种和地区得到长足发展(史洁等, 2010a, 2010b, 2010c; 魏皓等, 2010; 宋广军等, 2013; 刘学海等, 2015; Zhao et al, 2019), 包括对底播养殖虾夷扇贝养殖容量的评估(Zhao et al, 2019)。这些研究为所在海区的养殖活动提供了有益指导, 但确定养殖容量的方法大多通过模拟在不同放苗量和收获方式下产量的差异来预估。对于贝类而言这种养殖容量的评估方式并不能体现产品的品质继而不能确保最大的养殖收益。虾夷扇贝养殖生态系统模型(Nan et al, 2022)既能给出底播养殖虾夷扇贝的产量又能提供个体肥满度, 在评估底播养殖虾夷扇贝养殖容量时加入肥满度的影响能获得更合理的评估结果, 为长海海域养殖生产布局的优化和养殖空间的拓展提供科学依据。
1 材料与方法 1.1 研究海域长海海域位于黄海北部, 包含獐子岛、大长山岛、小长山岛、广鹿岛、乌莽岛及海洋岛等传统养殖区(图 1)。獐子岛养殖区是目前中国最大的虾夷扇贝底播养殖区(Zhao et al, 2019), 大长山岛和小长山岛附近海域以筏式养殖为主(Yuan et al, 2010; 李洪波等, 2012)。由于我国加强养殖生产布局的优化和养殖空间的拓展(陈家勇, 2019), 因此, 本文假设长海海域均为底播养殖方式来探究底播养殖虾夷扇贝的适宜生长区域, 为后续养殖区规划和养殖空间拓展提供科学支撑。
 |
| 图 1 研究区及虾夷扇贝养殖生态系统模型的网格点分布 Fig. 1 The study area and grid points in the ecosystem model of the Yesso scallop culture |
虾夷扇贝养殖生态系统模型(Nan et al, 2022)由水动力模型ROMS (Regional Ocean Modelling System)、生态模式CoSiNE (The Carbon, Silicate and Nitrogen Ecosystem)以及虾夷扇贝个体生长模型DEB (Dynamic energy budget)耦合而成。模型水平分辨率为1/24°× 1/24°, 整个长海海域在38.70°~39.40°N和122.30°~123.50°E共810个网格点(图 1)。模型中温度、盐度、溶解氧和营养盐的初始场数据来自WOA 13 V2 (World Ocean Atlas 2013 V2)。叶绿素初始条件使用NEMO-PISCES (nucleus for european modelling of the ocean-pelagic interactions scheme for carbon and ecosystem studies)全球模拟结果(https://doi.org/10.48670/moi-00015)。大气强迫数据来自ERA-Interim再分析资料, 包括风应力、热通量、太阳辐射以及淡水通量等。开边界温度、盐度、流速和水位数据来自HYCOM (Hybrid Coordinate Ocean Model, https://hycom.org/)全球再分析结果。开边界处的潮汐数据由OTPS (oregon state university tidal prediction software)模型计算所得, 包括K1、O1、Q1、P1、M2、S2、K2和N2等8个分潮的潮位和潮流数据。叶绿素、溶解氧和营养盐的开边界条件使用的是NEMO- PISCES全球模型的月平均结果。水温和饵料浓度是影响虾夷扇贝生长的关键因素(Nan et al, 2022)。模型基于虾夷扇贝的生理学特性(Kooijman, 2010), 当虾夷扇贝同化的饵料能量小于虾夷扇贝维持自身结构所需能量时受饵料胁迫; 温度超出呼吸活动的耐受范围时受到温度胁迫。模型中的死亡率根据温度和饵料胁迫下虾夷扇贝的生理实验结果(高振锟, 2016)按温度胁迫、饵料胁迫、温度和饵料双重胁迫加入, 取值分别为6.00%、6.33%和13.30%。虾夷扇贝养殖生态系统模型的放苗密度从1 ind/m2到50 ind/m2逐一计算每个网格点扇贝的生长情况, 每年的模拟时间(每年12月1日至次年11月30日)及虾夷扇贝苗种规格(壳长3.80 cm, 软体部干重0.23 g)均参照前人(Jiang et al, 2020)的观测设定。模型可输出温度、叶绿素、浮游生物、虾夷扇贝个体壳长、壳高、湿重、干重和体积等状态变量。Nan等(2022)已对底层水温、叶绿素、壳长、干重等做了严格校验并应用于探究长海海域底播养殖虾夷扇贝生长能力空间差异的原因。
1.3 养殖收益的计算及养殖容量的确定基于养殖容量的定义(董双林等, 1998; 杨红生等, 1999), 本文使用不同放苗密度下的养殖收益作为确定养殖容量和最适放苗密度的参考。仅以产量(以湿重表示)计算得到的养殖收益并不能说明虾夷扇贝的个体品质, 故引入肥满度(Crosby et al, 1990)说明产出虾夷扇贝的肥满情况, 其计算公式如下:
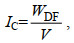 (1)
(1)其中, IC为虾夷扇贝的肥满度(单位: g/cm3); WDF为模型输出的虾夷扇贝个体的软体部干重(单位: g); V表示模型输出的虾夷扇贝的个体体积(单位: cm3)。在于佐安等(2016)计算养殖收益的公式中去掉维护成本、设备成本、环境影响和劳动力成本, 加入Min-max标准化处理之后的肥满度IC用以消除扇贝品质对养殖收益的影响, 标准化公式为
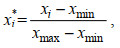 (2)
(2)其中, xi*表示第i个数标准化后的值; xi表示第i个数的原始值; xmax表示最大值; xmin表示最小值; 标准化处理之后的数值范围在0~1之间。养殖收益可表示为
 (3)
(3)其中, 产量用扇贝壳高计算得到的湿重(单位: kg/m2)表示; 为了剔除价格变动因素, 文中使用可比价格计算养殖收益, 即直接用产量乘某一年的不变价格计算(郭越等, 2012)。不变价格是以同类产品某年的平均价格作为固定价格, 用于计算各年的价值, 消除各时期价格变动的影响, 保证前后时期之间、地区之间、计划与实际之间指标的可比性(江西省发展和改革委员会课题组等, 2010), 因此, 文中以2014年为基期, 使用该年各规格虾夷扇贝的价格(表 1, 于佐安等, 2016)计算2007~2017年考虑肥满度后的养殖收益。实际生产中底播养殖虾夷扇贝的回捕率在40%左右(柳忠传等, 1995), 本文计算的是虾夷扇贝的理论最大收益并未考虑回捕率。需要说明的是, 本文主要是为了探讨扇贝的养殖容量和分区, 所计算的养殖收益只是为了统一考虑养殖产量和养殖品质而便于比较的综合指标, 并不是实际养殖收益。
表 1 虾夷扇贝规格及市场价格 Tab. 1 The shell length and cost of Yesso scallop
| 类别 | 规格(以壳高表示)/cm | 价格 |
| 苗种 | 3~4 | 0.15元/只 |
| 产品 | 6~8 | 12.00元/kg |
| > 8 | 20.00元/kg | |
| 注: 引自于佐安等(2016) | ||
通过计算各放苗密度下的养殖收益后, 选取各网格点的最大养殖收益所对应的放苗密度为最适放苗密度, 虾夷扇贝总干重为对应网格点的养殖容量。
1.4 养殖区区划养殖区用养殖收益划分。考虑扇贝肥满度后的最大养殖收益为0.00元/m2的海域直接认定为不宜底播养殖区, 其余海域对考虑肥满度后的最大养殖收益按年用Min-max标准化方法[公式(2)]做处理。养殖收益的年际差异能反映出各网格点的养殖风险, 各网格点考虑扇贝肥满度后的养殖收益的距平值取绝对值后用Min-max标准化方法[公式(2)]做处理。标准化处理后的考虑扇贝肥满度的养殖收益及养殖收益的距平值按照模型网格点使用k-means (Krishna et al, 1999)做聚类分析, 分类数用肘部法(Marutho et al, 2018)选取。
2 结果与分析 2.1 肥满度肥满度的时空差异明显。以放苗密度15 ind/m2为例, 长海海域多年平均肥满度范围在0.26~ 0.48 g/cm3之间, 空间上呈现由獐子岛附近海域至长海海域东北部的高值条带区向两侧降低的趋势(图 2)。除2007年外, 长海海域西北部的肥满度均最低。肥满度小于0.3 g/cm3的范围2010年最大, 2014年最小。研究区西北部的肥满度低主要受底层水温的限制(Nan et al, 2022), 在适宜扇贝生长的时段, 壳长和软体部干重均增长较快, 但是当底层水温限制扇贝摄食的时段, 扇贝只能通过消耗快速增长期存储在软体部干重中的能量来维持自身生存(Kooijman, 2010), 使得软体部干重降低, 肥满度偏小。长海海域南部的虾夷扇贝受饵料的限制壳长和软体部干重均生长缓慢(Nan et al, 2022), 虾夷扇贝的肥满度相对西北部较高, 只有在确定扇贝的大小时的肥满度才能说明扇贝的品质。
 |
| 图 2 2007~2017年肥满度的时空分布 Fig. 2 Spatio-temporal distribution of the condition index from 2007 to 2017 |
考虑肥满度后的最大养殖收益是各网格点在各放苗密度下养殖收益中的最大值, 多年区域平均值范围在0.00~9.77元/m2之间(图 3)。空间上, 2007~2017年考虑肥满度后的最大养殖收益的高值均位于獐子岛与小长山岛之间, 并向长海海域东北部延伸; 长海海域南部的养殖收益均最低。时间上, 考虑肥满度后的最大养殖收益2010年明显为多年最低, 区域平均为1.36元/m2; 2014年年为多年最大, 区域平均为4.74元/m2。
 |
| 图 3 2007~2017年考虑扇贝肥满度后的最大养殖收益的时空分布 Fig. 3 Spatio-temporal distribution of total profit considering the condition index from 2007 to 2017 |
考虑肥满度后的最大养殖收益所对应的放苗密度为对应海域的最适放苗密度。2007~2017年长海海域考虑肥满度后的底播养殖虾夷扇贝最大养殖收益的多年平均值所对应的最适放苗密度存在明显时空差异(图 4), 最适放苗密度范围在3~17 ind./m2之间, 区域平均为12 ind./m2。空间上, 长海海域西北部年际差异最大, 最适放苗密度范围在7~34 ind./m2之间; 研究区南部最适放苗密度最低, 大部分海区低于3 ind./m2, 主要由于该海域底层饵料浓度低所致(Nan et al, 2022), 这也说明该海区不适宜开展虾夷扇贝底播养殖。时间上, 2010年最适放苗密度为多年最大, 长海海域西北部达到34 ind./m2。
 |
| 图 4 2007~2017年最大养殖收益所对应的最适放苗密度的时空分布 Fig. 4 Spatio-temporal distribution of scallop density from 2007 to 2017 |
2007~2017年长海海域多年平均底播养殖虾夷扇贝的养殖容量范围为2.99~67.00 g/m2, 平均值为36.57 g/m2, 广鹿岛以南海域到獐子岛南部, 再经过海洋岛直至研究区东北部的条带区养殖容量最大, 南部边缘区最小(图 5)。养殖容量的年际差异明显, 2010年区域平均最小为25.57 g/m2, 研究区西北部和南部养殖容量低于30 g/m2, 仅獐子岛和海洋岛附近海域高于40 g/m2; 2009年区域平均最大为42.17 g/m2 (表 2), 獐子岛与大长山岛之间向研究区东北部延伸海域养殖收益均高于60 g/m2, 使得区域平均较高。虽然研究区西北部部分海域底播养殖虾夷扇贝放苗密度可达到34 ind./m2, 但养殖容量却低于30 g/m2, 说明底播养殖虾夷扇贝的养殖容量受养殖环境限制, 并不随放苗密度的增加而增大。
 |
| 图 5 2007~2017年虾夷扇贝养殖容量的时空分布 Fig. 5 Spatio-temporal distribution of production-carrying capacity from 2007 to 2017 |
表 2 2007~2017养殖容量与最适放苗密度的比较 Tab. 2 Comparison in production-carrying capacity and scallop density from 2007 to 2017
| 年度 | 养殖容量/(g/m2) | 最适放苗密度/(ind./m2) | |||
| 范围 | 平均值±标准误 | 范围 | 平均值±标准误 | ||
| 2007 | 2.65~75.69 | 39.17±17.47 | 3~15 | 11±4 | |
| 2008 | 2.65~66.94 | 37.03±18.41 | 3~20 | 12±4 | |
| 2009 | 2.95~83.48 | 42.17±20.06 | 3~16 | 12±4 | |
| 2010 | 2.95~59.45 | 25.57±12.14 | 3~34 | 11±5 | |
| 2011 | 2.95~72.95 | 36.50±19.40 | 3~15 | 11±4 | |
| 2012 | 2.95~64.66 | 36.79±17.04 | 3~22 | 12±4 | |
| 2013 | 2.95~67.00 | 34.52±16.35 | 3~21 | 12±4 | |
| 2014 | 3.23~84.66 | 37.95±17.21 | 3~17 | 11±3 | |
| 2015 | 3.05~79.14 | 39.40±19.59 | 3~16 | 12±3 | |
| 2016 | 3.23~75.97 | 37.21±17.40 | 3~15 | 11±3 | |
| 2017 | 2.93~81.56 | 36.50±17.90 | 3~15 | 12±4 | |
| 多年平均 | 2.99~67.00 | 36.57±17.56 | 3~17 | 12±4 | |
| 注: 数据形式为平均值±标准误, 最适放苗密度做了取整处理 | |||||
对2007~2017年标准化处理后的考虑虾夷扇贝肥满度的养殖收益按网格点使用k-means (Krishna et al, 1999)做聚类分析, 分类数由肘部法(Marutho et al, 2018)确定为4。按聚类结果将养殖区划分为4类。研究区南部大部分海域为不适宜底播养殖区(图 6, 紫色区域), 该区域主要由于饵料的匮乏使得底播养殖虾夷扇贝生长缓慢(Nan et al, 2022)。广鹿岛及小长山岛西北部海域为低收益区(图 6, 绿色区域), 该区域主要是因为底层水温的季节差异大所致, 冬季温度过低而夏季温度过高, 使得一年中适宜生长的时间较短(Nan et al, 2022), 虽然该区域饵料丰富, 在底层水温和饵料浓度完全匹配的阶段虾夷扇贝生长较快, 但是, 在温度限制摄食的时段内只能通过消耗储存在软体部干重中的能量来维持基本机能的运转, 从而使得虾夷扇贝的壳长较大, 但是肥满度不够; 研究区中部靠近不适宜底播区的条带区域受饵料限制同样为低收益区。研究区中部与低收益底播区相接的区域以及研究区西北部大长山岛附近海域为中收益区(图 6, 黄色区域)。从獐子岛和长山岛之间的海域向东北部的条带区域为高收益区(图 6, 红色区域)主要是由于该区域温度限制相对较少, 饵料相对充足, 也就是说该区域温度和饵料匹配的天数明显高于其他区域, 从而使得底播养殖虾夷扇贝的收益较高, 适宜底播养殖虾夷扇贝的发展。
 |
| 图 6 底播养殖虾夷扇贝养殖区区划图 Fig. 6 The planning of bottom culture area for Yesso scallop |
长海海域底播养殖虾夷扇贝在未考虑肥满度时的最大养殖收益在空间上均呈现出北部浅水高、南部海盆低的分布特征(图 7), 多年平均最大养殖收益的范围在0.51~14.91元/m2之间, 平均值为6.21元/m2。未考虑肥满度时的最大养殖收益高值位于大长山岛北部, 低值位于长海海域南部。2010年为多年区域平均最低, 养殖收益为3.69元/m2; 2015年为多年区域平均最高, 养殖收益为8.19元/m2 (表 3)。
 |
| 图 7 2007~2017年未考虑肥满度的最大养殖收益的时空分布 Fig. 7 Spatio-temporal distribution of total profit considering no condition index from 2007 to 2017 |
表 3 2007~2017是否考虑肥满度对养殖收益的影响 Tab. 3 Comparison in total profit with/without consideration of condition index from 2007 to 2017
| 年度 | 未考虑肥满度的养殖收益/(元/m2) | 考虑肥满度后的养殖收益/(元/m2) | |||
| 范围 | 平均值±标准误 | 范围 | 平均值±标准误 | ||
| 2007 | 0.51~16.05 | 6.57±4.30 | 0.02~11.44 | 4.13±2.68 | |
| 2008 | 0.51~12.50 | 4.69±2.68 | 0.02~ 7.37 | 2.72±1.79 | |
| 2009 | 0.51~14.77 | 5.22±3.08 | 0.01~11.59 | 3.34±2.10 | |
| 2010 | 0.51~ 7.97 | 3.69±2.07 | 0.00~ 4.38 | 1.36±0.87 | |
| 2011 | 0.51~10.67 | 4.58±2.66 | 0.01~ 6.00 | 2.60±1.56 | |
| 2012 | 0.51~13.65 | 4.92±2.88 | 0.01~ 7.91 | 2.73±1.74 | |
| 2013 | 0.51~14.97 | 5.15±3.53 | 0.01~ 9.96 | 2.65±1.97 | |
| 2014 | 0.51~17.35 | 7.80±5.00 | 0.00~13.84 | 4.74±3.14 | |
| 2015 | 0.51~18.67 | 8.19±5.57 | 0.00~12.96 | 4.35±3.38 | |
| 2016 | 0.51~18.00 | 6.96±4.89 | 0.00~13.21 | 3.79±3.27 | |
| 2017 | 0.51~19.25 | 8.08±5.72 | 0.00~13.69 | 4.09±3.36 | |
| 多年平均 | 0.51~14.91 | 6.21±4.21 | 0.00~ 9.77 | 3.32±2.28 | |
| 注: 数据形式为平均值±标准误 | |||||
考虑扇贝肥满度后的最大养殖收益(图 3)与不考虑扇贝肥满度的最大养殖收益(图 7)相比(表 3), 研究区西北部明显偏低, 主要是因为研究区北部受底层水温的限制, 适宜虾夷扇贝生长的时间较短(Nan et al, 2022)。由于不适宜生长天数较长, 虾夷扇贝通过消耗累积到软体部中的能量维持自身生存, 从而导致底播养殖虾夷扇贝的肥满度不高。可以看出, 考虑扇贝肥满度后的最大养殖收益的时空分布更加合理, 可作为研究区规划的依据。
3.2 养殖区区划的合理性及对现有底播养殖区的建议基于虾夷扇贝底播养殖规划的各养殖区(图 6), 求取各养殖区的养殖容量、最适放苗量和最大养殖收益的多年平均值作为指导养殖生产活动的参考值(表 4)。长海海域西北部的低收益养殖区参考放苗量为13 ind./m2; 研究区中部与不宜养殖区相接的低收益养殖区参考放苗量为15 ind./m2; 位于研究区西北部和研究区中部与低收益养殖区和高收益养殖区相接的中收益养殖区的扇贝参考放苗量分别为12和13 ind./m2; 高收益养殖区参考放苗量为12 ind./m2。本文区域平均放苗密度为12~15 ind./m2, 与Helm (2005)给出的壳高3 cm左右底播养殖虾夷扇贝的放苗密度的参考范围10~20 ind./m2相近。由于各养殖区海域较大, 表 4中的参考值只能作为达到平均养殖收益的参考, 而在实际小范围底播养殖活动中, 虾夷扇贝的放苗量应按照图 4实施更为合理。在后续工作中需要按照具体养殖区域给予具体的养殖放苗量范围。对比参考放苗密度和参考养殖收益(表 4)说明在实际虾夷扇贝养殖过程中只是盲目的增加养殖放苗量并不能达到增产增收的效果。
表 4 不同养殖区参考放苗密度及参考收益统计 Tab. 4 Reference scallop density and reference total profit in each aquaculture area
| 养殖区 | 参考放苗密度/(ind/m2) | 参考养殖收益/(元/m2) | ||||
| 平均值±标准误 | 范围 | 平均值±标准误 | 范围 | |||
| 高收益区 | 12±3 | 6~32 | 7.56±3.48 | 0.51~15.90 | ||
| 中收益区 | 北部 | 12±4 | 7~22 | 4.28±2.81 | 0.35~11.85 | |
| 南部 | 13±4 | 6~15 | 4.78±1.78 | 0.45~11.20 | ||
| 低收益区 | 北部 | 13±5 | 7~34 | 2.93±2.23 | 0.04~11.02 | |
| 南部 | 15±2 | 5~17 | 2.49±0.95 | 0.61~ 6.24 | ||
| 不宜底播区 | — | — | — | — | ||
| 注: 数据形式为平均值±标准误, 参考放苗密度的所有数值均做了取整处理; —表示无数据 | ||||||
长海海域现有底播养殖区(图 1)基本分布在底播养殖虾夷扇贝高、中收益养殖区范围内(图 6), 虾夷扇贝底播养殖区向长海海域南部海域的拓展需要谨慎, 因为该海域受到饵料限制, 底播养殖虾夷扇贝的生长受到影响(Nan et al, 2022)。
4 结论基于虾夷扇贝底播养殖生态系统模型在考虑扇贝肥满度的情况下探究底播养殖虾夷扇贝的养殖收益, 并评估养殖容量。经研究得到如下结论: (1) 考虑肥满度后的底播养殖虾夷扇贝养殖收益的空间分布更合理。(2) 底播养殖虾夷扇贝的养殖容量及最适放苗密度的空间差异明显。(3) 长海海域底播养殖区可划分为4类, 高收益区位于獐子岛和小长山岛之间并向研究区东北部延伸的条带海域, 适宜底播养殖的发展, 参考放苗量为12 ind./m2; 中收益区位于研究区西北部和中部均与低收益区和高收益区相接, 参考放苗量分别为13和12 ind./m2; 研究区西北部由于受温度限制产品肥满度不足, 属于低收益区, 参考放苗量为13 ind./m2; 研究区中部与不宜养殖区相接的条带区域受饵料限制同样属于低收益区, 参考放苗量为15 ind./m2; 研究区南部海域不适宜底播养殖。(4) 长海海域现有虾夷扇贝底播养殖区基本分布在底播养殖虾夷扇贝高、中收益养殖区范围内, 向长海海域南部的拓展需要谨慎。
参考文献
于佐安, 谭克非, 张明, 等, 2016. 筏式虾夷扇贝养成期不同密度生长与经济效益分析. 水产学报, 40(10): 1624-1633 |
方建光, 匡世焕, 孙慧玲, 等, 1996b. 桑沟湾栉孔扇贝养殖容量的研究. 海洋水产研究, 17(2): 18-31 |
方建光, 孙慧玲, 匡世焕, 等, 1996a. 桑沟湾海带养殖容量的研究. 海洋水产研究, 17(2): 7-17 |
卢振彬, 杜琦, 许翠娅, 等, 2005. 福建泉州湾贝类养殖容量评估. 热带海洋学报, 24(4): 22-29 DOI:10.3969/j.issn.1009-5470.2005.04.004 |
卢振彬, 杜琦, 阮金山, 等, 2000. 大港湾贝类养殖容量的评价. 福建水产, (1): 1-6 |
史洁, 魏皓, 赵亮, 等, 2010a. 桑沟湾多元养殖生态模型研究: Ⅰ养殖生态模型的建立和参数敏感性分析. 渔业科学进展, 31(4): 26-35 |
史洁, 魏皓, 赵亮, 等, 2010b. 桑沟湾多元养殖生态模型研究: Ⅱ生态环境模拟与生源要素循环. 渔业科学进展, 31(4): 36-42 |
史洁, 魏皓, 赵亮, 等, 2010c. 桑沟湾多元养殖生态模型研究: Ⅲ海带养殖容量的数值研究. 渔业科学进展, 31(4): 43-52 |
朱明远, 张学雷, 汤庭耀, 等, 2002. 应用生态模型研究近海贝类养殖的可持续发展. 海洋科学进展, 20(4): 34-42 DOI:10.3969/j.issn.1671-6647.2002.04.006 |
刘庆余, 1993. 紫贻贝养殖的环境容纳量问题. 水产科学, 12(9): 15-17 |
刘学海, 王宗灵, 张明亮, 等, 2015. 基于生态模型估算胶州湾菲律宾蛤仔养殖容量. 水产科学, 34(12): 733-740 |
刘剑昭, 李德尚, 董双林, 2000. 关于水产养殖容量的研究. 海洋科学, 24(10): 28-29, 13 DOI:10.3969/j.issn.1000-3096.2000.10.011 |
江西省发展和改革委员会课题组, 江西省价格理论研究所课题组., 2010. GDP核算中的价格问题与对策研究. 价格月刊, (5): 3-11 DOI:10.3969/j.issn.1006-2025.2010.05.001 |
麦康森, 2016. 中国的水产养殖、饲料原料与世界渔业资源. 饲料与畜牧, (6): 17-19 |
杜琦, 卢振彬, 戴泉水, 等, 2000. 同安湾贝类的养殖容量. 上海水产大学学报, 9(1): 21-26 |
李庆彪, 1990. 养殖扇贝的大量死亡与环境容纳量. 国外水产, (2): 9-11 |
李洪波, 梁玉波, 袁秀堂, 2012. 辽宁长海浮筏养殖对聚球藻蓝细菌分布的影响. 海洋学报, 34(5): 221-225 |
杨红生, 张福绥, 1999. 浅海筏式养殖系统贝类养殖容量研究进展. 水产学报, 23(1): 84-90 |
宋广军, 张雪, 宋伦, 等, 2013. 鸭绿江口浅海海域菲律宾蛤仔养殖容量估算. 水产科学, 32(1): 36-40 DOI:10.3969/j.issn.1003-1111.2013.01.007 |
陈家勇, 2019. 抓住机遇加快推进水产养殖业绿色发展. 中国渔业质量与标准, 9(4): 1-4 DOI:10.3969/j.issn.2095-1833.2019.04.001 |
柳忠传, 张贤励, 王尊清, 1995. 虾夷扇贝底播增殖技术. 海洋科学, 19(1): 14-16 |
贾后磊, 舒廷飞, 温琰茂, 2003. 水产养殖容量的研究. 水产科技情报, 30(1): 16-21 DOI:10.3969/j.issn.1001-1994.2003.01.003 |
高振锟, 2016. 环境胁迫对虾夷扇贝生理、免疫指标及行为学特性的影响[D]. 上海: 上海海洋大学: 8-43.
|
郭越, 宋维玲, 2012. 海洋生产总值可比价增速计算方法的思考. 海洋经济, 2(2): 40-43 |
唐启升, 1996. 关于容纳量及其研究. 海洋水产研究, 17(2): 1-6 |
董双林, 李德尚, 潘克厚, 1998. 论海水养殖的养殖容量. 青岛海洋大学学报, 28(2): 253-258 |
黎辉, 何慧, 金启增, 1999. 大亚湾三种经济贝类的营养生态位及潜在产量评估. 热带海洋, 18(4): 53-60 |
魏皓, 赵亮, 原野, 等, 2010. 桑沟湾水动力特征及其对养殖容量影响的研究——观测与模型. 渔业科学进展, 31(4): 65-71 DOI:10.3969/j.issn.1000-7075.2010.04.009 |
BACHER C, DUARTE P, FERREIRA J G, et al, 1997. Assessment and comparison of the Marennes-Oléron Bay (France) and Carlingford Lough (Ireland) carrying capacity with ecosystem models. Aquatic Ecology, 31(4): 379-394 DOI:10.1023/A:1009925228308 |
CARVER C E A, MALLET A L, 1990. Estimating the carrying capacity of a coastal inlet for mussel culture. Aquaculture, 88(1): 39-53 DOI:10.1016/0044-8486(90)90317-G |
CROSBY M P, GALE L D, 1990. A review of bivalve condition index methodologies with a suggested standard method. Journal of Shellfish Research, 9(1): 233-237 |
FAO, 2020. The State of World Fisheries and Aquaculture 2020. Sustainability in Action [M]. Rome, Italy: Food and Agriculture Organization of the United Nations: 1-21.
|
FILGUEIRA R, GRANT J, STRAND Ø, 2014. Implementation of marine spatial planning in shellfish aquaculture management: modeling studies in a Norwegian fjord. Ecological Applications, 24(4): 832-843 DOI:10.1890/13-0479.1 |
GUYONDET T, ROY S, KOUTITONSKY V G, et al, 2010. Integrating multiple spatial scales in the carrying capacity assessment of a coastal ecosystem for bivalve aquaculture. Journal of Sea Research, 64(3): 341-359 DOI:10.1016/j.seares.2010.05.003 |
HELM M, 2005. Cultured aquatic species information programme. Mizuhopecten yessoensis Masuda 1963 [PECTINIDAE] [N/OL]. (2006-06-12)[2022-03-01]. https://www.fao.org/fishery/en/culturedspecies/patinopecten_yessoensis/en.
|
JIANG W W, LIN F, DU M R, et al, 2020. Simulation of Yesso scallop, Patinopecten yessoensis, growth with a dynamic energy budget (DEB) model in the mariculture area of Zhangzidao Island. Aquaculture International, 28(1): 59-71 DOI:10.1007/s10499-019-00447-6 |
KOOIJMAN S A L M, 2010. Dynamic Energy Budget Theory for Metabolic Organisation. Cambridge, UK: Cambridge University Press, 1-284
|
KRISHNA K, MURTY M N, 1999. Genetic K-means algorithm. IEEE Transactions on Systems, Man, and cybernetics, Part B (Cybernetics), 29(3): 433-439 DOI:10.1109/3477.764879 |
MARUTHO D, HANDAKA S H, WIJAYA E, et al, 2018. The determination of cluster number at K-mean using elbow method and purity evaluation on headline news [C] // 2018 International Seminar on Application for Technology of Information and Communication. Semarang, Indonesia: IEEE: 533-538.
|
NAN X L, WEI H, ZHANG H Y, et al, 2022. Spatial difference in net growth rate of Yesso scallop Patinopecten yessoensis revealed by an aquaculture ecosystem model. Journal of Oceanology and Limnology, 40(1): 373-387 DOI:10.1007/s00343-021-0423-4 |
NAYLOR R L, HARDY R W, BUSCHMANN A H, et al, 2021. A 20-year retrospective review of global aquaculture. Nature, 591(7851): 551-563 |
YUAN X T, ZHANG M J, LIANG Y B, et al, 2010. Self-pollutant loading from a suspension aquaculture system of Japanese scallop (Patinopecten yessoensis) in the Changhai sea area, Northern Yellow Sea of China. Aquaculture, 304(1/2/3/4): 79-87 DOI:10.1016/j.aquaculture.2010.03.026 |
ZHANG X L, ZHU M Y, LI R X, et al, 2006. Density-dependent mortality of the scallop Chlamys farreri (Jones & Preston) in grow-out culture. Aquaculture Research, 37(8): 842-844 |
ZHAO Y X, ZHANG J H, LIN F, et al, 2019. An ecosystem model for estimating shellfish production carrying capacity in bottom culture systems. Ecological Modelling, 393: 1-11 |