 2022, Vol. 53
2022, Vol. 53

浙江南部近海春季鱼类群落结构特征及其年际变化

http://dx.doi.org/10.11693/hyhz20220300080
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 王寅, 赵静, 高春霞, 王士聪, 叶深. 2022.
- WANG Yin, ZHAO Jing, GAO Chun-Xia, WANG Shi-Cong, YE Shen. 2022.
- 浙江南部近海春季鱼类群落结构特征及其年际变化
- STRUCTURE AND INTERANNUAL VARIATION OF FISH COMMUNITIES IN THE OFFSHORE WATERS OF SOUTHERN ZHEJIANG PROVINCE IN SPRING
- 海洋与湖沼, 53(6): 1455-1466
- Oceanologia et Limnologia Sinica, 53(6): 1455-1466.
- http://dx.doi.org/10.11693/hyhz20220300080
文章历史
-
收稿日期:2022-03-31
收修改稿日期:2022-06-10
引用本文
王寅, 赵静, 高春霞, 王士聪, 叶深. 2022. 浙江南部近海春季鱼类群落结构特征及其年际变化[J]. 海洋与湖沼, 53(6): 1455-1466

WANG Yin, ZHAO Jing, GAO Chun-Xia, WANG Shi-Cong, YE Shen. 2022. STRUCTURE AND INTERANNUAL VARIATION OF FISH COMMUNITIES IN THE OFFSHORE WATERS OF SOUTHERN ZHEJIANG PROVINCE IN SPRING[J]. Oceanologia et Limnologia Sinica, 53(6): 1455-1466.
浙江南部近海春季鱼类群落结构特征及其年际变化
1. 上海海洋大学海洋科学学院 上海 201306;
2. 国家远洋渔业工程技术研究中心 上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室 上海 201306;
4. 农业农村部大洋渔业资源环境科学观测实验站 上海 201306;
5. 浙江省海洋水产养殖研究所 浙江温州 325005
2. 国家远洋渔业工程技术研究中心 上海 201306;
3. 大洋渔业资源可持续开发教育部重点实验室 上海 201306;
4. 农业农村部大洋渔业资源环境科学观测实验站 上海 201306;
5. 浙江省海洋水产养殖研究所 浙江温州 325005
摘要:根据2016年和2020年5月在浙江南部近海采集的渔业资源调查资料, 采用相对重要性指数、Bray-Curtis聚类和相似性百分比SIMPER分析、生物量谱及ABC曲线等方法, 分析该水域春季鱼类群落结构特征及其稳定性的年际变化。结果显示: (1) 2016年春季共捕获鱼类65种, 优势种为带鱼Trichiurus lepturus、七星底灯鱼Benthosema pterotum和鳀Engraulis japonicus, 而2020年春季共捕获鱼类30种, 优势种为七星底灯鱼和带鱼; (2) 通过聚类分析发现, 2016和2020年春季鱼类群落空间格局相似, 均可划分为南、北群落; (3) 通过SIMPER分析, 2016年春季南部群落典型种为二长棘鲷Evynnis cardinalis、龙头鱼Harpadon nehereus和鳀, 北部群落典型种为龙头鱼, 群落间的分歧种为带鱼; 2020年南部群落典型种为七星底灯鱼, 北部群落典型种为镰鲳Pampus echinogaster和小黄鱼Larimichthys polyactis, 群落间的分歧种为带鱼、镰鲳和小黄鱼; (4) 基于生物量谱分析, 2016年和2020年春季鱼类标准生物量谱的斜率均小于–1, 表明生物量均随个体平均体质量的增加而减少; (5) ABC曲线结果表明, 2016年春季鱼类群落处于中等干扰状态(W= –0.22), 而2020年春季鱼类群落则处于严重干扰状态(W=–0.09); (6) 经RDA排序分析, 2016年和2020年春季影响鱼类群落分布的主要环境因子均为表层水温、盐度和水深。综上所述, 浙江南部近海的鱼类年际变化为鱼类种类显著下降, 群落特征种更替明显, 群落受外界干扰影响加剧。
关键词:浙江南部近海 春季 鱼类群落 生物量谱 ABC曲线
STRUCTURE AND INTERANNUAL VARIATION OF FISH COMMUNITIES IN THE OFFSHORE WATERS OF SOUTHERN ZHEJIANG PROVINCE IN SPRING
1. College of Marine Sciences of Shanghai Ocean University, Shanghai 201306, China;
2. National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai 201306, China;
3. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Shanghai Ocean University, Shanghai 201306, China;
4. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
5. Zhejiang Mariculture Research Institute, Wenzhou 325005, China
2. National Engineering Research Center for Oceanic Fisheries, Shanghai Ocean University, Shanghai 201306, China;
3. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Shanghai Ocean University, Shanghai 201306, China;
4. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
5. Zhejiang Mariculture Research Institute, Wenzhou 325005, China
Abstract: Based on the fishery survey data collected in the offshore waters of southern Zhejiang Province in May 2016 and May 2020, the index of relative importance, Bray-Curtis cluster analysis, similarity percentage (SIPMER) analysis, biomass size spectrum, and ABC curves were selected to determine the fish community structure over time. A total of 65 fish species were captured in the spring of 2016, and the dominant species were Trichiurus lepturus, Benthosema pterotum and Engraulis japonicus, while 30 fish species of fish were obtained in the spring of 2020, and the dominant species were B. pterotum and T. lepturus. The cluster analysis revealed similar patterns of fish community in 2016 and 2020, from which the northern and southern communities could be divided. As SIMPER analysis showed, in spring 2016, typical species of the southern community were Parargyrops edita, Harpadon nehereus, and E. japonicus, while in the northern community, H. nehereus was typical; the divergent species between the two communities was T. lepturus. In spring 2020, the typical species of the southern community was B. pterotum, while those in the northern community were Pampus echinogaster and Larimichthys polyactis; the divergent species between communities were T. lepturus, P. echinogaster, and L. polyactis. In addition, as the biomass spectra show, the slopes of normalized biomass spectra were below –1, and the biomass decreased with the increase of average individual weight. The fish community was in a state of moderate disturbance (W= –0.22) in spring of 2016 but seriously disturbed (W= –0.09) in spring 2020 as revealed on the ABC curves. Moreover, surface water temperature, salinity, and water depth were main impact factors on fish community in the two spring seasons. Therefore, the interannual variation of fish community in the offshore waters of southern Zhejiang Province was manifested by significant decline in fish species, obvious change of community characteristic species, and intensification of external disturbance to the local bio-communities.
Key words:
offshore waters of southern Zhejiang Province spring fish community biomass spectra ABC curve
近年来, 过度捕捞、资源的不合理利用以及生态环境恶化等因素造成了近海渔业资源的不断衰退, 渔业群落结构呈现小型种类和小个体增多、大个体减少的变化趋势(程家骅等, 2006; 刘勇等, 2015), 水域内渔业群落结构稳定性受到不同程度的干扰。目前对群落结构稳态的量化手段有ABC曲线和生物量谱法(Rice, 2003; Rochet et al, 2003; Shin et al, 2005)。其中ABC曲线(Abundance-Biomass Comparison Curves)通过分析外界干扰对生物群落的影响来确定生物群落对干扰产生预期的反应(李圣法, 2008), Wijeyaratne等(2018)利用ABC曲线预测迪亚弯那湿地系统的环境压力水平; 刘前等(2021)利用ABC曲线发现湛江港海域游泳动物群落受到外界干扰具有季节差异性。生物量谱(Biomass size spectrum, BSS)是由Sheldon等(1972)提出的粒径谱逐步发展而来, 反映水生生态系统的群落结构个体组成及稳定性状况, Magnussen (2002)利用生物量谱发现法罗群岛浅滩区域占主导地位的物种由大个体鱼类逐步向小个体鱼类转移; 徐姗楠等(2020)利用生物量粒径谱方法确定大亚湾各海区不同年份鱼类群落的受干扰状态。
浙江南部近海地处27°~29°N之间的东海近岸, 属暖温带海区。此处为高盐度的台湾暖流及两股低盐度海流交汇处, 营养盐和饵料生物丰富, 为渔业生物栖息、生长及繁殖提供了适宜的环境条件(宋海棠等, 1995)。近年来, 有报道该水域的鱼类群落结构及稳定性发生较大变化(杜晓雪等, 2018; 杨柯迩等, 2022), 水域内出现生物多样性降低、渔获物趋于小型化和低龄化以及营养级下降等诸多问题(张琥顺等, 2021), 因此对群落结构的变动开展研究具有重要的现实意义。目前对于浙江南部近海群落结构的研究主要侧重于季节变化(杜晓雪等, 2018; 丁朋朋等, 2019; 戴小杰等, 2019; 杨柯迩等, 2022), 而对年份间变化特征的趋势性研究鲜有开展。本研究利用2016年和2020年春季的底拖网数据, 运用聚类和相似性分析、生物量谱和ABC曲线以及冗余分析等方法分析该海域春季鱼类群落结构、受干扰状况以及与环境因子的关系等, 旨在揭示浙江南部近海群落结构的演变趋势, 为该海域渔业资源保护和生态系统修复提供理论依据。
1 材料与方法 1.1 数据来源2016年5月和2020年5月在浙江南部近海(27°00′~29°00′N, 120°00′~123°00′E)开展底拖网渔业资源调查, 采样调查20个站点(图 1)。调查船为浙洞渔10109, 总吨位305 t, 主机马力540 hp, 为近海大型单船底拖网作业船。渔船作业时底拖网网口的平均扩张高度和宽度分别为7.5 m和40 m, 根据实际测算得到不同水深时网口扩张的高度为4 m, 网囊网目为2 cm。现场采样拖速控制在2.2~5.6 kn, 拖曳时间控制在1 h以内, 统一对渔获数据进行拖网时间(1 h)和拖网速度(3 kn)的标准化处理。调查及样品保存参照《海洋调查规范》(GB/T 12763-2007)开展, 种类鉴定参照《浙江鱼类志》和《中国海洋鱼类》。同步利用多功能水质分析仪(WTW-Multi 3430)采集物理环境数据, 包括表层水温、底层水温、盐度和pH。
 |
| 图 1 调查区域和采样点 Fig. 1 The area of investigation and the sampling site |
利用Pinkas等(1971)提出的相对重要性指数(index of relative importance, IRI)确定生物种类在群落中的重要性, 计算公式为
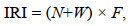 (1)
(1)式中, N为某一物种数量占总数量的百分比; W为某一物种生物量占总生物量的百分比; F为某一物种出现的站位数占总站位数的百分比。
本研究中, 优势种为IRI≥1 000, 重要种为1 000 > IRI≥100, 常见种为100 > IRI≥10, 少见种为IRI < 10 (程济生, 2005)。根据鱼类的适温类型可将鱼种分为暖水种、暖温种和冷水种(田明诚等, 1993)。
1.2.2 聚类和相似性分析利用PRIMER 6.0软件开展鱼类群落结构分析, 对鱼类的标准化生物量数据进行平方根转换, 随后计算Bray-Curtis相似性系数, 构建相似性矩阵; 利用CLUSTER等级聚类分析将鱼类群落进行聚类处理, 并根据聚类结果进行SIMPER分析, 确定不同鱼类对各群落组内相似性和组间相异性的平均贡献率(Clarke, 1993)。本研究将组内平均相似性贡献率大于4%的鱼类定为典型种, 组间平均相异性贡献率大于4%的鱼类定为分歧种。
1.2.3 生物量谱Platt等(1977)提出P-D模型是用来表示生物量谱的典型模型之一, 它以对数化生物量为纵坐标, 以对数化粒径为横坐标, 所得到的谱线称为非标准化生物量谱。本文采用生物量谱分析浙江南部近海鱼类生物量与平均体质量间的关系。首先选取数量占比超过0.01%的鱼类计算每种鱼类的平均体质量, 以10 g为单位将其分为若干区间, 计算各区间总生物量。然后以平均个体体质量除以区间间隔的对数化值为x轴, 以各区间总生物量对数化值为y轴, 线性回归后可得到标准化生物量谱(Magnussen, 2002)。当谱线斜率为–1时, 表示生物群落为稳定状态, 即生物量随体质量的增加均匀分布; 当斜率大于–1时, 生物量随体质量增加而增加; 当斜率小于–1时, 生物量随体质量增加而减少。
1.2.4 ABC曲线ABC曲线以生物进化r-选择和k-选择理论为基础, 反映不同干扰情况下群落的反应(Yemane et al, 2005)。未干扰状态下, 群落中以k-选择种类(即生长慢、性成熟晚的大个体种类)为主, 生物量优势度曲线会高于数量优势度曲线; 中等干扰状态下, 群落中k-选择种类数量将逐渐减少, r-选择种类(即生长快、性成熟早的小个体种类)的数量逐渐增多, 生物量和数量优势度曲线出现相交; 严重干扰状态下, 群落中以r-选择种类为主, 数量优势度曲线则高于生物量优势度曲线。本研究ABC曲线以调查站点的物种总数据为样本, W值表示生物量与数量的相对关系统计量(晏磊等, 2015), 利用PRIMER 6.0绘制ABC曲线, 计算公式如下:
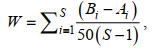 (2)
(2)式中, Ai和Bi分别为ABC曲线中种类序号i对应的数量和生物量的累积百分比, S为物种种类数。
1.2.5 冗余分析(redundancy analysis, RDA)本研究选取表层水温、底层水温、盐度、水深、pH作为解释变量, 标准化数量作为响应变量进行RDA排序, 通过Monter-Carlo置换检验筛选对鱼类群落有显著影响的环境变量, 以上操作在CANOCO 5软件中进行, 排序图采用物种-环境因子的双序图表示。在RDA排序图中, 环境因子用带有箭头的线段表示, 连线的长短表示鱼类群落分布和种类分布与该环境因子关系的相关性大小, 连线越长说明相关性越大, 反之越小; 物种与环境因子箭头之间的夹角的余弦值反映它们之间的相关性, 夹角越小, 相关性越大(Ter Braak, 1989)。在利用RDA分析前进行去趋势对应分析诊断, 若结果中梯度长度最大值小于3, 则选择RDA分析; 若大于4, 则选择典范对应分析; 若在3~4之间, 则两种分析方法皆可。本文对两个年份春季的鱼类生物量数据进行去趋势对应分析, 结果表明前4个轴最大值均小于3, 因此选用RDA分析。
2 结果 2.1 种类组成2016年春季共采集鱼类65种, 隶属于1纲13目43科58属, 均为硬骨鱼纲。其中, 鲈形目30种, 占种类总数的46.15%; 鲱形目、鲉形目、鳗鲡目、鲽形目均为5种, 各占种类总数的7.69%, 仙女鱼目4种, 占种类总数的6.15%; 鲭形目、灯笼鱼目、鳕形目、鲀形目均为2种, 各占种类总数的3.07%; 
2020年春季共采集鱼类30种, 隶属于2纲10目20科27属, 除尖头斜齿鲨(Scoliodon laticaudus)为软骨鱼纲外, 其余均为硬骨鱼纲。其中, 鲈形目16种, 占种类总数的53.33%; 鲱形目3种, 占种类总数的10.00%; 鲭形目、仙女鱼目、鲉形目均为2种, 各占种类总数的6.67%; 灯笼鱼目、
2016年春季暖水种鱼类39种, 占60.00%; 暖温种鱼类25种, 占38.46%; 冷水种鱼类仅1种, 占1.54%。2020年春季暖水种鱼类19种, 占63.33%; 暖温种鱼类10种, 占33.33%; 冷水种鱼类仅1种, 占3.34%。两年所调查到的冷水种鱼类均为黄
2016年春季调查渔获优势种3种、重要种9种、常见种15种、少见种38种, 其中带鱼相对重要性指数最高(IRI=8 317), 七星底灯鱼次之(IRI=2 671), 鳀为第三(IRI=1 245)。2020年春季调查渔获优势种2种、重要种7种、常见种5种、少见种16种, 其中七星底灯鱼优势度最高(IRI=9 159), 其次为带鱼(IRI=5 373)。
表 1 2016年和2020年春季主要鱼类相对重要性指数 Tab. 1 The relative importance indexes of major fish species in the spring of 2016 and 2020
| 种类 | 2016年 | 2020年 | |||||||
| N/% | W/% | F/% | IRI | N/% | W/% | F/% | IRI | ||
| 带鱼Trichiurus lepturus | 33.47 | 64.38 | 85 | 8 317 | 48.35 | 8.21 | 95 | 5 373 | |
| 七星底灯鱼Benthosema pterotum | 49.89 | 3.54 | 50 | 2 671 | 19.95 | 81.82 | 90 | 9 159 | |
| 鳀Engraulis japonicus | 3.65 | 10.99 | 85 | 1 245 | 1.92 | 2.87 | 35 | 168 | |
| 镰鲳Pampus echinogaster | 8.97 | 0.48 | 90 | 850 | |||||
| 蓝圆鲹Decapterus maruadsi | 2.18 | 4.13 | 90 | 567 | |||||
| 龙头鱼Harpadon nehereus | 0.70 | 3.46 | 95 | 395 | 2.72 | 0.11 | 95 | 269 | |
| 银姑鱼Pennahia argentata | 1.62 | 2.14 | 90 | 338 | |||||
| 小黄鱼Larimichthys polyactis | 0.44 | 2.15 | 80 | 208 | 5.97 | 0.25 | 50 | 311 | |
| 刺鲳Psenopsis anomala | 0.67 | 1.34 | 100 | 201 | 2.24 | 0.12 | 95 | 224 | |
| 黄鲫Setipinna taty | 2.65 | 0.30 | 70 | 206 | |||||
| 绿鳍鱼Chelidonichthys kumu | 0.96 | 0.70 | 100 | 166 | |||||
| 六丝钝尾虾虎鱼Amblychaeturichthys hexanema | 1.93 | 0.76 | 55 | 148 | |||||
| 二长棘鲷Evynnis cardinalis | 0.49 | 0.86 | 100 | 135 | |||||
| 日本鲭Scomber japonicus | 0.22 | 1.23 | 85 | 123 | |||||
| 细条天竺鲷Apogonichthys lineatus | 0.53 | 0.83 | 85 | 116 | |||||
图 2、图 3为浙江南部近海2016年和2020年春季底拖网鱼类生物量和站点聚类结果。2016年春季在39.6%的相似性水平上鱼类群落聚为3组, A组包括2个调查站位, B组包括13个调查站位, C组包括5个调查站位, 调查海域大致可划分为北部群落(B组)和南部群落(A组和C组)。2020年春季鱼在40.5%的相似性水平上类群落同样聚为2组, A组包括3个调查站位, B组包括16个调查站位, 调查海域可划分为北部群落(A组)和南部群落(B组)。
 |
| 图 2 2016年春季基于Bray-Curtis相似性系数的站位聚类分析 Fig. 2 Cluster dendrogram based on Bray-Curtis similarity of biomass in spring 2016 |
 |
| 图 3 2020年春季基于Bray-Curtis相似性系数的站位聚类分析 Fig. 3 Cluster dendrogram based on Bray-Curtis similarity of biomass in spring 2020 |
根据SIMPER结果(表 2), 2016年春季A组组内的平均相似性百分比为48.42%, 主要典型种有海鳗(Muraenesox cinereus)、六丝钝尾虾虎鱼、鳀、黄
表 2 2016年春季组内相似性和组间相异性贡献率超过4%的种类 Tab. 2 Species contributing above 4% of accumulative similarity within a group and dissimilarity between groups in spring 2016
| 种类 | 组内典型种贡献率/% | 组间分歧种贡献率/% | |||||
| A组 | B组 | C组 | A组 & B组 | A组 & C组 | B组 & C组 | ||
| 银姑鱼 | 5.62 | ||||||
| 刺鲳 | 6.57 | 4.71 | |||||
| 带鱼 | 30.25 | 10.42 | 22.43 | 6.73 | 21.60 | ||
| 二长棘鲷 | 4.05 | 12.84 | |||||
| 海鳗 | 15.96 | 7.58 | |||||
| 黄鮟鱇 | 7.53 | 4.96 | |||||
| 黄鲫 | 5.39 | 5.01 | |||||
| 孔虾虎鱼 | 7.11 | ||||||
| 拉氏狼牙虾虎鱼 | 7.12 | ||||||
| 六丝钝尾虾虎鱼 | 11.76 | 4.22 | 4.89 | 8.14 | |||
| 龙头鱼 | 10.04 | 10.11 | 5.04 | 4.20 | |||
| 绿鳍鱼 | 5.95 | ||||||
| 皮氏叫姑鱼 | 6.36 | 4.01 | |||||
| 七星底灯鱼 | 4.47 | ||||||
| 日本鲭 | 9.31 | 4.12 | |||||
| 鳀 | 11.29 | 10.13 | 4.59 | 7.25 | 7.66 | ||
| 细条天竺鲷 | 4.21 | ||||||
| 小黄鱼 | 5.74 | 4.40 | 4.07 | ||||
| 竹荚鱼 | 4.37 | ||||||
| 累积贡献率 | 77.45 | 66.47 | 71.15 | 41.35 | 48.00 | 37.81 | |
根据SIMPER结果(表 3), 2020年5月A组组内的平均相似性百分比为65.81%, 主要典型种有带鱼、镰鲳、小黄鱼、刺鲳等, 对组内平均相似性的贡献率达93.03%; B组组内的平均相似性百分比为47.57%, 主要典型种有带鱼、七星底灯鱼、镰鲳、刺鲳、多齿蛇鲻(Saurida tumbil)、蓝圆鲹、黄鲫等, 对组内平均相似性的贡献率达88.77%。A组和B组组间的平均相异性百分比为66.84%, 主要分歧种为带鱼、镰鲳、小黄鱼、七星底灯鱼、龙头鱼、刺鲳, 对组间相异性的贡献率为78.63%, 其中带鱼对组间相异性的贡献率为30.47%, 镰鲳为15.49%, 小黄鱼为14.04%。
表 3 2020年春季组内相似性和组间相异性贡献率超过4%的种类 Tab. 3 Species contributing above 4% of accumulative similarity within a group and dissimilarity between groups in spring 2020
| 种类 | 组内典型种贡献率/% | 组间分歧种贡献率/% | ||
| A组 | B组 | A组 & B组 | ||
| 七星底灯鱼 | 20.86 | 7.82 | ||
| 带鱼 | 51.76 | 35.47 | 30.47 | |
| 镰鲳 | 20.57 | 8.05 | 15.49 | |
| 蓝圆鲹 | 5.80 | |||
| 小黄鱼 | 15.79 | 14.04 | ||
| 龙头鱼 | 6.07 | |||
| 刺鲳 | 4.91 | 7.30 | 4.74 | |
| 黄鲫 | 5.35 | |||
| 多齿蛇鲻 | 5.94 | |||
| 累积贡献率 | 93.03 | 88.77 | 78.63 | |
图 4为标准化生物量谱的回归分析图, 2016年春季标准化生物量谱线性回归方程为y = –1.495 9x + 20.178 (R2=0.570 5); 2020年春季标准化生物量谱线性回归方程为y = –1.451 8 x + 21.606 (R2=0.582 8)。
 |
| 图 4 生物量谱的回归分析 Fig. 4 Regression analysis for biomass spectra |
图 5可以看出生物量随平均个体质量的变化, 2016年春季平均个体质量小于10 g的鱼类的总生物量占比7.31%; 10~20 g的鱼类的总生物量占比为71.38%, 其中带鱼占比高达64.68%; 20~50 g的鱼类的总生物量占比为18.32%; 50 g以上的鱼类仅占2.99%。2020年春季平均个体质量小于10 g的鱼类的总生物量占比23.23%, 较2016年占比有大幅上升, 其中七星底灯鱼占18.31%; 10~20 g的鱼类的总生物量占比为51.18%, 其中带鱼占比50.84%; 20~50 g的鱼类的总生物量占比为5.20%; 50 g以上的鱼类占比20.39%, 占比较2016年有所增加。
 |
| 图 5 各个平均个体质量的生物量比例 Fig. 5 Biomass ratio of average individual weight |
2016年5月鱼类群落的数量优势度曲线与生物量优势度曲线相交, 其W统计值为–0.022, 处于中等干扰状态。生物量百分比排在前5位的鱼类为带鱼、鳀、七星底灯鱼、龙头鱼、小黄鱼, 数量百分比排在前5位的鱼类为七星底灯鱼、带鱼、鳀、六丝钝尾虾虎鱼、银姑鱼。以上7种渔获中, 仅七星底灯鱼和六丝钝尾虾虎鱼数量百分比高于生物量百分比, 其中七星底灯鱼数量占比高出生物量占比46.39%。
2020年5月鱼类群落的数量优势度曲线在生物量优势度曲线上方, 其W统计值为–0.09, 处于严重干扰状态。生物量百分比排在前5位的鱼类为带鱼、七星底灯鱼、镰鲳、小黄鱼、龙头鱼, 数量百分比排在前5位的鱼类为七星底灯鱼、带鱼、蓝圆鲹、鳀、麦氏犀鳕(Bregmaceros mcclellandi)。以上8种渔获中, 七星底灯鱼、蓝圆鲹、鳀、麦氏犀鳕数量百分比高于生物量百分比, 其中七星底灯鱼数量占比高出生物量占比62.98%。综合来看, 浙江南部近海的鱼类群落稳定性受干扰加剧, 小个体鱼类占比增多, 在群落结构中占据绝对优势。
 |
| 图 6 ABC曲线 Fig. 6 The ABC curves |
根据RDA分析结果(图 7), 2016年春季5项环境因子共解释该海域36.9%的鱼类变异情况, 第一排序轴和第二排序轴分别解释了方差变化的14.64%和8.24%。蒙特卡罗置换检验表明, 第一排序轴(P=0.01)和所有排序轴(P=0.002)均差异显著(P < 0.05), 说明排序结果可靠。表层水温(P=0.002)、水深(P=0.01)和盐度(P=0.026)对鱼类群落分布影响显著(P < 0.05), 共解释了27.2%的物种变化信息。
 |
图 7 春季鱼类与环境因子的RDA排序 Fig. 7 The RDA ordination of fish species and environment factors in spring 注: 图中SST表示表层温度; SDT表示底层温度; Sal表示盐度; Dep表示水深; ○形表示南部群落; △形表示北部群落。数字代表鱼种: 1. 带鱼; 2. 鳀; 3. 七星底灯鱼; 4. 龙头鱼; 5. 小黄鱼; 6. 银姑鱼; 7. 刺鲳; 8. 日本鲭; 9. 黄鲫; 10. 皮氏叫姑鱼; 11. 海鳗; 12. 二长棘鲷; 13. 细条天竺鲷; 14. 小带鱼Eupleurogrammus muticus; 15. 六丝钝尾虾虎鱼; 16. 绿鳍鱼; 17. 黄; 18. 日本䲢Uranoscopus japonicus; 19. 前肛鳗Dysomma anguillare; 20. 发光鲷Acropoma japonicum; 21. 短鳄齿鱼Champsodon snyderi; 22. 麦氏犀鳕; 23. 鳓Ilisha elongate; 24. 大头银姑鱼Pennahia macrocephalus; 25. 大黄鱼Larimichthys crocea; 26. 虻 Erisphex pottii; 27. 蓝圆鲹; 28. 丁氏 Erisphex pottii; 27. 蓝圆鲹; 28. 丁氏 Johnius distinctus; 29. 竹荚鱼; 30. 长蛇鲻Saurida elongate; 31. 花斑蛇鲻Saurida undosquami; 32. 六指马鲅Polydactylus sextarius; 33. 短尾吻鳗Rhynchocymba sivicola; 34. 黄泽小沙丁鱼Sardinella lemuru; 35. 黑尾吻鳗Rhynchoconger ectenurus; 36. 拉氏狼牙虾虎鱼; 37. 多齿蛇鲻; 38. 断线舌鳎Cynoglossus interruptus; 39. 尖吻蛇鳗Ophichthus apicalis; 40. 宽体舌鳎Cynoglossus robustus; 41. 黑鳃兔头鲀Lagocephalus inermis; 42. 镰鲳; 43. 孔虾虎鱼; 44. 日本方头鱼Branchiostegus japonicus; 45. 五眼斑鲆Pseudorhombus pentophthalmus; 46. 栉鳞鳎Aseraggodes kaianus; 47. 中华栉孔虾虎鱼Ctenotrypauchen chinensis; 48. 黑潮新鼬鳚Neobythites sivicola; 49. 短尾大眼鲷Priacanthus macracanthus; 50. 无斑圆鲹Decapterus kurroides; 51. 丝背细鳞鲀Stephaolepis cirrhifer; 52. 条尾绯鲤Upeneus japonicus; 53. 六带拟鲈Parapercis sexfasciata; 54. 鹿斑鲾Secutor ruconius; 55. 舒氏海龙Syngnathus schlegeli; 56. 多棘腔吻鳕Coelorinchus multispinulosus; 57. 鳞首方头鲳Cubiceps squamiceps; 58. 印度小公鱼Stolephorus indicus; 59. 带纹条鳎Zebrias zebra; 60. 褐鲳蚰Sebastiscus marmoratu; 61. 背点棘赤刀鱼Acanthocepola limbate; 62. 印度棘赤刀鱼Acanthocepola indica; 63. 日本裸蜥鱼Lestrolepis japonica; 64. 丝鳍虎鲉Minous pusillus; 65. 翼红娘鱼Lepidotrigla alata; 66. 尖头斜齿鲨; 67. 黑姑鱼Atrobucca nibe; 68. 尖牙鲷Synagrops argyreus; 69. 真赤鲷Pagrus major Johnius distinctus; 29. 竹荚鱼; 30. 长蛇鲻Saurida elongate; 31. 花斑蛇鲻Saurida undosquami; 32. 六指马鲅Polydactylus sextarius; 33. 短尾吻鳗Rhynchocymba sivicola; 34. 黄泽小沙丁鱼Sardinella lemuru; 35. 黑尾吻鳗Rhynchoconger ectenurus; 36. 拉氏狼牙虾虎鱼; 37. 多齿蛇鲻; 38. 断线舌鳎Cynoglossus interruptus; 39. 尖吻蛇鳗Ophichthus apicalis; 40. 宽体舌鳎Cynoglossus robustus; 41. 黑鳃兔头鲀Lagocephalus inermis; 42. 镰鲳; 43. 孔虾虎鱼; 44. 日本方头鱼Branchiostegus japonicus; 45. 五眼斑鲆Pseudorhombus pentophthalmus; 46. 栉鳞鳎Aseraggodes kaianus; 47. 中华栉孔虾虎鱼Ctenotrypauchen chinensis; 48. 黑潮新鼬鳚Neobythites sivicola; 49. 短尾大眼鲷Priacanthus macracanthus; 50. 无斑圆鲹Decapterus kurroides; 51. 丝背细鳞鲀Stephaolepis cirrhifer; 52. 条尾绯鲤Upeneus japonicus; 53. 六带拟鲈Parapercis sexfasciata; 54. 鹿斑鲾Secutor ruconius; 55. 舒氏海龙Syngnathus schlegeli; 56. 多棘腔吻鳕Coelorinchus multispinulosus; 57. 鳞首方头鲳Cubiceps squamiceps; 58. 印度小公鱼Stolephorus indicus; 59. 带纹条鳎Zebrias zebra; 60. 褐鲳蚰Sebastiscus marmoratu; 61. 背点棘赤刀鱼Acanthocepola limbate; 62. 印度棘赤刀鱼Acanthocepola indica; 63. 日本裸蜥鱼Lestrolepis japonica; 64. 丝鳍虎鲉Minous pusillus; 65. 翼红娘鱼Lepidotrigla alata; 66. 尖头斜齿鲨; 67. 黑姑鱼Atrobucca nibe; 68. 尖牙鲷Synagrops argyreus; 69. 真赤鲷Pagrus major |
2020年春季4项环境因子可解释该海域34.5%的鱼类变异情况, 第一排序轴和第二排序轴特征值分别解释了方差变化的14.1%和9.07%。蒙特卡罗置换检验显示, 第一排序轴(P=0.016)和所有排序轴(P=0.004)均差异显著(P < 0.05), 表明环境因子与鱼类群落分布存在线性关系, 其中表层温度(P=0.024)、水深(P=0.026)和盐度(P=0.042)是影响鱼类群落分布的主要环境因子(P < 0.05), 共解释了28.4%的物种变化信息。
3 讨论 3.1 鱼类群落组成变化浙江南部近海是咸淡水交汇区域, 是多种广温广盐性鱼类栖息、生长和繁殖的重要海域(戴小杰等, 2019)。研究发现, 2016年和2020年春季调查均以暖水种鱼类居多, 其次为暖温种, 各个适温性的鱼类所占比例并无较大变化。从渔获鱼类的适温性可以看出, 受沿岸海流和台湾暖流影响(王玉衡等, 1990), 该海域的区系组成主要为暖水种和暖温种, 另外由于浙江南部近海具有亚热带海区的特点, 因此暖水种在该海域更占优势(卢占晖等, 2009), 与以往的调查结果(刘勇等, 2007; 张洪亮等, 2013)一致。然而, 从调查种类来看, 2020年春季调查鱼类30种, 远低于2016年春季调查的65种, 仅是2016年调查种类数量的46.15%, 出现这一现象的主要原因与人类活动密切相关, 高强度渔业捕捞会迫使鱼类自身在繁殖、索饵、迁移等方面做出适应性变化, 而一些适应性较差的鱼类会出现数量减少甚至灭绝的情况(戴小杰等, 2019; 于南京, 2021); 另一方面, 人类活动导致的海洋环境污染以及气候变暖等也是进一步造成鱼类物种多样性降低的原因之一(程炎宏等, 2001)。
根据聚类结果, 2016年和2020年春季鱼类群落可划分为南部群落和北部群落, 与杜晓雪等(2018)发现浙江南部近海鱼类群落结构呈现南、北向的空间格局结果一致。研究中南、北鱼类群落组成中典型种和分歧种有显著的年际变化, 相较于2016年, 2020年南部群落主要典型种由二长棘鲷、带鱼、鳀、龙头鱼转变为带鱼和七星底灯鱼, 北部群落主要典型种由带鱼、龙头鱼转变为带鱼、镰鲳和小黄鱼; 镰鲳和小黄鱼对组间相异性贡献率大幅提升, 与带鱼共同成为南、北群落间的主要分歧种。这种转变不仅表明南、北鱼类群落组成存在时空差异性, 还体现了群落结构的演替现象, 即伴随着龙头鱼、鳀和二长棘鲷等传统鱼类在群落中的功能地位下降, 镰鲳和小黄鱼在群落的地位上升; 七星底灯鱼渔获数量最高, 逐渐取代了鳀在南部群落中的位置, 成为群落中的关键饵料(王士聪等, 2022); 带鱼在群落结构变动中始终保持优势地位, 王士聪等(2022)通过食物网生态网络分析发现带鱼是浙江南部近海的关键种, 与较多鱼类间保持营养相互作用, 其利用资源的能力和竞争能力均较强。整体来看, 浙江南部近海鱼类群落结构是动态的, 主要仍以广食性和中小型鱼类为主, 种类演替正是其适应外部干扰和内部种间相互作用的最好体现。不过, 水域内饵料鱼类数量的激增, 表明食物网下行控制在变弱, 这对群落结构稳定性将会造成一定威胁, 值得关注。
3.2 优势种变化纵观浙江南部近海的优势种变化(表 4), 可以发现该水域的优势种年际变化显著, 小型鱼类优势种从矛尾虾虎鱼(Chaeturichthys stigmatias)转变为鳀、七星底灯鱼, 银姑鱼和龙头鱼优势度下降, 广食性的带鱼在群落结构中的优势地位凸显, 这种优势种的变化体现了群落在应对捕捞压力及环境变动下的动态适应性, 如金显仕等(2000)发现, 在高强度捕捞下, 莱州湾鱼类群落个体较大、营养层次较高的优势种群会逐步被个体小、营养层次低的小型中上层鱼类替代; 黄东海鱼类群落结构中一些特定种类会经常随主要环境因子的变化逐渐或同步变化(唐启升等, 2000)。与东海其他水域相比, 均能发现带鱼在群落中占据了绝对的优势地位, 杜萍等(2020)认为带鱼具有偏r-选择型的生活史特征、摄饵可塑性强的习性以及较强的栖息地环境适应性, 高春霞(2020)研究发现带鱼具有强烈的占有食物资源及利用资源的能力, 这些都保证了带鱼在高强度的捕捞压力下始终“衰而不竭”。
表 4 浙江南部近海及其邻近海域不同年份鱼类优势种 Tab. 4 Dominant fish species in offshore waters of southern Zhejiang Province and the adjacency in different years
| 海域 | 调查时间(年.月) | 优势种 | 文献 |
| 浙江南部近海 | 2020.05 | 七星底灯鱼、带鱼 | 本研究 |
| 2016.05 | 带鱼、七星底灯鱼、鳀 | ||
| 浙江南部近海 | 2011.06 | 矛尾虾虎鱼、银姑鱼、带鱼 | 张洪亮等(2013) |
| 2011.05 | 矛尾虾虎鱼、龙头鱼、银姑鱼 | ||
| 2011.04 | 龙头鱼、矛尾虾虎鱼、赤鼻棱鳀 | ||
| 浙江中部近海 | 2008.05 | 带鱼、短鳄齿鱼、麦氏犀鳕 | 卢占晖等(2009) |
| 东海中部 | 2000.12 | 带鱼、发光鲷、短鳄 齿鱼 |
刘勇等(2007) |
本研究还发现优势种的IRI值在年份间的波动较大, 2020年调查优势鱼种带鱼和鳀的IRI值大幅下降(8317→5373; 1245→168), 而七星底灯鱼的IRI值大幅上升(2671→9159), 超过带鱼成为绝对优势种, 这也间接表明小型饵料鱼类在浙江南部近海的占比显著提高, 究其原因, 小型鱼类对捕捞等干扰因素的敏感程度较低, 同时海域内的高营养级生物减少, 小个体生物的被捕食压力降低(卢占晖等, 2009), 从而使得七星底灯鱼作为小型饵料鱼类能够逐步从群落结构的演替中脱颖而出; 另外林龙山等(2005)研究发现带鱼和七星底灯鱼存在强摄食关系, 因此两者之间可能存在一种此消彼长的关系。
3.3 鱼类生物量分布及其受干扰状况生物量谱揭示生态系统中所有成员的生物量分布模式(左涛等, 2008)。研究结果表明, 2016年和2020年春季鱼类生物量随平均体质量的变化趋势和变化程度大致相同, 但谱线斜率均小于–1, 这说明近年来该海域鱼类群落的生物量随平均体质量的增加而减少。有研究表明, 捕捞是影响谱线斜率的一个重要因素(Zwanenburg, 2000), 捕捞死亡会使得鱼类生物量谱的斜率下降, 即大个体鱼类减少(Blanchard et al, 2009)。从生物量分布情况(图 4)可以看出, 2020年10 g以下的小型鱼类生物量占比较2016年大幅提高, 主要以七星底灯鱼的生物量提高为主。以带鱼为代表的中型鱼类依旧在该海域占据较大比例, 但带鱼的平均个体质量较2016年有所降低, 这使得带鱼在总生物量上的占比也有显著下降, 说明带鱼的大个体减少而小个体增多, 呈现明显的小型化趋势。大型鱼类的生物量较2016年占比虽有小幅提高, 但从整个分布模式来看, 大个体鱼类的生物量占比依然不高。
鱼类群落的ABC曲线可以反映群落中的中、大型鱼类和小型鱼类相对数量的变化以及个体大小组成的变化, 而群落优势种一定程度上决定了生物量优势度曲线和数量优势度曲线的位置(李忠义等, 2017)。从本研究的ABC曲线结果可以看出, 2016年春季鱼类生物量优势度曲线与数量优势度曲线相交, 仅有带鱼的生物量占据主要优势, 生物量占比高达64.38%, 而数量优势第一位的七星底灯鱼数量占比49.93%, 群落虽处于中度干扰, 但已经濒于严重干扰的状态; 2020年春季数量优势度曲线完全在生物量优势度曲线之上, 此时带鱼的生物量占比50.45%, 而七星底灯鱼数量占比高达81.15%, 小个体饵料鱼类的数量呈绝对优势, 群落处于严重干扰状态。群落中以七星底灯鱼为代表的小个体饵料鱼类的占比不断提高, 而原先生物量曲线占据优势的带鱼的资源量出现一定程度的衰退, 导致上述现象的原因, 一方面是以带鱼为代表的优势鱼类迫于过度捕捞的压力出现了一定程度的适应性改变, 如性成熟年龄提早、性成熟时间缩短等(周永东等, 2002), 使得其呈现小型化趋势, 食性出现变动, 这种变化在以往的研究(刘勇等, 2007; 卢占晖等, 2009; 张琥顺等, 2021)中也得到了证实; 另一方面中高级捕食者的减少也为小型鱼类的发展提供了一定的生存条件(张琥顺等, 2021)。由此进一步证实浙江南部近海鱼类群落结构中捕食者资源正在衰退, 营养下行效应降低, 群落稳定性下降。
3.4 鱼类群落结构与环境因子的关系鱼类群落与环境因子之间存在着密切的关系, 且生境条件的变化在一定程度上会导致群落结构的异质化(Porto et. al, 2018)。本研究中, 2016年和2020年春季影响鱼类群落分布的主要理化因子均为表层水温、水深和盐度, 尤其是表层水温的影响最为显著, 符合水温是控制鱼类生长和资源密度最重要的水文要素这一说法(Gibson, 1994)。调查中两个航次春季的表层水温范围为19.9~24.2 ℃, 是暖水性鱼类生存的适宜温度(王小荟, 2013), 结果也显示暖水性鱼种数量在群落组成中占据优势, 如七星底灯鱼和龙头鱼等暖水种数量达到60%以上, 进一步证明了水温对鱼类分布的影响。本研究与胡翠林等(2018)对带鱼保护区春季鱼类群落主要环境因子的分析结果基本一致, 而与马稳等(2022)通过GAM模型分析出2020年春季影响鱼类群落贡献率最大的环境因子为盐度的结论存在差异, 究其原因, 这主要由于其采用分季节分别建模的方法, 从而降低了温度因素的影响性, 另外模型中的水文因子数据分辨率也可能会造成研究差异性。
水深和盐度也是影响鱼类分布的关键影响因子, 这在多项研究中已被证实, 水深可以通过影响饵料生物的分布、溶解氧含量以及光照等来间接影响鱼类分布(王雪辉等, 2010; Pinault et al, 2014; 胡翠林等, 2018), 如柳晓雪等(2020)研究发现浙江南部近海黄鲫受浮游饵料生物栖息深度影响, 最适水深变浅, 且胡翠林等(2018)发现多数鱼类分布与水深呈正相关性, 这在本研究的排序图中同样能发现这一特征。本调查水域为咸淡水交汇区域, 盐度范围广, 为不同生态类型的鱼类提供了良好的栖息环境, 马稳等(2022)研究发现浙江南部近海鱼类资源密度会随着盐度的增加而呈现增加的趋势, 通过RDA排序图也可发现多数鱼类聚集在更靠近外海的调查站点。
4 结论2020年相比2016年, 浙江南部近海春季鱼类群落结构发生了一定变化, 主要体现在以下几个方面: (1) 鱼类群落组成呈现小型化趋势, 且多样性明显下降; (2) 优势种逐渐由以七星底灯鱼为代表的小型饵料鱼类占据主导地位, 食物网的营养下行效应降低; (3) 鱼类群落的受干扰程度正在不断加剧, 群落结构稳定性减弱。通过年份间变化分析, 浙江南部近海的鱼类群落结构在站点簇和种类组成并不具有一定的空间连续性, 因此该海域的鱼类群落在空间格局上稳定性较低, 这对该海域的渔业资源可持续发展带来了严峻的挑战, 尤其是在全球气候变化、环境污染和人类捕捞等多重压力下, 如何维持渔业群落结构稳定性、加快渔业资源的恢复和增长, 值得渔业研究者和管理者们的重视和思考。
参考文献
丁朋朋, 高春霞, 田思泉, 等, 2019. 浙江南部近海蟹类群落结构及其与环境因子的关系. 海洋渔业, 41(6): 652-662 DOI:10.3969/j.issn.1004-2490.2019.06.002 |
于南京, 2021. 舟山群岛近岸海域鱼类群落结构及生物多样性的年际变化[D]. 舟山: 浙江海洋大学: 47-48.
|
马稳, 秦松, 赵静, 2022. 浙江南部近海鱼类资源季节分布特征及其影响因素. 渔业科学进展, 43(3): 1-11 |
王士聪, 杨蕊, 高春霞, 等, 2022. 基于生态网络结构的浙江南部近海鱼类群落关键种识别. 中国水产科学, 29(1): 118-129 |
王小荟, 2013. 海州湾主要鱼种的空间分布及其与环境因子的关系[D]. 青岛: 中国海洋大学.
|
王玉衡, 蒋国昌, 董恒霖, 1990. 春季浙江南部海区溶解氧、pH值和营养盐分布特征及相互关系研究. 海洋学报, 12(5): 654-660 |
王雪辉, 邱永松, 杜飞雁, 等, 2010. 北部湾鱼类群落格局及其与环境因子的关系. 水产学报, 34(10): 1579-1586 |
左涛, 王俊, 金显仕, 等, 2008. 春季长江口邻近外海网采浮游生物的生物量谱. 生态学报, 28(3): 1174-1182 DOI:10.3321/j.issn:1000-0933.2008.03.033 |
卢占晖, 苗振清, 林楠, 2009. 浙江中部近海及其邻近海域春季鱼类群落结构及其多样性. 浙江海洋学院学报(自然科学版), 28(1): 51-56 DOI:10.3969/j.issn.1008-830X.2009.01.010 |
田明诚, 孙宝龄, 杨纪明, 1993. 渤海鱼类区系分析. 海洋科学集刊: 157-167 |
刘前, 王学锋, 吕少梁, 等, 2021. 湛江港海域游泳动物群落结构及多样性分析. 广东海洋大学学报, 41(2): 103-110 DOI:10.3969/j.issn.1673-9159.2021.02.014 |
刘勇, 李圣法, 陈学刚, 等, 2007. 东、黄海2000年冬季底层鱼类群落结构及其多样性. 海洋科学, 31(10): 19-24 DOI:10.3969/j.issn.1000-3096.2007.10.009 |
刘勇, 程家骅, 2015. 东海、黄海秋季渔业生物群落结构及其平均营养级变化特征初步分析. 水产学报, 39(5): 691-702 |
杜晓雪, 田思泉, 王家启, 等, 2018. 浙江南部近海鱼类群落结构的时空特征. 大连海洋大学学报, 33(4): 522-531 |
杜萍, 陈全震, 李尚鲁, 等, 2020. 东海带鱼资源变动及其栖息地驱动因子研究进展. 广东海洋大学学报, 40(1): 126-132 |
李圣法, 2008. 以数量生物量比较曲线评价东海鱼类群落的状况. 中国水产科学, 15(1): 136-144 |
李忠义, 吴强, 单秀娟, 等, 2017. 渤海鱼类群落结构的年际变化. 中国水产科学, 24(2): 403-413 |
杨柯迩, 周曦杰, 秦松, 等, 2022. 浙江南部近海鱼类粒径谱特征. 南方水产科学, 18(1): 10-21 |
宋海棠, 丁天明, 1995. 浙江渔场鲐鱼Scomber japonicus蓝园鲹Decapterus maruadsi不同群体的组成及分布. 浙江水产学院学报, 14(1): 29-35 |
张洪亮, 宋之琦, 潘国良, 等, 2013. 浙江南部近海春季鱼类多样性分析. 海洋与湖沼, 44(1): 126-134 |
张琥顺, 宋普庆, 李渊, 等, 2021. 东海中南部秋季游泳动物多样性及群落结构. 应用海洋学学报, 40(4): 575-586 |
林龙山, 严利平, 凌建忠, 等, 2005. 东海带鱼摄食习性的研究. 海洋渔业, 27(3): 187-192 |
金显仕, 邓景耀, 2000. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 8(1): 65-72 |
周永东, 徐汉祥, 刘子藩, 等, 2002. 东海带鱼群体结构变动的研究. 浙江海洋学院学报(自然科学版), 21(4): 314-320 |
胡翠林, 张洪亮, 张亚洲, 等, 2018. 带鱼保护区春秋季鱼类群落特征及与环境因子的关系. 水产学报, 42(5): 694-703 |
柳晓雪, 高春霞, 田思泉, 等, 2020. 基于栖息地适宜指数的浙江南部近海黄鲫最适栖息地分布. 中国水产科学, 27(12): 1485-1495 |
晏磊, 谭永光, 杨吝, 等, 2015. 南海珠江口沿岸张网渔业资源群落结构分析. 生物学杂志, 32(5): 52-57 |
徐姗楠, 郭建忠, 范江涛, 等, 2020. 大亚湾夏季鱼类生物量粒径谱年际变化特征. 南方水产科学, 16(4): 28-38 |
高春霞, 2020. 基于稳定同位素技术的浙江中南部近海渔业生物群落营养结构研究[D]. 上海: 上海海洋大学: 3-5.
|
唐启升, 苏纪兰, 2000. 中国海洋生态系统动力学研究-I-关键科学问题与研究发展战略. 北京: 科学出版社, 66-71
|
程炎宏, 樊伟, 2001. 东海区海洋捕捞产量的时间序列分析研究. 中国水产科学, 8(3): 31-34 |
程济生, 2005. 黄海无脊椎动物资源结构及多样性. 中国水产科学, 12(1): 68-75 |
程家骅, 丁峰元, 李圣法, 等, 2006. 夏季东海北部近海鱼类群落结构变化. 自然资源学报, 21(5): 775-781 |
戴小杰, 杨志金, 田思泉, 等, 2019. 浙江南部近海鱼类分类多样性研究. 海洋学报, 41(8): 43-51 |
BLANCHARD J L, JENNINGS S, LAW R, et al, 2009. How does abundance scale with body size in coupled size-structured food webs?. Journal of Animal Ecology, 78(1): 270-280 |
CLARKE K R, 1993. Non-parametric multivariate analyses of changes in community structure. Austral Ecology, 18(1): 117-143 |
GIBSON R N, 1994. Impact of habitat quality and quantity on the recruitment of juvenile flatfishes. Netherlands Journal of Sea Research, 32(2): 191-206 |
MAGNUSSEN E, 2002. Demersal fish assemblages of Faroe Bank: species composition, distribution, biomass spectrum and diversity. Marine Ecology Progress Series, 238: 211-225 |
PINAULT M, BISSERY C, GASSIOLE G, et al, 2014. Fish community structure in relation to environmental variation in coastal volcanic habitats. Journal of Experimental Marine Biology and Ecology, 460: 62-71 |
PINKAS L, OLIPHANT M S, IVERSON I L K, 1971. Food Habits of Albacore, Bluefin Tuna, and Bonito in California Waters [M]. 1-105.
|
PLATT T, DENMAN K, 1977. Organisation in the pelagic ecosystem. Helgoländer wiss. Meeresunters, 30(1/2/3/4): 575-581 |
PORTO T J, PINTO-DA-ROCHA R, DE ROCHA P L B, 2018. Regional distribution patterns can predict the local habitat specialization of arachnids in heterogeneous landscapes of the Atlantic forest. Diversity and Distributions, 24(3): 375-386 |
RICE J, 2003. Environmental health indicators. Ocean & Coastal Management, 46(3/4): 235-259 |
ROCHET M J, TRENKEL V M, 2003. Which community indicators can measure the impact of fishing? A review and proposals. Canadian Journal of Fisheries and Aquatic Sciences, 60(1): 86-99 |
SHELDON R W, PRAKASH A, SUTCLIFFE W H JR, 1972. The size distribution of particles in the ocean. Limnology and Oceanography, 17(3): 327-340 |
SHIN Y J, ROCHET M J, JENNINGS S, et al, 2005. Using size-based indicators to evaluate the ecosystem effects of fishing. ICES Journal of Marine Science, 62(3): 384-396 |
TER BRAAK C J F, 1989. CANOCO—an extension of DECORANA to analyze species-environment relationships. Hydrobiologia, 184(3): 169-170 |
WIJEYARATNE W M D N, BELLANTHUDAWA B K A, 2018. Abundance-Biomass Comparison approach to assess the environmental stressors in Diyawannawa wetland in monsoonal and non-monsoonal seasons. Sri Lanka Journal of Aquatic Sciences, 23(2): 135-149 |
YEMANE D, FIELD J G, LESLIE R W, 2005. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves. ICES Journal of Marine Science, 62(3): 374-379 |
ZWANENBURG K C T, 2000. The effects of fishing on demersal fish communities of the Scotian shelf. ICES Journal of Marine Science, 57(3): 503-509 |