 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 郝贵杰, 林锋, 黄爱霞, 叶霆, 姚嘉赟, 潘晓艺, 袁雪梅, 徐磊, 胡金春, 沈锦玉. 2022.
- HAO Gui-Jie, LIN Feng, HUANG Ai-Xia, YE Ting, YAO Jia-Yun, PAN Xiao-Yi, YUAN Xue-Mei, XU Lei, HU Jin-Chun, SHEN Jin-Yu. 2022.
- 草鱼(Ctenopharyngodon idellus)出血病二价细胞灭活疫苗体液免疫效果及免疫保护率评价研究
- EVALUATION OF HUMORAL IMMUNE EFFICACY AND PROTECTION RATE OF BIVALENT CELL INACTIVATED VACCINE AGAINST GRASS CARP HEMORRHAGIC DISEASE
- 海洋与湖沼, 53(6): 1467-1475
- Oceanologia et Limnologia Sinica, 53(6): 1467-1475.
- http://dx.doi.org/10.11693/hyhz20220300082
文章历史
-
收稿日期:2022-03-31
收修改稿日期:2022-06-10




2. 衢州市水产技术推广中心 浙江衢州 324000
草鱼(grass carp, Ctenopharyngodon idellus)是我国传统的“四大家鱼”之一, 因营养高、味道鲜美且饲养成本低而广受欢迎, 根据2021年中国渔业统计年鉴, 2020年草鱼产量占淡水养殖总产量的18%, 依然是中国淡水养殖鱼类产量的卫冕冠军, 比2019年增加0.69%, 具有较高的经济价值。但随着草鱼大规模集约化养殖的发展, 草鱼病害时有发生, 如细菌性疾病、病毒性出血病、寄生虫病等, 损失最大的是草鱼出血病, 据不完全统计, 每年由该病导致的经济损失达20余亿元(陈爱平等, 2010)。其病原为草鱼呼肠孤病毒(grass carp reovirus, GCRV), 该病毒属于水生呼肠孤病毒属, 是该属成员中毒力最强的一种, 也是中国分离鉴定, 并在国际上首次完成全基因序列分析的第一株鱼类病毒, 可导致草鱼在鱼种阶段发生出血病, 死亡率一般为30%~50%, 最高可达60%~80% (Winton et al, 1987; 柯丽华等, 1990; Rangel et al, 1999; 王方华等, 2006)。GCRV为双链分节段RNA病毒, 基因组分11个节段, 病毒颗粒为二十面体对称的球形颗粒, 病毒粒子平均直径为60~70nm, 具有双层衣壳, 没有囊膜, 对氯仿和乙醚处理不敏感, 在pH 3~11范围内十分稳定(Francki et al, 1991; Fang et al, 2000, 2005; Qiu et al, 2001)。GCRV病原学比较复杂, 目前国内报道有几十个分离株, 但不同分离株产生的细胞病变效应(CPE), 对草鱼的致病力差异较大, 而且其基因组带型及序列也存在差异。目前还没在血清学上对其进行分类, 但根据现有病毒分离株的核苷酸及氨基酸的序列信息, 构建系统进化树进行比对分析, 共有三种基因型GCRV, 即GCRV Ⅰ型(GCRV-Ⅰ, 代表株为GCRV-873与GCRV- ZV8909)、GCRV Ⅱ型(GCRV-Ⅱ, 代表株为GCRV- HZ08与GCRV-GD108)和GCRV Ⅲ型(GCRV-Ⅲ, 代表株为GCRV-104)。近年来, 国内多家单位包括本实验室针对草鱼出血病开展的流行病学研究结果表明, 临床流行的毒株主要为GCRV Ⅰ型和Ⅱ型, Ⅲ型很少(张超等, 2010; Wang et al, 2012; 李永刚等, 2013; 杨映等, 2015)。
草鱼出血病一直是我国草鱼养殖中最难控制的疾病, 但目前未见病毒病的特效治疗药物, 因此采用疫苗免疫, 再联合早期诊断, 可有效降低该病的危害, 是控制病毒病最可取也是最有效的方法。传统的“土法疫苗”组织浆灭活疫苗在20世纪70、80年代时为各地控制草鱼出血病疫情起到了积极成效, 但其需大量鱼体扩增病毒, 费时费力, 并存在扩散病毒的危险(刘栭等, 1977)。目前细胞培养弱毒疫苗已取得了疫苗生产批准文号, 在多个地区进行了推广应用, 对草鱼出血病的控制起了很大的作用, 但也存在病毒毒力返强的风险(许淑英等, 1994); 因此研制细胞灭活疫苗尤其是二价细胞灭活疫苗具有较大的经济和社会效益。课题组前期研究表明疫苗免疫刺激机体启动了非特异性和特异性免疫应答(奕志娟等, 2015)。本研究从体液免疫及免疫保护率的角度对草鱼出血病二价细胞灭活疫苗进行免疫效果评价, 以表达纯化的GCRV结构蛋白VP7作为测试抗原, 以实验室自制的草鱼IgM单抗为一抗, 以商业化的羊抗鼠HRP酶标抗体为二抗, 建立了测试未知抗体的三抗体夹心酶联免疫方法(TAS-ELISA)检测方法评价疫苗特异性体液免疫应答的免疫效果, 并在实验室开展了二价疫苗免疫保护率实验, 为进一步开展二价疫苗的免疫示范和新兽药证书申报提供基础数据。
1 材料与方法 1.1 主要仪器和试剂超微量分光光度计(NanoDrop 2000); Mx3005P荧光定量PCR仪(Stratagene公司); MIR-154低温恒温培养箱(日本三洋); 550型酶标仪(Bio-rad); CKX31倒置显微镜(奥林巴斯)。RNA提取试剂盒购自QIAGEN公司, 反转录试剂盒[FastQuant RT Kit (with gDNase)]、质粒小提试剂盒(TIANprep Mini Plasmid Kit)等购自TIANGEN公司, iTaqTM Universal SYBR® Green Supermix购自Bio-RAD公司, SDS-PAGE凝胶配制试剂盒, ELISA相关试剂等购自索莱宝生物科技有限公司, M199细胞培养基、0.25%胰酶消化液、双抗(青霉素、链霉素)均购自上海普飞生物技术有限公司, 引物合成及序列测定委托南京金斯瑞生物科技有限公司, 其他生化试剂均为国产分析纯试剂。
1.2 实验用质粒、细胞株及实验动物等pET28a原核表达载体由宁波大学惠赠, 大肠杆菌DH5α、BL21 (DE3)、草鱼IgM单抗、CIK细胞由本实验室保存; 所用GCRV毒株为本实验室分离保存, 其中标准参考株GCRV873细胞悬液由中国水产科学研究院长江水产研究所鱼病研究室惠赠, (20±5) g的健康草鱼购自长兴某养殖场。
1.3 两种基因型GCRV的实时荧光绝对定量PCR (AQ-PCR)方法的建立根据GenBank公布的基因Ⅰ型和Ⅱ型GCRV第六基因序列设计绝对荧光定量PCR特异性扩增引物, 目的片段大小分别为102 bp和169 bp。上游和下游引物序列分别为F1: 5′-CCCA CATCCTCCTCCAAT-3′, R1: 5′-AACGACTTCGCCCTTGTA-3′; F2: 5′-GGACG GCGGCTGC TATGAT-3′, R2: 5′-GCCCGTAACGCAG GGTAT-3′。
按常规方法构建含上述目的基因的阳性重组质粒。用NanoDrop2000测定所制备的基因Ⅰ型、Ⅱ型GCRV的标准质粒的浓度和纯度, 根据公式: 质粒拷贝数(单位: copies/μL)=6.02×1023(单位: copies/mol)×质粒浓度(单位: g/μL)/质粒分子量(单位: g/mol), 得出质粒拷贝数(李丽等, 2011)。基因Ⅰ型、Ⅱ型GCRV的标准质粒浓度分别为157.1、118.8 ng/μL, 按公式得拷贝数分别为1.4×1012、6.4×1011 copies/μL。将基因Ⅰ型、Ⅱ型GCRV质粒分别用ddH2O进行10倍梯度稀释, 稀释成1×108~1×102 copies/μL的7个梯度, 将其分别作为模板进行荧光定量PCR反应, 每个模板重复做3次, 采用20 μL PCR反应体系: iQ SYBR® Green SuperMix 10 μL, 荧光定量PCR引物(10 μmol/L) 各0.5 μL, 模板2 μL, ddH2O 7 μL。PCR反应程序: 95 ℃预变性10 min; 95 ℃变性30 s, 58 ℃退火20 s, 72 ℃延伸30 s, 40个循环。
1.4 细胞病毒液培养及其病毒含量测定两种基因型GCRV (GCRV-ZV8909及GCRV-HZ13)分别利用细胞瓶培养的方法进行扩增培养, 待基因Ⅰ型GCRV产生明显的细胞病变及基因Ⅱ型GCRV病毒复制达到一定的拷贝数后收获细胞病毒悬液, 经反复冻融3次后将其分别合并备用。
分别提取上述两种基因型GCRV细胞病毒液RNA并进行反转录, 按照1.3中方法进行定量PCR扩增, 根据系统给出的结果计算两种细胞病毒液的病毒拷贝数。
1.5 草鱼出血病二价细胞灭活疫苗的制备按相同病毒拷贝数将GCRV-ZV8909和GCRV-HZ13细胞病毒悬液混合作为病毒原液, 按照体积加入终浓度为3.75 mL/L福尔马林溶液(含40%甲醛), 用恒温水浴锅65 ℃灭活2 h, 灭活后的病毒液重接CIK细胞, 培养7 d后观察细胞病变。经灭活检验后, 将疫苗原液用生理盐水进行稀释, 分别进行50倍、100倍稀释备用。
1.6 草鱼免疫及血清样品制备草鱼在恒温循环系统中充氧饲育, 水温24~26 ℃。试验共分4个组: 疫苗原液组、50倍稀释组、100倍稀释组、生理盐水对照组, 每组50尾, 分别为腹腔注射, 剂量均为0.2 mL/尾。各组在免疫后的3、5、7、14、21、28、42、56、70、84 d各取3尾鱼, 每尾鱼采集血液, 4 ℃冰箱过夜放置后2 500 r/min, 离心10 min收集血清置于–80 ℃的超低温冰箱中保存备用。
1.7 两种基因型GCRV S10基因原核表达载体的构建及鉴定分别根据GCRV-Ⅰ及GCRV-Ⅱ第10基因片段序列(Ⅰ型S10 909 bp, Ⅱ型1 124 bp)和pET-28a表达序列的特征, 设计原核表达引物F3:
5′-CGCATATGCCACTGCACATGATTCCGCAAG-3′;
R3: 5′-CGCTCGAGTTAATCGGACGGTTCCACATGC-3′;
F4: 5′-CGCATATG ATGGCGGGTGTGTCTCTCAACA-3′
R4: 5′-CGCTCGAG CTACAGCATCTGCGCAAATATA-3′。
该两对引物上下游引物5′端分别含Nde Ⅰ及Xho Ⅰ酶切位点, 扩增片段分别为840 bp和1 050 bp。PCR反应体系按试剂盒配制, cDNA模板为各病毒悬液提取RNA并反转录获得, 纯化回收PCR扩增产物后, 将其分别克隆至pMD19-T载体, 进行阳性克隆筛选并测序。重组表达质粒pET-28a-S10的构建采用常规酶切连接方法, 连接产物转化至BL21 (DE3)感受态细胞, 进行阳性克隆筛选并测序, 同时进行重组菌保种待用。
1.8 重组蛋白的表达纯化与鉴定 1.8.1 重组蛋白的诱导表达取上述保存的菌液30 μL, 接种至3 mL含有100 μg/mL氨苄青霉素的LB液体培养基中, 置速度为250 r/min的37 ℃摇床过夜培养。第2天分管培养新鲜菌液, 同样条件恒温摇至OD600结果为0.6~0.8时, 加入IPTG诱导, 其终浓度为1 mmol/L, 同时设置未诱导组, 分别放至两种温度的摇床进行诱导培养, 37 ℃摇床培养4 h, 15 ℃摇床培养16 h, 然后收集菌液, 8 000 r/min离心10 min后收集菌体, 加入2倍体积的PBS重悬菌体, 再加入溶菌酶进行超声破碎, 其终浓度为1 mg/mL, 最后, 分别取全细胞裂解液以及经离心处理的沉淀和上清进行SDS-PAGE分析。
1.8.2 表达蛋白纯化与Western blot鉴定重组菌液经37 ℃摇床扩大培养至OD600结果为0.6~0.8时, 如上加入IPTG进行诱导培养16 h后收集菌体。按照Ni-Agarose His标签蛋白纯化试剂盒说明进行重组蛋白纯化, 依次进行装柱、平衡、菌体裂解、离心、上柱、冲洗、洗脱等。分别收集蛋白洗脱液, SDS-PAGE电泳分析, 并进行蛋白免疫印迹(Western blot)鉴定。
1.9 受免草鱼体液免疫效果评价免疫草鱼不同时间血清抗体水平测试: 用实验室前期建立的TAS-ELISA方法进行测定, 首先用pH 9.6的碳酸盐缓冲液即包被液CBS稀释上述纯化的VP7融合蛋白至终浓度为50 μg/mL, 按照100 µL/孔的量加入96孔酶标板, 置4 ℃冰箱过夜包被, 用PBST洗液充分洗涤包被板, 洗涤3~4次, 每次3~5 min; 用含8%脱脂奶粉的PBS封闭, 200 µL/孔, 37 ℃封闭2 h, 同上洗涤。将草鱼血清分别进行1︰80稀释, 每孔加入100 µL, 37 ℃孵育1h, 同上洗涤。然后每孔加入100 µL 1︰1 000稀释的草鱼IgM单抗腹水(本室制备), 37 ℃孵育1h, 同上洗涤。每孔加入50 µL 1︰2 000稀释的羊抗鼠IgG酶标抗体, 37 ℃孵育1h, 同上洗涤。每孔加入200 μL TMB 37 ℃避光孵育显色10 min, 用50 μL 2mol/L浓硫酸终止反应后, 于酶标仪上读取OD450值。P/N≥2.1者判为阳性, P/N < 2.1者判为阴性(P代表TAS-ELISA所测受免草鱼血清中抗体的OD值, N代表所测健康草鱼的相应血清中抗体的OD值)。
受免草鱼血清抗体效价测定: 根据上述测定结果, 分别取与两种基因型病毒抗原结合抗体水平达到高峰时的草鱼血清, 进行1︰50, 1︰100, 1︰200, 1︰400, 1︰600, 1︰800, 1︰1 200梯度稀释, 按照上述方法分别测定抗两种基因型GCRV抗原的血清抗体效价, 未免疫草鱼血清分别做同比例稀释进行对照。
1.10 相对免疫保护率测定草鱼免疫按照1.6中方法进行, 0.2 mL/尾, 每组免疫30尾, 注射同样剂量的PBS作为对照组。攻毒所用GCRV为实验室自行分离保存的毒株即江西株JX2008, 其半数致死量为107.5 LD50/mL, 半数组织细胞感染量为109.0 TCID50/mL, 攻毒注射在草鱼免疫后35 d进行, 腹腔注射剂量为0.2 mL。水温28 ℃左右饲养, 观察15 d并记录死亡情况, 计算对照组和试验组死亡率, 并按下列公式进行计算:
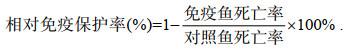 (1)
(1)正常生长的CIK细胞接种GCRV-ZV8909和GCRV-HZ13后, 用倒置荧光显微镜观察其病变情况如图 1, GCRV-HZ13株可见部分细胞脱落死亡, 但不表现出明显的破鱼网状, 而GCRV-ZV8909株则出现边缘不整齐的空洞, 少许细胞脱落, 随着病变发展, 最后呈现典型破鱼网状, 与GCRV-Ⅰ型代表株GCRV-873的细胞病变一致, 而对照细胞未出现任何病变。
 |
| 图 1 倒置荧光显微镜观察GCRV感染细胞后的病变(400×) Fig. 1 Fluorescent microscopic observation of the CPE of GCRV cultured with the cells (400×) 注: a. 对照(正常CIK细胞); b. GCRV-HZ13感染CIK细胞; c. GCRV-ZV8909感染CIK细胞; d. GCRV-873感染CIK细胞 |
将基因Ⅰ型、Ⅱ型GCRV质粒梯度稀释后选取1×108~1×102 copies/μL的7个梯度作为标准品进行实时荧光定量PCR扩增后, 由系统自动生成的基因Ⅰ型、Ⅱ型GCRV的熔解曲线及标准曲线可知, 基因Ⅰ型、Ⅱ型GCRV质粒在梯度稀释浓度范围内具有较好的线性关系, 显示本研究建立的基因Ⅰ型、Ⅱ型GCRV质粒标准曲线能够准确地反映目的基因的扩增。由熔解曲线可知产物的Tm(DNA熔解温度)值均一, 熔解曲线表现为单一的峰, 说明引物的特异性较好。
2.3 两种基因型GCRV在细胞病毒悬液中拷贝数测定结果根据1.3中AQ-PCR方法测定1.4中病毒悬液RNA得出的Ct值(循环阈值), 再由标准曲线得出的公式及试验过程中的稀释情况, 计算GCRV-ZV8909病毒液和GCRV-HZ13病毒液的拷贝数分别为2.0×109 copies/mL和1.5×106 copies/mL。
2.4 草鱼出血病二价细胞灭活疫苗的制备将GCRV-ZV8909和GCRV-HZ13按病毒拷贝数测定结果等比例混合, 均含病毒拷贝数约1.5×106 copies/mL, 即每mL混合病毒液中含有0.75 μL GCRV-ZV8909和999.25 μL GCRV-HZ13, 并以此作为原液。混合病毒原液按照1.5中方法进行灭活, 灭活检验结果表明合格, 将该疫苗原液用生理盐水进行稀释, 分别进行50倍、100倍稀释备用。
2.5 两种基因型GCRV S10基因原核表达载体的构建及鉴定 2.5.1 两种基因型GCRV S10基因编码区PCR扩增两种基因型GCRV目的基因经PCR扩增后电泳, 结果显示目的条带大小与预期一致, 分别为840 bp和1 050 bp, 测序为正确序列, 如图 2所示。
 |
| 图 2 两种基因型GCRV S10目的基因PCR扩增电泳图谱 Fig. 2 PCR amplification of S10 gene of genotype Ⅰ and Ⅱ GCRV 注: M: marker; 1: 目的片段; 左: GCRV-Ⅰ; 右: GCRV-Ⅱ |
将上述目的条带克隆于原核表达载体pET28a载体中, 构建了表达目的蛋白的原核表达载体pET28a-S10, 小量提取重组质粒, 进行双酶切鉴定, 结果表明: S10基因片段已成功克隆入pET28a中(图 3)。
 |
| 图 3 重组质粒PET-S10双酶切鉴定结果 Fig. 3 Restriction of recombinant plasmid PET-S10 注: M: marker; 1: 目的片段; 左: GCRV-Ⅰ; 右: GCRV-Ⅱ |
将上述重组质粒转化至大肠杆菌BL21 (DE3)中进行IPTG诱导表达, 分别于20 ℃诱导过夜; 37 ℃诱导4 h, 收集菌体, 超声破碎, 分别离心收集上清和沉淀, 沉淀用500 µL包涵体溶解液溶解, 聚丙烯酰胺凝胶电泳(SDS-PAGE)分析融合蛋白的表达情况, 结果表明, 两种融合蛋白均能在大肠杆菌中以包涵体形式表达, 分子量分别为32 kDa和41 kDa, 如图 4所示。
 |
| 图 4 GCRV-S10融合蛋白SDS-PAGE分析结果图 Fig. 4 Analysis results of the expression of PET-S10 by SDS-PAGE 注: M: marker; 1: 诱导前总蛋白; 2: 20 ℃上清; 3: 20 ℃沉淀; 4: 37 ℃上清; 5: 37 ℃沉淀。左: GCRV-Ⅰ; 右: GCRV-Ⅱ |
大量诱导表达融合蛋白, 超声破碎菌体, 镍琼脂糖亲和层析纯化, 进行SDS-PAGE及Western blot检测, 见图 5、图 6, 表明两个蛋白均获得了较好的纯化, 测定结果表明两个纯化蛋白的浓度分别为0.70和0.72 mg/mL; 蛋白纯度检测采用SDS-PAGE, 通过软件扫描测定, 纯度均大于80%。两个蛋白的表达纯化为进一步ELISA评价疫苗免疫效果提供了有用的试剂。
 |
| 图 5 GCRV-Ⅰ纯化蛋白SDS-PAGE及Western blotting分析 Fig. 5 Analysis results of the purified protein of GCRV-Ⅰ by SDS-PAGE and the Western blotting 注: M: prestained marker; 1: 目的蛋白 |
 |
| 图 6 GCRV-Ⅱ纯化蛋白SDS-PAGE及Western blotting分析 Fig. 6 Analysis results of the purified protein of GCRV-Ⅰ by SDS-PAGE and the Western blotting 注: M: prestained marker; 1: 目的蛋白 |
草鱼腹腔注射不同剂量的细胞灭活疫苗后, 血清中出现了抗GCRV抗体的变化(如图 7, 图 8), 抗基因Ⅰ型GCRV抗体的变化与对照组相比, 三种不同免疫剂量在接种疫苗5 d后, 草鱼血清抗GCRV的抗体稍有增加, 接种7 d后, 抗GCRV抗体均有明显增加, 28 d后增加更为显著, 达到最高峰, 但42 d后开始下降, 且一直持续到第84 d。抗基因Ⅱ型GCRV抗体的变化与对照组相比, 三种不同免疫剂量在接种疫苗5 d后, 草鱼血清抗GCRV的抗体稍有增加, 14 d后增加更为显著, 达到最高峰, 但21 d后开始下降, 且一直持续到第84 d。由图可以看出, 原液组与50倍稀释组产生的抗体水平基本相同, 且都高于100倍稀释组。
 |
| 图 7 草鱼腹腔注射疫苗不同时间后血清中抗Ⅰ型GCRV抗体水平 Fig. 7 The serum antibody levels of anti-GCRV-Ⅰ on different times post intraperitoneal injection of vaccine |
 |
| 图 8 草鱼腹腔注射疫苗不同时间后血清中抗Ⅱ型GCRV抗体水平 Fig. 8 The serum antibody levels of anti-GCRV-Ⅱ on different times post intraperitoneal injection of vaccine |
TAS-ELISA法测定血清效价, 结果表明, 抗基因Ⅰ型GCRV的血清效价是1︰800, 抗基因Ⅱ型GCRV的血清效价是1︰600。
2.9 相对免疫保护率测定按照1.8中方法测定核酸疫苗免疫草鱼的相对免疫保护率, 结果表明, 核酸疫苗pEGFP-N1-VP7以0.5 μg/尾注射免疫草鱼, 其相对免疫保护率为67%, 而对照组为30%, 显示出较好的免疫效果。
3 讨论2019年我国农业农村部印发了关于加快推进水产养殖业绿色发展的若干意见, 提出了三个方面的措施, 其中最重要的就是加强疫病防控, 全面实施苗种检疫及疫苗的推广, 因此, 在“减抗替抗”及绿色养殖的大环境下, 水产养殖用疫苗的推广势在必行。
草鱼是我国重要的淡水经济鱼类, “四大家鱼”之一, 产量约占我国淡水养殖鱼类产量的1/5, 连续多年单个鱼类产量排名第一。但在养殖过程中病害也较多, 危害较大的疾病俗称草鱼“四病”, 即草鱼出血病、烂鳃病、肠炎病、赤皮病, 其中每年引起草鱼损失最严重的为草鱼出血病, 病原为草鱼呼肠孤病毒, 目前公认的该病毒有三个基因型, 最早于1980年分离和研究的GCRV属于基因Ⅰ型(如GCRV-873株等); 张超等(2010)首次报道了基因Ⅱ型GCRV, 如GCRV-HZ08株; Fan等(2013)在国内首次分离并报道了基因Ⅲ型GCRV, 即HGDRV (GCRV-104)。不同基因型毒株各阶段序列同源性相似性为15.1%~ 46.1%不等, 各基因型之间交叉保护较低, 这也意味着即使免疫过单种疫苗, 仍然会有被其他基因型病毒感染的风险, 目前临床上流行的GCRV主要为基因Ⅰ型和Ⅱ型。
草鱼出血病疫苗的研究起步于20世纪70年代, 历经了组织奖灭活疫苗即“土法疫苗”、细胞灭活疫苗、减毒活疫苗、基因工程疫苗等。多年来, 许多科学家致力于该病毒的研究, 全国各地相继应用组织浆灭活疫苗和细胞灭活疫苗进行免疫防治, 使得出血病在一定程度上得到了控制。而目前代表最高研究水平的是草鱼出血病活疫苗(GCRV-892株), 2012年, 中国水产科学研究院珠江水产研究所将该活疫苗转让给大华农公司且重新获得了该产品的兽药生产批准文号: 兽药生字(2014)190026013, 至此, 我国水产疫苗开始进入产业化生产阶段, 该疫苗自上市以来, 逐渐在各草鱼主养区推广, 免疫草鱼的平均成活率为65%~90%, 比未免疫草鱼的成活率要高15%~20%, 显示出了较好的效果(吉华松等, 2019)。但该疫苗仅仅对基因Ⅰ型草鱼出血病有较好的保护。
本研究通过研制针对基因Ⅰ型和Ⅱ型GCRV的草鱼出血病二价细胞灭活疫苗, 通过绝对定量的方法, 测定基因Ⅰ型和Ⅱ型的病毒含量分别为2.0×109及1.5×106 copies/mL。用生理盐水将其分别进行50倍稀释和100倍稀释, 包括原液及生理盐水对照共4组, 通过腹腔注射免疫(20±5) g健康草鱼, 并开展了免疫效果评价。众所周知, 鱼类作为特异性与非特异性免疫并存的脊椎动物, 在病原微生物侵袭或疫苗免疫后都会产生非特异性免疫或特异性的体液免疫应答, 课题组之前从免疫基因及小分子的转录水平做过免疫效果评价, 与对照组相比, 注射免疫不同剂量的草鱼出血病灭活疫苗均能刺激草鱼脾脏中主要组织相容性复合体Ⅰ(MHCⅠ)、干扰素Ⅰ(IFNⅠ)、补体3 (C3)等6种主要免疫基因不同程度的上调表达, 说明疫苗刺激机体启动了非特异性和特异性免疫应答(奕志娟等, 2015)。本研究利用原核重组系统成功表达出两种基因型GCRV的VP7蛋白, 并进行了纯化制备和鉴定, 利用实验室保存的草鱼IgM单抗, 建立了三抗体夹心ELISA分析评价疫苗体液免疫效果。结果表明, 草鱼经腹腔注射不同剂量的细胞灭活疫苗后, 与对照组相比, 各免疫组血清抗体均有不同程度的提高, 且原液组与50倍稀释组基本持平, 但都高于100倍稀释组, 各免疫组在免疫后5 d即可检测到两种基因型GCRV抗体, 且抗体水平持续到84 d, 但两种基因型抗体水平达到高峰的时间不一致, 抗基因Ⅰ型GCRV抗体达到高峰是在28 d, 抗基因Ⅱ型GCRV抗体是在14 d达到高峰。两种基因型抗体水平达到高峰时的血清效价分别为1︰800、1︰600。
目前, 全球已经有很多商品化的鱼类病毒灭活疫苗, 有美国、荷兰等国的针对鲑的传染性胰腺坏死病毒灭活疫苗, 有日本的针对长吻鲹(Caranx delicatissimus)、点带石斑鱼(Epinephelus malabaricus)的虹彩病毒灭活疫苗, 有捷克的针对鲤的鲤春病毒灭活疫苗等等(吴淑勤等, 2014), 这些疫苗具有较高的免疫保护率, 如Fan等(2012)研制的针对大菱鲆红体病虹彩病毒研制的灭活疫苗通过皮下注射其免疫保护率达到83%以上。本研究利用实验室自行分离的当前流行强毒株GCRV-HZ13进行草鱼出血病二价细胞灭活疫苗免疫保护率实验, 结果表明, 原液组保护率最高, 为67%, 50倍稀释组为60%, 100倍稀释组只有33%, 因此实际应用中可考虑用50倍稀释的剂量进行草鱼免疫, 下一步将进行免疫中试实验。
目前国外鱼用疫苗自动注射装置较为完备, 为疫苗的推广使用奠定了重要基础, 朱烨等(2020)研制的鱼用疫苗自动注射装置, 以10~12 cm长的草鱼苗作为自动注射对象, 开展活鱼注射试验, 并将注射生理盐水的育苗暂养3周, 发现存活率为98.8%, 自动注射装置的研发, 成功解决了人工注射效率低、损伤大的问题, 有望为今后鱼用疫苗的推广应用提供有力的保障。
4 结论本研究从体液免疫及免疫保护率的角度出发, 对草鱼出血病二价细胞灭活疫苗进行免疫效果评价, 建立了测试未知抗体的三抗体夹心酶联免疫方法(TAS-ELISA), 评价疫苗特异性体液免疫应答的免疫效果, 并在实验室开展了二价疫苗免疫保护率实验。免疫保护率实验结果表明原液组保护率最高, 为67%, 50倍稀释组为60%, 100倍稀释组为33%, 可为进一步开展二价疫苗的免疫示范和新兽药证书申报提供基础数据。
王方华, 李安兴, 2006. 草鱼病毒性出血病研究进展. 南方水产, 2(3): 66-71 DOI:10.3969/j.issn.2095-0780.2006.03.012 |
吉华松, 张翔, 高任, 等, 2019. 草鱼出血病疫苗的发展历程及市场现状剖析. 江西水产科技, (5): 38-44 DOI:10.3969/j.issn.1006-3188.2019.05.016 |
朱烨, 徐皓, 江涛, 等, 2020. 草鱼疫苗自动注射技术研究. 渔业现代化, 47(4): 12-19 DOI:10.3969/j.issn.1007-9580.2020.04.003 |
刘栭, 冯祖强, 1977. 注射土法免疫组织浆防治草鱼鱼种出血病. 淡水渔业, (12): 10-13 |
许淑英, 李焕林, 邓国成, 等, 1994. 草鱼出血病细胞培养弱毒疫苗的制备及其免疫效果. 水产学报, 18(2): 110-117 DOI:10.3321/j.issn:1000-0615.1994.02.009 |
李丽, 赵成萍, 李宏, 等, 2011. 质粒制备绝对定量PCR标准曲线方法的建立. 农业生物技术学报, 19(6): 1157-1162 DOI:10.3969/j.issn.1674-7968.2011.06.024 |
李永刚, 曾伟伟, 王庆, 等, 2013. 草鱼呼肠孤病毒分子生物学研究进展. 动物医学进展, 34(4): 97-103 |
杨映, 于辉, 古勇明, 2015. 草鱼呼肠孤病毒研究进展. 广东农业科学, 42(15): 92-97 DOI:10.3969/j.issn.1004-874X.2015.15.017 |
吴淑勤, 陶家发, 巩华, 等, 2014. 渔用疫苗发展现状及趋势. 中国渔业质量与标准, 4(1): 1-13 |
张超, 王庆, 石存斌, 等, 2010. 草鱼呼肠孤病毒HZ08株的分离与鉴定. 中国水产科学, 17(6): 1257-1263 |
陈爱平, 江育林, 钱冬, 等, 2010. 草鱼出血病. 中国水产, (11): 56-57 |
柯丽华, 方勤, 蔡宜权, 1990. 一株新的草鱼出血病病毒分离物的特性. 水生生物学报, 14(2): 153-159 |
奕志娟, 郝贵杰, 袁雪梅, 等, 2015. 草鱼出血病灭活疫苗上调草鱼脾细胞主要免疫分子的表达. 细胞与分子免疫学杂志, 31(2): 177-181 |
FAN T J, HU X Z, WANG L Y, et al, 2012. Development of an inactivated iridovirus vaccine against turbot viral reddish body syndrome. Journal of Ocean University of China, 11(1): 65-69 |
FAN Y D, RAO S J, ZENG L B, et al, 2013. Identification and genomic characterization of a novel fish reovirus, Hubei grass carp disease reovirus, isolated in 2009 in China. Journal of General Virology, 94(10): 2266-2277 |
FANG Q, ATTOUI H, FRANÇOIS J, et al, 2000. Sequence of genome segments 1, 2, and 3 of the grass carp reovirus (genus Aquareovirus, family Reoviridae). Biochemical and Biophysical Research Communications, 274(3): 762-766 |
FANG Q, SHAH S, LIANG Y Y, et al, 2005. 3D Reconstruction and capsid protein characterization of grass carp reovirus. Science in China Series C: Life Sciences, 48(6): 593-600 |
FRANCKI R I B, FAUQUET C M, KNUDSON D L, et al, 1991. Classification and Nomenclature of Viruses: Fifth Report of the International Committee on Taxonomy of Viruses. Vienna: Springer, 186-192
|
QIU T, LU R H, ZHANG J, et al, 2001. The molecular characterization of RNA segment S9 of grass carp hemorrhage virus (GCHV), an aquareovirus. Aquaculture, 203(1/2): 1-7 |
RANGEL A A C, ROCKEMANN D D, HETRICK F M, et al, 1999. Identification of grass carp haemorrhage virus as a new genogroup of aquareovirus. Journal of General Virology, 80(9): 2399-2402 |
WANG Q, ZENG W W, LIU C, et al, 2012. Complete genome sequence of a reovirus isolated from grass carp, indicating different genotypes of GCRV in China. Journal of Virology, 86(22): 12466 |
WINTON J R, LANNAN C N, FRYER J L, et al, 1987. Morphological and biochemical properties of four members of a novel group of reoviruses isolated from aquatic animals. Journal of General Virology, 68(2): 353-364 |